Effects of Salt on Root Aeration, Nitrification, and Nitrogen Uptake in Mangroves

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Preparation
2.2. Experimental Design and Salt Treatments
2.3. Plant Growth and Concentrations of N and Na+ in the Tissues of A. marina
2.4. N Dynamics in the Soils
2.5. Soil DNA Extraction and qPCR Analysis
2.6. The Measurements of ROL and Root Porosity
2.7. Root Suberization Observation
2.8. The Measurements of NH4+ and NO3− Fluxes at the Rhizo–Root Interface
2.9. Statistical Analysis
3. Results
3.1. The Effects of Salt on Growth and N Uptake by A. marina
3.2. The Effects of Salt on ROL and Root Porosity
3.3. The Effects of Salt on Root Suberization
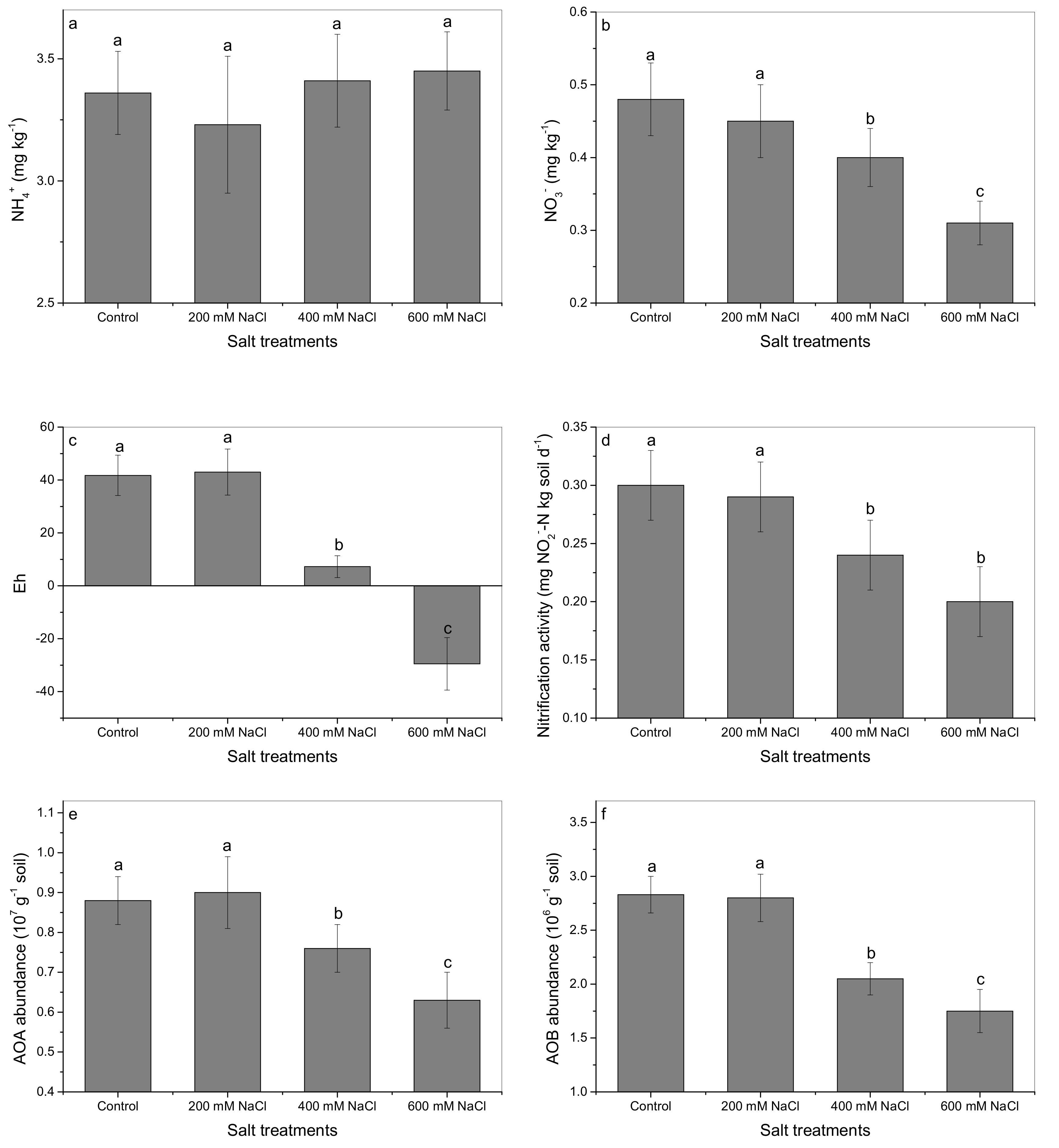
3.4. The Effects of Salt on N Dynamics in the Soils Planted with A. marina
3.5. The Effects of Salt on NH4+ and NO3− Fluxes at the Rhizo–Root Interface
4. Discussion
4.1. Growth and N Uptake by A. marina under Elevated Salinity
4.2. The Effects of Salt on Root Anatomy and ROL
4.3. N Uptake by A. marina Stressed by Salt in Relation to ROL and Root Suberzation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, W.Q.; Li, X.F.; Wang, M. Propagules dispersal determines mangrove zonation at intertidal and estuarine scales. Forests 2019, 10, 245. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, P.; Ramanathan, A.L.; Kumar, A.; Singhal, R.K.; Datta, D.; Venkatesh, M. Trace metal distribution, assessment and enrichment in the surface sediments of Sundarban mangrove ecosystem in India and Bangladesh. Mar. Pollut. Bull. 2018, 127, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, S.P.; Kazama, S.; Sawamoto, M. Effects of climate and land use changes on groundwater resources in coastal aquifers. J. Environ. Manag. 2006, 80, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J.; Holland, E.A. Net primary production in tropical forests: An evaluation and synthesis of existing field data. Ecol. Appl. 2001, 11, 371–384. [Google Scholar] [CrossRef]
- Zheng, W.J.; Wang, W.Q.; Lin, P. Dynamics of element contents during the development of the hypocotyles and leaves of certain mangrove species. J. Exp. Mar. Biol. Ecol. 1999, 233, 247–257. [Google Scholar] [CrossRef]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Jiang, G.F.; Goodale, U.M.; Liu, Y.Y.; Hao, G.Y.; Cao, K.F. Salt management strategy defines the stem and leaf hydraulic characteristics of six mangrove tree species. Tree Physiol. 2017, 37, 289–401. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Lee, S.C.; Chen, T.H.; Tian, G.L.; Chiu, C.Y. Water salinity effects on growth and nitrogen assimilation rate of mangrove (Kandelia candel) seedlings. Aquat. Bot. 2017, 137, 50–55. [Google Scholar] [CrossRef]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef] [Green Version]
- Feller, I.C.; McKee, K.L.; Whigham, D.F.; O’Neill, J.P. Nitrogen vs. phosphorus limitation across an ecotonal gradient in a mangrove forest. Biogeochemistry 2003, 62, 145–175. [Google Scholar] [CrossRef]
- Lin, Y.M.; Liu, X.W.; Zhang, H.; Fan, H.Q.; Lin, G.H. Nutrient conservation strategies of a mangrove species Rhizophora stylosa under nutrient limitation. Plant Soil 2010, 326, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhou, H.C.; Pan, Y.; Shyla, F.S.; Tam, N.F.Y. Effects of polybrominated diphenyl ethers and plant species on nitrification, denitrification and anammox in mangrove soils. Sci. Total Environ. 2016, 553, 60–70. [Google Scholar] [CrossRef]
- Alongi, D.M. Impact of global change on nutrient dynamics in mangrove forests. Forests 2018, 9, 596. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Nohara, S.; Takagi, H.; Anzai, Y. Contrast of nitrogen contents around roots of mangrove plants. Plant Soil 2011, 399, 471–483. [Google Scholar] [CrossRef]
- Wang, Y.F.; Gu, J.D. Higher diversity of ammonia/ammonium-oxidizing prokaryotes in constructed freshwater wetland than natural coastal marine wetland. Appl. Microbiol. Biotechnol. 2013, 97, 7015–7033. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Wu, J.P.; Li, H.L.; Hong, Y.G.; Wilson, A.M.; Jiao, J.J.; Shananan, M. Nitrogen fate in a subtropical mangrove swamp: Potential association with seawater-groundwater exchange. Sci. Total Environ. 2018, 635, 586–597. [Google Scholar] [CrossRef]
- Raman, D.R.; Spanswick, R.M.; Walker, L.P. The kinetics of nitrate uptake from lowing nutrient solutions by rice: Influence of pretreatment and light. Bioresour. Technol. 1995, 53, 125–132. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Siddiqi, M.Y.; Glass, A.D.M.; Kirk, G.J.D. Nitrate-ammonium synergism in rice: A subcellular flux analysis. Plant Physiol. 1999, 119, 1041–1046. [Google Scholar] [CrossRef] [Green Version]
- Youssef, T.; Saenger, P. Anatomical adaptive strategies to flooding and rhizosphere oxidation in mangrove seedlings. Aust. J. Bot. 1996, 44, 297–313. [Google Scholar] [CrossRef]
- Pi, N.; Tam, N.F.Y.; Wu, Y.; Wong, M.H. Root anatomy and spatial pattern of radial oxygen loss of eight true mangrove species. Aquat. Bot. 2009, 90, 222–230. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, Y.S.; Fei, J.; Jiang, Z.Y.; Ye, Z.H. Differences in root aeration, iron plaque formation and waterlogging tolerance in six mangroves along a continues tidal gradient. Ecotoxicology 2015, 242, 1659–1667. [Google Scholar] [CrossRef]
- Armstrong, W. Radial oxygen losses from intact rice roots as affected by distance from apex, respiration and waterlogging. Plant Physiol. 1971, 25, 192–197. [Google Scholar] [CrossRef]
- Armstrong, W.; Beckett, P.M. Internal aeration and the development of stelar anoxia in submerged roots: A multishelled mathematical model combining axial diffusion of oxygen in the cortex with radial losses to the stele, the wall layers and the rhizosphere. New Phytol. 1987, 105, 221–245. [Google Scholar] [CrossRef]
- Colmer, T.D.; Pedersen, O. Oxygen dynamics in submerged rice (Oryza sativa). New Phytol. 2008, 178, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Fan, X.R.; Shen, Q.R. The relationship between rhizosphere nitrification and nitrogen-use efficiency in rice plants. Plant Cell Environ. 2008, 31, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Phukan, U.J.; Mishra, S.; Shukla, R.K. Waterlogging and submergence stress: Affects and acclimation. Crit. Rev. Biotechnol. 2016, 36, 956–966. [Google Scholar] [CrossRef]
- Evans, D.E. Aerenchyma formation. New Phytol. 2003, 161, 35–49. [Google Scholar] [CrossRef]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Soukup, A.; Armstrong, W.; Schreiber, L.; Franke, R.; Votrubová, O. Apoplastic barriers to radial oxygen loss and solute penetration: A chemical and functional comparison of the exodermis of two wetland species, Phragmites australis and Glyceria maxima. New Phytol. 2007, 173, 264–278. [Google Scholar] [CrossRef]
- Garthwaite, A.J.; Armstrong, W.; Colmer, T.D. Assessment of O2 diffusivity across the barrier to radial O2 loss in adventitious roots of Hordeum marinum. New Phytol. 2008, 179, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, B.; Gimmler, H. Cell wall adaptations to multiple environmental stresses in maize roots. Plant Cell Environ. 2000, 51, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, M.; Beisson, F.; Li, Y.H.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Jyothi-Prakash, P.A.; Qin, L.; He, J.; Lin, Q.S.; Loh, C.S.; Kumar, P.P. Role of hydrophobic barriers in salt exclusion of a mangrove plant Avicennia officinalis. Plant Cell Environ. 2014, 37, 1656–1671. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, Y.S.; Ye, Z.H.; Chen, D.T.; Wang, Y.T.; Peng, Y.L.; Wang, L.Y. Influence of N deficiency and salinity on metal (Pb, Zn and Cu) accumulation and tolerance by Rhizophora stylosa in relation to root anatomy and permeability. Environ. Pollut. 2012, 164, 110–117. [Google Scholar] [CrossRef]
- Miyamoto, N.; Steudle, E.; Hirasawa, T.; Lafitte, R. Hydraulic conductivity of rice roots. J. Exp. Bot. 2001, 52, 1835–1846. [Google Scholar] [CrossRef]
- Kludze, H.K.; DeLaune, R.D.; Patrick, W.H. Aerenchyma formation and methane and oxygen exchange in rice. Soil Sci. Soc. Am. J. 1993, 57, 386–391. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan red 7B or Fluorol yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Luo, J.; Li, H.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, B.J.; Robbins, S.; Proter, R.B. Nitrogen uptake over entire root systems of tree seedlings. Tree Physiol. 2014, 34, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Downton, W.J.S. Growth and osmotic relations of the mangrove Avicennia marina, as influenced by salinity. Aust. J. Plant Physiol. 1982, 9, 519–528. [Google Scholar] [CrossRef]
- Burchett, M.D.; Clarke, C.J.; Field, C.D.; Pulkownik, A. Growth and respiration in two mangrove species at a range of salinities. Physiol. Plant 1989, 75, 299–303. [Google Scholar] [CrossRef]
- Patel, N.T.; Gupta, A.; Pandey, A.N. Salinity tolerance of Avicennia marina (Forssk.) Vierh. from Gujarat coasts of India. Aquat. Bot. 2010, 93, 9–16. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Effects of NaCl stress on nitrogen and phosphorous metabolism in a true mangrove Bruguiera parviflora grown under hydroponic culture. J. Plant Physiol. 2004, 161, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.D.; Inafuku, M.; Iwasaki, H.; Taira, N.; Mostofa, M.G.; Oku, H. Differential enzymatic defense mechanisms in leaves and roots of two true mangrove species under long-term salt stress. Aquat. Bot. 2017, 142, 32–40. [Google Scholar] [CrossRef]
- Lv, X.B.; Li, D.H.; Yang, X.B.; Zhang, M.W.; Deng, Q. Leaf enzyme and plant productivity responses to environmental stress associated with sea level rise in two Asian mangrove species. Forests 2019, 10, 250. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.A.; Xiao, Q.A.; Wu, F.H.; Dong, X.J.; He, J.X.; Pei, Z.M.; Zheng, H.L. Nitric oxide enhances salt secretion and Na+ sequestration in a mangrove plant, Avicennia marina, through increasing the expression of H+-ATPase and Na+/H+ antiporter under high salinity. Tree Physiol. 2010, 30, 1570–1585. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Q.; You, S.Y.; Wang, Y.B.; Huang, L.; Wang, M. Influence of frost on nutrient resorption during leaf senescence in a mangrove at its latitudinal limit of distribution. Plant Soil 2011, 342, 105–115. [Google Scholar] [CrossRef]
- Cheng, H.; Tam, N.F.Y.; Wang, Y.S.; Li, S.Y.; Chen, G.Z.; Ye, Z.H. Effect of copper on growth, radial oxygen loss and root permeability of seedlings of the mangroves Bruguiera gymnorrhiza and Rhizophora stylosa. Plant Soil 2012, 359, 255–266. [Google Scholar] [CrossRef]
- Xu, Q.T.; Yang, L.; Zhou, Z.Q.; Mei, F.Z.; Qu, L.H.; Zhou, G.S. Process of aerenchyma formation and reactive oxygen species induced by waterlogging in wheat seminal roots. Plant Soil 2013, 238, 969–982. [Google Scholar] [CrossRef]
- Dai, M.Y.; Liu, J.C.; Liu, W.W.; Lu, H.L.; Jia, H.; Hong, H.L.; Yan, C.L. Phosphorus effects on radial oxygen loss, root porosity and iron plaque in two mangrove seedlings under cadmium stress. Mar. Pollut. Bull. 2017, 119, 262–269. [Google Scholar] [CrossRef]
- Yoo, Y.H.; Choi, H.K.; Jung, K.H. Genome-wide identification and analysis of genes associated with lysigenous aerenchyma formation in rice roots. J. Plant Biol. 2015, 58, 117–127. [Google Scholar] [CrossRef]
- Yamauchi, T.; Colmer, T.D.; Pedersen, O.; Nakazono, M. Regulation of root traits for internal aeration and tolerance to soil waterlogging-flooding stress. Plant Physiol. 2018, 176, 118–1130. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Jiang, Z.Y.; Liu, Y.; Ye, Z.H.; Wu, M.L.; Sun, C.C.; Sun, F.L.; Fei, J.; Wang, Y.S. Metal (Pb, Zn and Cu) uptake and tolerance by mangroves in relation to root anatomy and lignification/suberization. Tree Physiol. 2014, 34, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Kulichikhin, K.; Yamauchi, T.; Watanabe, K.; Nakazono, M. Biochemical and molecular characterization of rice (Oryza sativa L.) roots forming a barrier to radial oxygen loss. Plant Cell Environ. 2014, 37, 2406–2420. [Google Scholar] [CrossRef]
- Kotula, L.; Schreiber, L.; Colmer, T.D.; Nakazono, M. Anatomical and biochemical characterisation of a barrier to radial O2 loss in adventitious roots of two contrasting Hordeum marinum accessions. Funct. Plant Biol. 2017, 44, 845–857. [Google Scholar] [CrossRef]
- Shiono, K.; Yamauchi, T.; Yamazaki, S.; Mohanty, B.; Malik, A.I.; Nagamura, Y.; Nishizawa, N.K.; Tsutsumi, N.; Colmer, T.D.; Nakazono, M. Microarray analysis of laser-microdissected tissues indicates the biosynthesis of suberin in the outer part of roots during formation of a barrier to radial oxygen loss in rice (Oryza sativa). J. Exp. Bot. 2014, 65, 4795–4806. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Liu, Y.; Tam, N.F.Y.; Wang, X.; Li, S.Y.; Chen, G.Z.; Ye, Z.H. The role of radial oxygen loss and root anatomy on zinc uptake and tolerance in mangrove seedlings. Environ. Pollut. 2010, 158, 1189–1196. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W. Rice: Sulfide-induced barriers to root radial oxygen loss, Fe2+ and water uptake, and lateral root emergence. Ann. Bot. 2005, 96, 625–638. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.; Armstrong, W. Rice and Phragamites: Effects of organic acids on growth, root permeability, and radial oxygen loss to the rhizosphere. Am. J. Bot. 2001, 88, 1359–1370. [Google Scholar] [CrossRef]
- Hose, E.; Clarkson, D.T.; Steudle, E.; Schreiber, L.; Hartung, W. The exodermis: A variable apoplastic barrier. J. Exp. Bot. 2001, 52, 2245–2264. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, L. Transport barriers made of cutin, suberin and associated waxes. Trends Plant Sci. 2010, 15, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, L.; Franke, R.; Hartmann, K. Effect of NO3− deficiency and NaCl stress on suberin deposition rhizo-and hypodermal and endodermal cell walls of castor bean (Ricinus communis L.) roots. Plant Soil 2005, 269, 333–339. [Google Scholar] [CrossRef]
- Ranathunge, K.; Schreiber, L.; Bi, Y.M.; Rothstein, S.J. Ammonium-induced architectural and anatomical changes with altered suberin and lignin levels significantly change water and solute permeabilities of rice (Oryza sativa L.) roots. Planta 2016, 243, 231–249. [Google Scholar] [CrossRef]
- Saengwilai, P.; Nord, E.A.; Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Root cortical aerenchyma enhances nitrogen acquisition from low-nitrogen soils in Maize. Plant Physiol. 2014, 166, 726–735. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salt Treatments (mM NaCl) | Survival (%) | Shoot Height (cm) | No. of Living Leaves | Biomass (g) |
|---|---|---|---|---|
| Control | 100 | 14.68 ± 1.03 b | 8.00 ± 0.82 a | 2.28 ± 0.21 a |
| 200 | 100 | 16.08 ± 0.16 a | 8.75 ± 0.96 a | 2.66 ± 0.07 a |
| 400 | 100 | 11.65 ± 0.53 c | 6.50 ± 1.00 b | 1.76 ± 0.09 b |
| 600 | 100 | 8.75 ± 0.72 d | 6.00 ± 1.63 b | 1.32 ± 0.11 c |
| Salt Treatments (mM NaCl) | Root Porosity (%) | ROL (µmol O2 d−1 g−1 Dry Weight root) |
|---|---|---|
| Control | 25.58 ± 1.44 a | 30.98 ± 2.50 a |
| 200 | 24.31 ± 0.81 a | 29.75 ± 1.30 a |
| 400 | 19.24 ± 1.01 b | 22.38 ± 0.63 b |
| 600 | 16.98 ± 0.96 c | 18.43 ± 1.63 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, X.; Wang, Y.; Jiang, Z.; Ma, X.; Inyang, A.I.; Cheng, H. Effects of Salt on Root Aeration, Nitrification, and Nitrogen Uptake in Mangroves. Forests 2019, 10, 1131. https://doi.org/10.3390/f10121131
Zhao Y, Wang X, Wang Y, Jiang Z, Ma X, Inyang AI, Cheng H. Effects of Salt on Root Aeration, Nitrification, and Nitrogen Uptake in Mangroves. Forests. 2019; 10(12):1131. https://doi.org/10.3390/f10121131
Chicago/Turabian StyleZhao, Yan, Xun Wang, Youshao Wang, Zhaoyu Jiang, Xiaoyu Ma, Aniefiok Ini Inyang, and Hao Cheng. 2019. "Effects of Salt on Root Aeration, Nitrification, and Nitrogen Uptake in Mangroves" Forests 10, no. 12: 1131. https://doi.org/10.3390/f10121131