Effects of Arbuscular Mycorrhizal Fungi on Growth, Photosynthesis, and Nutrient Uptake of Zelkova serrata (Thunb.) Makino Seedlings under Salt Stress

Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Seedlings, AMF Inocula, and Soil
2.2. Inoculation and Experimental Design
2.3. Plant Growth Parameters and AMF Colonization
2.4. Leaf Photosynthesis, Photosynthetic Pigments, and Chlorophyll a Fluorescence
2.5. Leaf Water Status and Membrane Stability Index
2.6. Leaf and Root Nutrients Content
2.7. Statistical Analysis
3. Results
3.1. AMF Colonization
3.2. Plant Growth
3.3. Leaf Photosynthesis, Photosynthetic Pigments, and Fluorescence Parameters
3.4. Leaf Relative Water Content and Membrane Stability Index
3.5. Leaf and Root Nutrients and Ions Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Horikawa, K. Coast. Dunes: Form and Process; Nordstrom, K.F., Psuty, N., Carter, B., Eds.; Wiley: Chichester, UK, 1990; ISBN 0-471-91842-3. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; Mascarenhas, A.; Jagtap, T.G. An evaluation of flora from coastal sand dunes of India: Rationale for conservation and management. Ocean Coast. Manag. 2011, 54, 181–188. [Google Scholar] [CrossRef]
- Ruizlozano, J.M.; Porcel, R.; Azcón, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Lӓuchi, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Dames, J.F.; Gupta, A.; Sharma, S.; Gilbert, J.A.; Ahmad, P. Current developments in arbuscular mycorrhizal fungi research and its role in salinity stress alleviation: A biotechnological perspective. Crit. Rev. Biotechnol. 2015, 35, 461–474. [Google Scholar] [CrossRef] [PubMed]
- LóPez-Ráez, J.A. How drought and salinity affect arbuscular mycorrhizal symbiosis and strigolactone biosynthesis? Planta 2016, 243, 1375–1385. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bomito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum may be partly related to elevated K/Na ratios in root and shoot tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef]
- Frosi, G.; Barros, V.A.; Oliveira, M.T.; Santos, M.; Ramos, D.G.; Maia, L.C.; Santos, M.G. Arbuscular mycorrhizal fungi and foliar phosphorus inorganic supply alleviate salt stress effects in physiological attributes, but only arbuscular mycorrhizal fungi increase biomass in woody species of a semiarid environment. Tree Physiol. 2017, 38, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Leifheit, E.F.; Veresoglou, S.D.; Lehmann, A.; Kathryn Morris, E.; Rillig, M.C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation—A meta-analysis. Plant Soil 2014, 374, 523–537. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boughattas, S.; Hu, S.J.; Oh, S.H.; Sa, T.M. A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress. Mycorrhiza 2014, 24, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Estrada, B.; Aroca, R.; Maathuis, F.J.; Barea, J.M.; Ruizlozano, J.M. Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. Plant Cell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcel, R.; Redondogómez, S.; Mateosnaranjo, E.; Aroca, R.; Garcia, R.; Ruiziozano, J.M. Arbuscular mycorrhizal symbiosis ameliorates the optimum quantum yield of photosystem II and reduces non-photochemical quenching in rice plants subjected to salt stress. J. Plant Physiol. 2015, 185, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Bago, A.; Sutka, M.; Paz, J.A.; Cano, C.; Amodeo, G.; Ruiz-Lozano, J.M. Expression analysis of the first arbuscular mycorrhizal fungi aquaporin described reveals concerted gene expression between salt-stressed and nonstressed mycelium. Mol. Plant Microbe Interact. 2009, 22, 1169–1178. [Google Scholar] [CrossRef]
- Dou, Q.Q.; Jiao, X.J.; Zhang, M.; Kai-Yue, H.H.; Huang, L.B. Physiological response of Zelkova schneideriana seedlings in the soil under NaCl stress. Acta Bot. Boreali-Occident. Sin. 2009, 22, 2063–2069. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Liu, X.; Fu, Z.Y.; Zhang, B.; Zhai, L.; Meng, M.J.; Lin, J.; Zhuang, J.Y.; Wang, G.G.; Zhang, J.C. Effects of sulfuric, nitric, and mixed acid rain on Chinese fir sapling growth in Southern China. Ecotoxicol. Environ. Saf. 2018, 160, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Yue, J.M.; Fu, Z.Y.; Zhang, L.; Zhang, Z.H.; Zhang, J.C. The positive effect of different 24-epiBL pretreatments on salinity tolerance in Robinia pseudoacacia L. seedlings. Forests 2019, 10, 4. [Google Scholar] [CrossRef]
- Yue, J.M.; You, Y.Y.; Zhang, L.; Fu, Z.Y.; Wang, J.P.; Zhang, J.C.; Robert, D.G. Exogenous 24-epibrassinolide alleviates effects of salt stress on chloroplasts and photosynthesis in Robinia pseudoacacia L. seedlings. J. Plant Growth Regul. 2018, 1–14. [Google Scholar] [CrossRef]
- Allen, M. Mycorrhizae and rehabilitation of disturbed arid soils: Processes and practices. Arid Soil Res. Rehabil. 1989, 3, 229–241. [Google Scholar] [CrossRef]
- Paludan-Müller, G.; Saxe, H.; Pedersen, L.B.; Randrup, T.B. Differences in salt sensitivity of four deciduous tree species to soil or airborne salt. Physiol. Plant. 2010, 114, 223–230. [Google Scholar] [CrossRef]
- Allen, J.A.; Chambers, J.L.; Stine, M. Prospects for increasing the salt tolerance of forest trees: A review. Tree Physiol. 1994, 14, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Estrada, B.; Barea, M.J.; Aroca, R.; Ruiz-Lozano, J.M. A native Glomus intraradices strain from a Mediterranean saline area exhibits salt tolerance and enhanced symbiotic efficiency with maize plants under salt stress conditions. Plant Soil 2013, 366, 333–349. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.M. Arbuscular mycorrhizal fungal application to improve growth and tolerance of wheat (Triticum aestivum L.) plants grown in saline soil. Acta Physiol. Plant. 2012, 34, 267–277. [Google Scholar] [CrossRef]
- Navarro, J.M.; Olaya, P.R.T.; Asunciã, N.M. Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J. Plant Physiol. 2014, 171, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Hajiboland, R.; Aliasgharzadeh, N.; Laiegh, S.F.; Poschenrieder, C. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 2010, 331, 313–327. [Google Scholar] [CrossRef]
- Pollastri, S.; Savvides, A.; Pesando, M.; Lumini, E.; Volpe, M.G.; Ozudogru, E.E.; Faccio, A.; Cunzo, F.F.; Michelozzi, M.; Lambardi, M.; et al. Impact of two arbuscular mycorrhizal fungi on Arundo donax L. response to salt stress. Planta 2018, 247, 573–585. [Google Scholar] [CrossRef]
- Liu, S.L.; Guo, X.L.; Feng, G.; Maimaitiaili, B.; Fan, J.L.; He, X.H. Indigenous arbuscular mycorrhizal fungi can alleviate salt stress and promote growth of cotton and maize in saline fields. Plant Soil 2016, 398, 195–206. [Google Scholar] [CrossRef]
- Qu, L.Q.; Huang, Y.Y.; Zhu, C.M.; Zeng, H.Q.; Shen, C.J.; Liu, C.; Zhao, Y.; Pi, E.X. Rhizobia-inoculation enhances the soybean’s tolerance to salt stress. Plant Soil 2016, 400, 209–222. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Zhang, K.; Tian, C.Y.; Guo, J.X. Arbuscular mycorrhizal fungi improve plant growth of Ricinus communis by altering photosynthetic properties and increasing pigments under drought and salt stress. Ind. Crop. Prod. 2018, 117, 13–19. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Zhang, X.; Tang, M. Arbuscular mycorrhizal symbiosis alleviates salt stress in Black locust through improved photosynthesis, water status, and K+/Na+ homeostasis. Front. Plant Sci. 2017, 8, 1739. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Sui, X.; Fan, X.X.; Jia, T.T.; Song, F.Q. Arbuscular mycorrhizal symbiosis modulates antioxidant response and ion distribution in salt-stressed Elaeagnus angustifolia Seedlings. Front. Microbiol. 2018, 9, 652. [Google Scholar] [CrossRef] [PubMed]
- Juniper, S.; Abbott, L.K. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza 2006, 16, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, F.; Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microb. Ecol. 2008, 55, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Zhang, J.C.; Huang, Y.Q. Effects of arbuscular mycorrhizal fungi on the drought tolerance of Cyclobalanopsis glauca seedlings under greenhouse conditions. New For. 2014, 45, 545–556. [Google Scholar] [CrossRef]
- Urgiles, N.; Strauß, A.; Loján, P.; Schüßler, A. Cultured arbuscular mycorrhizal fungi and native soil inocula improve seedling development of two pioneer trees in the Andean region. New For. 2014, 45, 859–874. [Google Scholar] [CrossRef]
- Azcón, R.; El-Atrash, F. Influence of arbuscular mycorrhizae and phosphorus fertilization on growth, nodulation and N2 fixation (15N) in Medicago sativa at four salinity levels. Biol. Fertil. Soils 1997, 24, 81–86. [Google Scholar] [CrossRef]
- Miransari, M.; Bahrami, H.A.; Rejali, F.; Malakouti, M.J. Using arbuscular mycorrhiza to alleviate the stress of soil compaction on wheat (Triticum aestivum L.) growth. Soil Biol. Biochem. 2008, 40, 1197–1206. [Google Scholar] [CrossRef]
- Pitman, M.G.; Läuchli, A. Global Impact of salinity and agricultural ecosystems. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2002; pp. 3–20. [Google Scholar]
- Akram, M.S.; Ashraf, M. Exogenous application of potassium dihydrogen phosphate can alleviate the adverse effects of salt stress on sunflower. J. Plant Nutr. 2011, 34, 1041–1057. [Google Scholar] [CrossRef]
- Zuccarini, P.; Okurowska, P. Effects of mycorrhizal colonization and fertilization on growth and photosynthesis of sweet basil under salt stress. J. Plant Nutr. 2008, 31, 497–513. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 2014, 98, 20–31. [Google Scholar] [CrossRef]
- Zai, X.M.; Zhu, S.N.; Qin, P.; Wang, X.Y.; Che, L.; Luo, F.X. Effect of Glomus mosseae on chlorophyll content, chlorophyll fluorescence parameters, and chloroplast ultrastructure of beach plum (Prunus maritima) under NaCl stress. Photosynthetica 2012, 50, 323–328. [Google Scholar] [CrossRef]
- Zhu, X.C.; Song, F.B.; Xu, H.W. Arbuscular mycorrhizae improves low temperature stress in maize via alterations in host water status and photosynthesis. Plant Soil 2010, 331, 129–137. [Google Scholar] [CrossRef]
- Aroca, R.; Ruiz-Lozano, J.M.; Zamarreño, A.M.; Paz, J.A.; Garcia-Mina, J.M.; Pozo, M.J.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. J. Plant Physiol. 2013, 170, 47–55. [Google Scholar] [CrossRef]
- Kiers, E.T.; Duhamei, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef]
- Tian, C.Y.; Feng, G.; Li, X.L.; Zhang, F.S. Different effects of arbuscular mycorrhizal fungal isolates from saline or non-saline soil on salinity tolerance of plants. Appl. Soil Ecol. 2004, 26, 143–148. [Google Scholar] [CrossRef]
- Giri, B.; Mukerji, K.G. Mycorrhizal inoculant alleviates salt stress in Sesbania aegyptiaca and Sesbania grandiflora under field conditions: Evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 2004, 14, 307–312. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant physiology. Q. Rev. Biol. 2006, 167, 161–168. [Google Scholar]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass, and mineral nutrition of Acacia auriculiformis. Biol. Fertil. Soils 2003, 38, 170–175. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Symbiotic efficiency and infectivity of an autochthonous arbuscular mycorrhizal Glomus sp. from saline soils and Glomus deserticola under salinity. Mycorrhiza 2000, 10, 137–143. [Google Scholar] [CrossRef]
- Cantrell, I.C.; Linderman, R.G. Preinoculation of lettuce and onion with VA mycorrhizal fungi reduces deleterious effects of soil salinity. Plant Soil 2001, 233, 269–281. [Google Scholar] [CrossRef]
- Rinaldelli, E.; Mancuso, S. Response of young mycorrhizal and non-mycorrhizal plants of olive tree (Olea europaea L.) to saline conditions. I. Short-term electrophysiological and long-term vegetative salt effects. Adv. Hortic. Sci. 1996, 10, 126–134. [Google Scholar]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.A.; Eissa, A.M.; El-Shazly, S.M.; Nasr, A.M.A. Improved growth of salinity-stressed citrus after inoculation with mycorrhizal fungi. Sci. Hortic. 2011, 130, 624–632. [Google Scholar] [CrossRef]





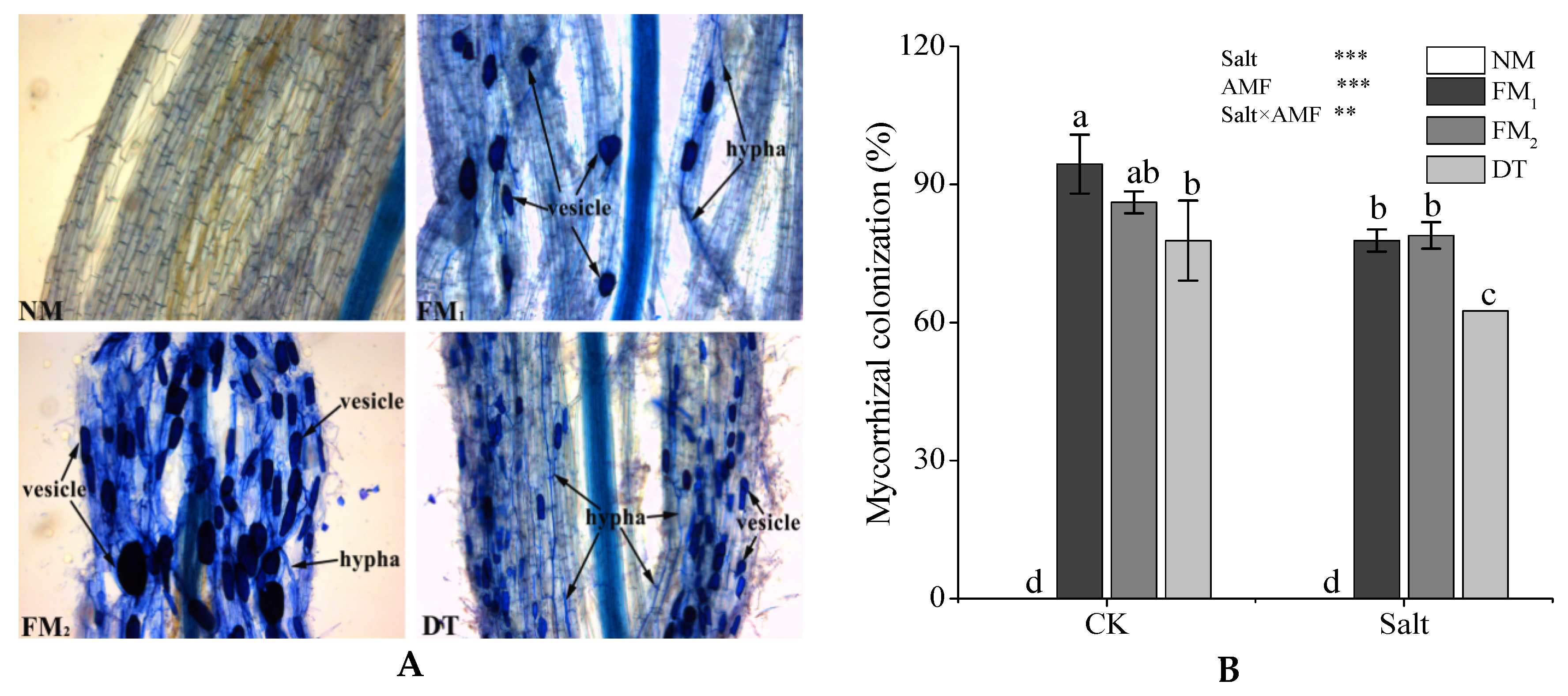{kind=link}
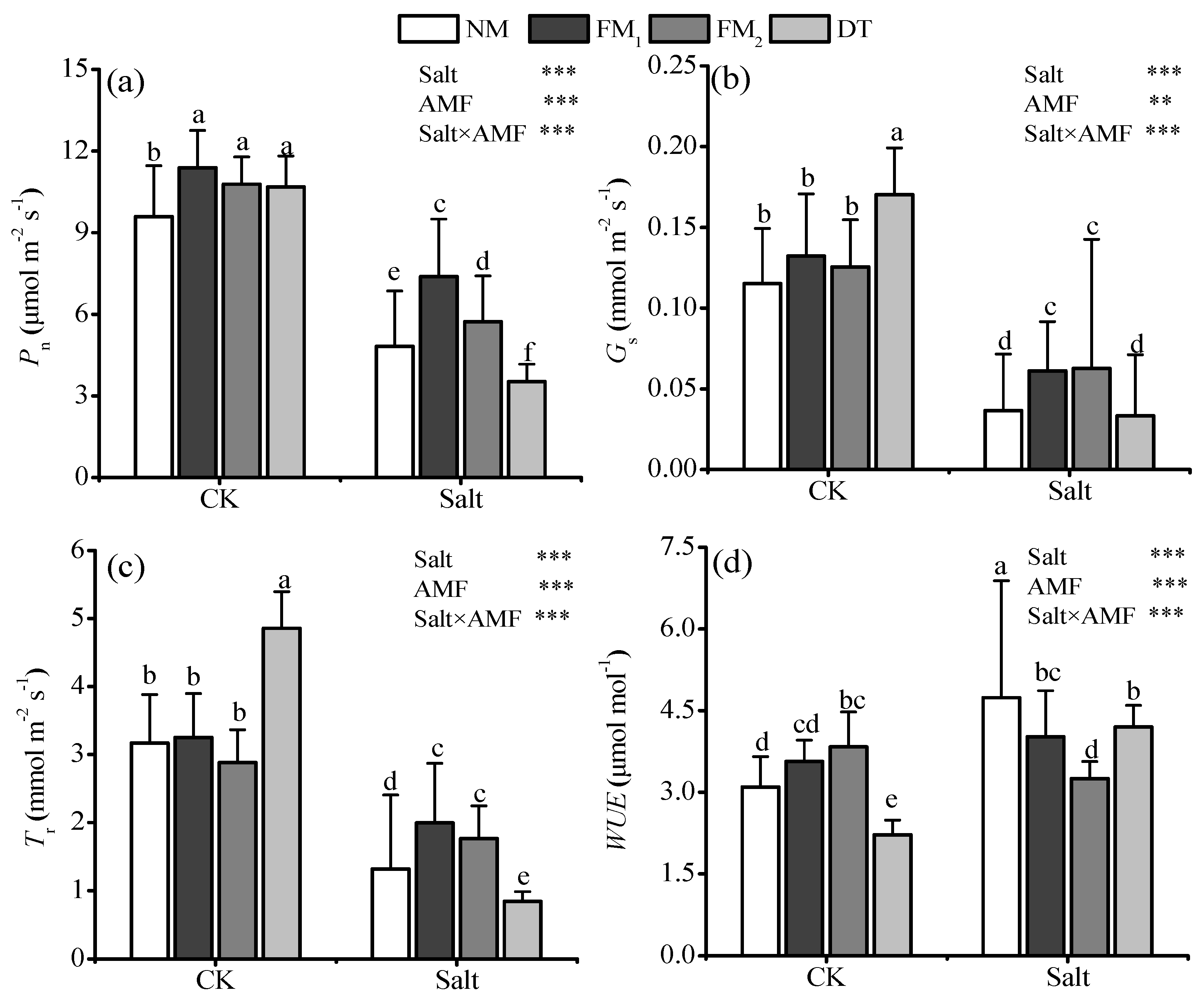{kind=link}
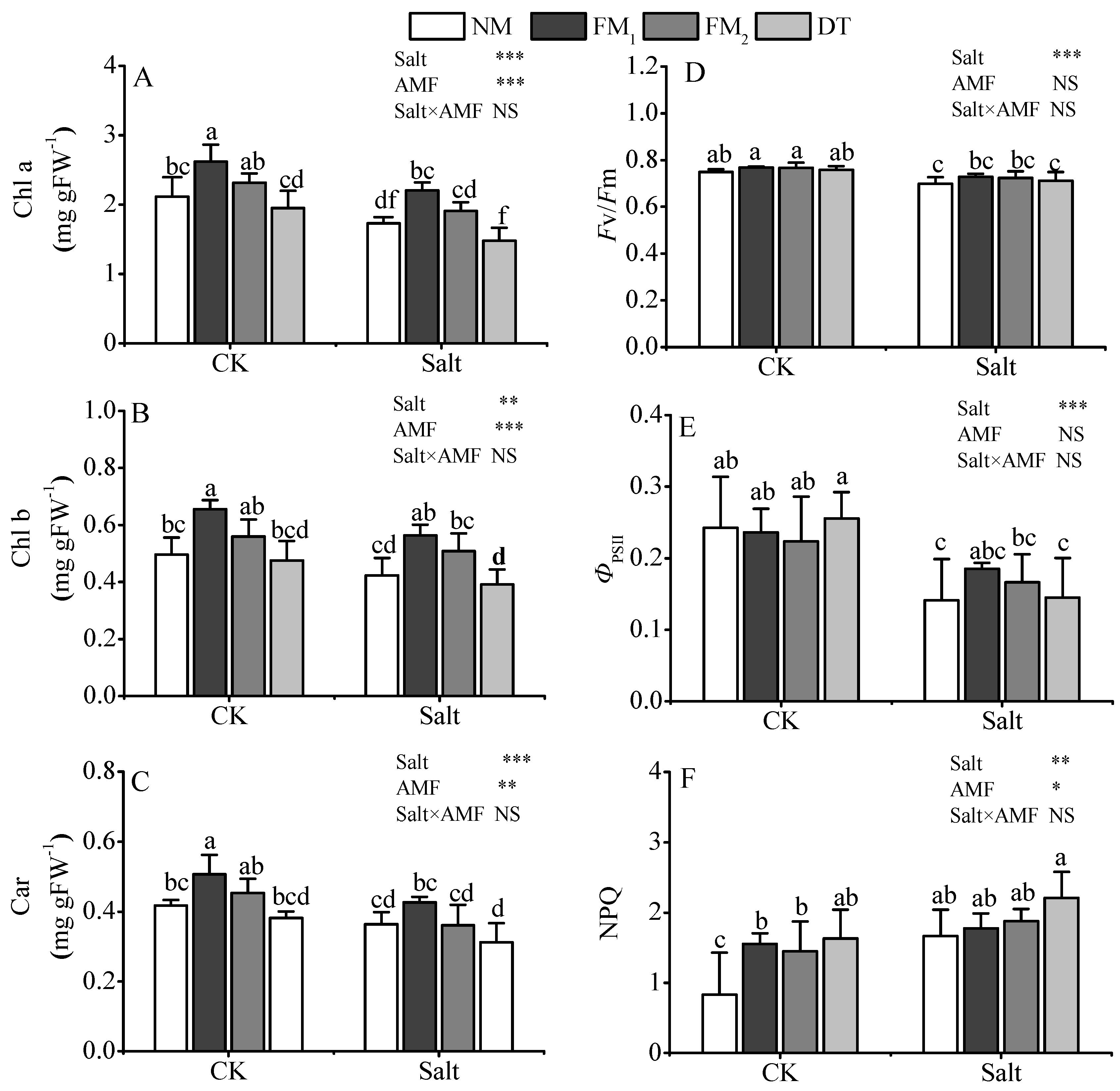{kind=link}
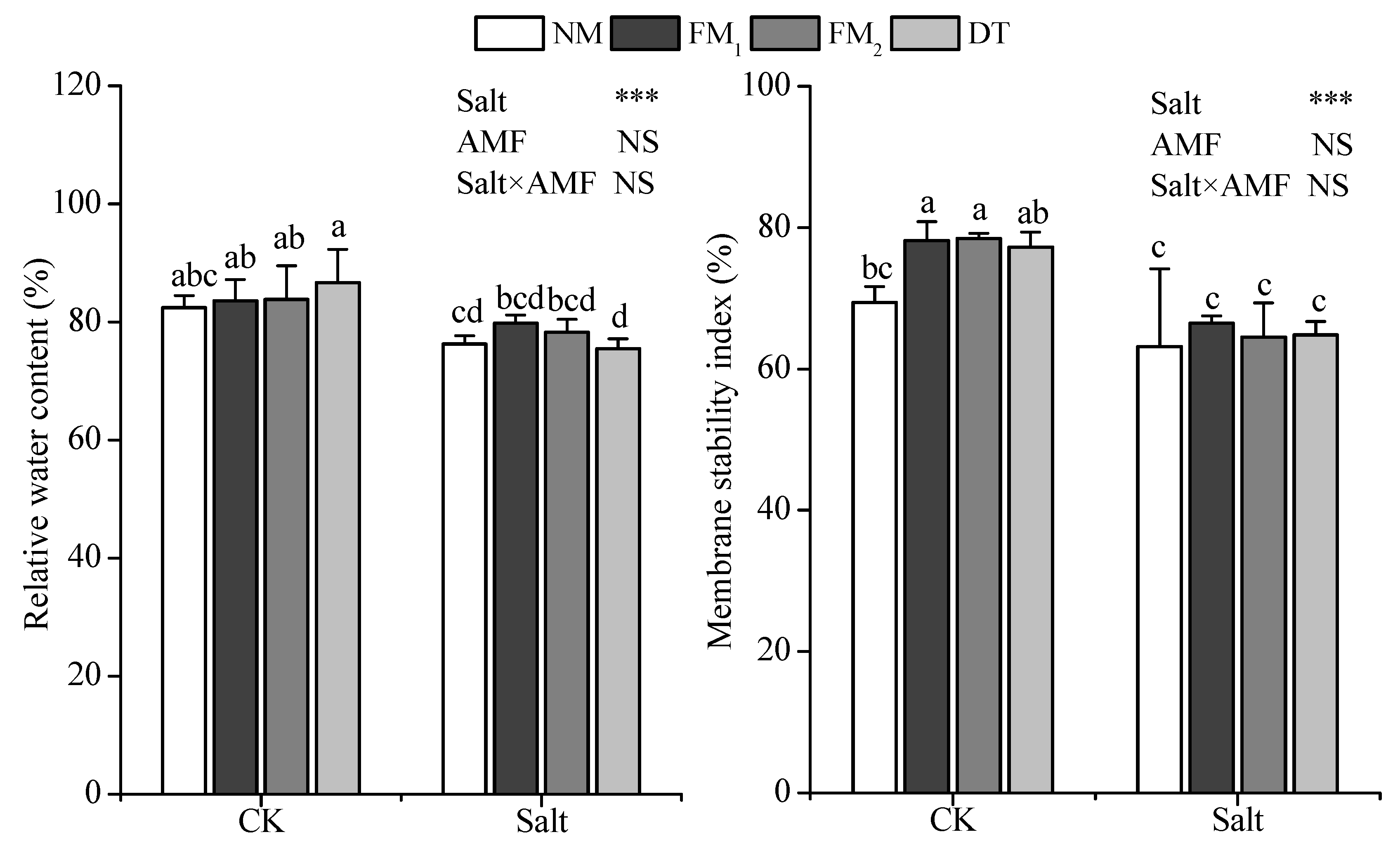{kind=link}
| Salt Status | AM Treatments | Growth of Seedling Height (cm) | Growth of Seedling Diameter (cm) | Leaf Area (cm2) | Leaf Dry Weight (g) | Stem Dry Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|---|---|---|
| CK | NM | 24.2 ± 8.5 b | 3.0 ± 1.7 ab | 16.0 ± 0.7 ab | 7.5 ± 0.9 ab | 18.6 ± 1.7 abc | 9.9 ± 1.9 ab |
| FM1 | 29.2 ± 7.1 ab | 3.2 ± 0.7 a | 17.6 ± 0.7 a | 8.7 ± 1.5 a | 21.2 ± 3.7 ab | 10.6 ± 2.2 ab | |
| FM2 | 35.5 ± 5.4 a | 2.9 ± 0.5 ab | 16.5 ± 0.9 ab | 8.0 ± 0.3 a | 19.7 ± 3.2 abc | 10.3 ± 3.6 ab | |
| DT | 30.3 ± 4.4 ab | 3.1 ± 0.7 a | 16.4 ± 2.0 ab | 9.5 ± 1.6 a | 23.3 ± 5.1 a | 11.3 ± 2.2 a | |
| Salt | NM | 9.5 ± 3.3 c | 1.8 ± 0.4 c | 10.9 ± 1.1 c | 4.2 ± 1.0 c | 14.0 ± 1.4 d | 7.1 ± 0.9 b |
| FM1 | 8.7 ± 4.7 c | 2.7 ± 0.4 ab | 14.3 ± 0.6 b | 5.5 ± 1.6 bc | 16.9 ± 1.0 bcd | 8.7 ± 0.2 ab | |
| FM2 | 9.6 ± 5.4 c | 2.8 ± 0.6 ab | 11.6 ± 1.9 c | 5.0 ± 1.3 c | 15.0 ± 1.5 cd | 8.2 ± 0.5 ab | |
| DT | 8.9 ± 4.5 c | 2.1 ± 0.3 bc | 10.2 ± 0.7 c | 4.2 ± 0.2 c | 14.2 ± 0.9 d | 7.1 ± 1.0 b | |
| Significance | |||||||
| Salt | *** | ** | *** | *** | *** | ** | |
| AMF | NS | NS | ** | NS | NS | NS | |
| Salt × AMF | NS | NS | NS | NS | NS | NS |
| Salt Status | AM Treatments | C (mg/g) | N (mg/g) | P (mg/g) | K (mg/g) | Ca (mg/g) | Mg (mg/g) | Na (mg/g) |
|---|---|---|---|---|---|---|---|---|
| Leaf | ||||||||
| CK | NM | 435.1 ± 12.1 bc | 23.6 ± 2.9 ab | 1.2 ± 0.2 a | 12.9 ± 0.4 cd | 22.3 ± 1.6 bc | 2.4 ± 0.3 abc | 1.4 ± 0.0 d |
| FM1 | 448.3 ± 3.1 ab | 24.1 ± 1.8 ab | 1.1 ± 0.1 ab | 11.2 ± 0.7 d | 21.5 ± 1.6 bc | 2.3 ± 0.1 bc | 1.0 ± 0.1 d | |
| FM2 | 443.6 ± 12.5 abc | 24.5 ± 2.1 a | 1.3 ± 0.2 a | 12.5 ± 0.5 d | 21.9 ± 1.5 bc | 2.7 ± 0.4 ab | 1.1 ± 0.1 d | |
| DT | 457.3 ± 8.3 a | 22.7 ± 1.3 abc | 1.2 ± 0.1 a | 11.6 ± 0.6 d | 18.6 ± 3.4 c | 2.4 ± 0.2 bc | 1.0 ± 0.1 d | |
| Salt | NM | 431.0 ± 4.1 c | 20.5 ± 0.5 bc | 0.9 ± 0.2 bc | 14.65 ± 1.3 bc | 21.0 ± 08 bc | 1.9 ± 0.2 c | 6.6 ± 0.1 a |
| FM1 | 432.8 ± 10.1 bc | 23.4 ± 2.7 ab | 1.0 ± 0.2 abc | 18.3 ± 1.5 a | 24.4 ± 2.1 ab | 2.6 ± 0.1 ab | 5.2 ± 0.4 c | |
| FM2 | 432.0 ± 1.1 c | 19.7 ± 2.3 c | 0.9 ± 0.1 bc | 19.3 ± 2.2 a | 27.0 ± 4.7 a | 2.9 ± 0.4 a | 5.9 ± 0.5 b | |
| DT | 431.2 ± 9.0 c | 19.5 ± 0.8 c | 0.8 ± 0.2 c | 15.1 ± 0.2 b | 20.5 ± 1.6 bc | 2.2 ± 0.2 bc | 6.3 ± 0.4 ab | |
| Significance | ||||||||
| Salt | ** | ** | *** | *** | NS | NS | *** | |
| AMF | NS | NS | NS | ** | * | ** | ** | |
| Salt × AMF | NS | NS | NS | ** | NS | NS | * | |
| Root | ||||||||
| CK | NM | 406.7 ± 0.8 b | 14.4 ± 1.0 ab | 1.5 ± 0.1 bc | 7.1 ± 2.0 ab | 17.4 ± 2.7 a | 2.3 ± 0.3 a | 1.1 ± 0.4 d |
| FM1 | 404.6 ± 5.4 b | 14.6 ± 1.0 ab | 1.4 ± 0.3 bcd | 6.7 ± 0.3 b | 17.5 ± 2.8 a | 2.1 ± 0.4 a | 1.0 ± 0.2 d | |
| FM2 | 400.2 ± 1.3 b | 14.1 ± 0.9 ab | 2.1 ± 0.0 a | 8.5 ± 0.9 a | 19.9 ± 2.0 a | 2.5 ± 0.4 a | 1.3 ± 0.1 d | |
| DT | 420.9 ± 1.2 a | 12.4 ± 0.2 b | 1.3 ± 0.1 bcd | 7.1 ± 0.9 ab | 18.6 ± 3.9 a | 2.5 ± 0.5 a | 1.1 ± 0.2 d | |
| Salt | NM | 396.6 ± 4.2 b | 16.0 ± 1.5 a | 1.0 ± 0.1 cd | 3.0 ± 0.3 d | 16.6 ± 1.8 a | 1.9 ± 0.2 a | 7.6 ± 0.3 a |
| FM1 | 405.8 ± 9.7 b | 16.0 ± 0.6 a | 1.3 ± 0.4 bcd | 4.8 ± 0.2 c | 16.3 ± 1.3 a | 2.1 ± 0.3 a | 6.2 ± 0.3 c | |
| FM2 | 395.8 ± 15.0 b | 16.2 ± 0.8 a | 1.7 ± 0.4 ab | 4.5 ± 0.8 cd | 18.3 ± 0.5 a | 2.1 ± 0.2 a | 6.7 ± 0.2 b | |
| DT | 395.7 ± 10.6 b | 14.9 ± 3.6 ab | 0.8 ± 0.2 d | 4.1 ± 0.1 cd | 15.6 ± 1.8 a | 1.9 ± 0.1 a | 7.6 ± 0.3 a | |
| Significance | ||||||||
| Salt | ** | ** | ** | *** | NS | * | *** | |
| AMF | NS | NS | ** | NS | NS | NS | *** | |
| Salt × AMF | NS | NS | NS | NS | NS | NS | ** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Fu, Z.; Ren, Q.; Zhu, L.; Lin, J.; Zhang, J.; Cheng, X.; Ma, J.; Yue, J. Effects of Arbuscular Mycorrhizal Fungi on Growth, Photosynthesis, and Nutrient Uptake of Zelkova serrata (Thunb.) Makino Seedlings under Salt Stress. Forests 2019, 10, 186. https://doi.org/10.3390/f10020186
Wang J, Fu Z, Ren Q, Zhu L, Lin J, Zhang J, Cheng X, Ma J, Yue J. Effects of Arbuscular Mycorrhizal Fungi on Growth, Photosynthesis, and Nutrient Uptake of Zelkova serrata (Thunb.) Makino Seedlings under Salt Stress. Forests. 2019; 10(2):186. https://doi.org/10.3390/f10020186
Chicago/Turabian StyleWang, Jinping, Zhiyuan Fu, Qiong Ren, Lingjun Zhu, Jie Lin, Jinchi Zhang, Xuefei Cheng, Jieyi Ma, and Jianmin Yue. 2019. "Effects of Arbuscular Mycorrhizal Fungi on Growth, Photosynthesis, and Nutrient Uptake of Zelkova serrata (Thunb.) Makino Seedlings under Salt Stress" Forests 10, no. 2: 186. https://doi.org/10.3390/f10020186