1. Introduction
The mechanisms influencing nitrogen (N) deposition in wetland ecosystems are an important focus of study in research on global climate change [
1,
2]. Human activities have accelerated the deposition of N in terrestrial ecosystems and, in many regions of the world, ecosystems and wetlands, in particular, display sensitivity to reactive N [
3,
4]. Several global N deposition prediction models have estimated that the subtropical regions of China will become the most severely impacted area in terms of global atmospheric N deposition in the coming decades [
5,
6,
7]. Large amounts of N deposition alter the physicochemical properties of soil and affect plant–soil–microbial interactions [
8,
9]. Soil microbial communities play a vital role in the functioning of an ecosystem, as they are crucial for the establishment of a sustainable ecosystem that relies on healthy soil development and biological interactions [
10,
11]. In the past 30 years, although the responses of natural terrestrial ecosystems to increased N deposition have been extensively studied, the interactions within plant–soil–microbial communities under this condition are not well understood, especially in the urban wetland [
6]. Although stringent measures that reduce N emissions to maintain ecosystem structure and function have been established in the past several decades, the Yangtze River Delta Metropolitan Area remains one of the more rapidly developing areas of industrialization and urbanization and one of the main areas of N deposition in China. Its dense plain river network and lakes form urban marginal wetlands, often with typical dike-pond systems [
12]. Due to accelerated urbanization and the intensification of N deposition in the Yangtze River Delta, limited knowledge exists regarding changes in the soil microbial composition and structure in these wetlands.
Soil microorganisms can be affected by N deposition and features of the soil microhabitat, such as pH [
13,
14,
15], bulk density and water-holding capacity [
16,
17], particulate size [
18,
19], and aggregate stability [
20,
21]. However, alterations in the composition of soil microbial communities due to changes in pH and particle size as a result of N deposition are not well understood despite the importance of microbial communities in the N cycle [
15,
22,
23].
Soil pH varies both naturally and as a consequence of fertilization associated with silviculture and agronomic practices. It influences the soil microbial community structure and function due to the changes of microhabitat [
24]. There is general agreement in the scientific community regarding ecosystem acidification caused by N deposition [
15]; however, little is known about the mechanisms by which the soil microbial community copes with this acidification. A study of agricultural soils by Oehl et al. [
25] revealed that different fungal species (saprophytic, ectomycorrhizal, and arbuscular mycorrhizal fungi) respond in different ways to variations in their microhabitat, with some species being more affected by soil pH and related parameters, such as soil texture, than others. The decrease in soil pH caused by elevated N inputs was also found to reduce the activity of soil microbes [
13]. A watershed laboratory profile showed that the fungal internal transcribed spacer (ITS) was the most abundant at low pH and that bacterial 16S was the most abundant at high pH [
24]; however, on average, simulated nitrogen deposition (SND) significantly decreased the ratio of fungi-to-bacteria [
26]. N deposition also changes the composition and structure of the microbial community, and some fungi in microbial communities may be replaced by bacteria with higher N utilization [
27]. N addition was shown to have no significant effect on microbial community structure along a soil pH gradient [
27]; this may be due to the different effects of N forms and to nitrification on soil pH and, thus, fungal and bacterial composition and functions [
24,
26].
Particle-size fractionation, which allows the separation of soil organic matter with varying degrees of microbial alteration, might help elucidate microbe-mediated soil carbon (C) and N cycling characteristics [
28,
29]. Microbial acquisition of a substrate is compounded by relatively stable C and N levels [
30], high nutrient and O
2 availability [
31], and adsorption of extracellular enzymes to the clay and silt fractions but not to their aggregates [
20]. As soil changes induced by N deposition are mainly the result of microbiological processes, different substrates and soil properties are selected by specifically adapted microbial communities [
18,
29]. A prior study demonstrated that changes in microbial communities depend on their microhabitats, with a greater abundance of bacterial and fungal genes observed in the soil fractions with larger-sized particles (>63 μm) compared to silt (2–20 μm) [
28]. Poll et al. [
32] revealed that coarse sand fractions are colonized mainly by a fungus-dominated community and a rather simple bacterial community whereas silt and clay fractions are colonized by a complex bacterial community. Several studies have found that bacteria and fungi have a range of habitat requirements for soil with different particle sizes, which contain different amounts of C and N [
18,
25]. A physical separation of soils with different particle sizes is, therefore, necessary to retain their attached organic material and to ultimately provide a strategy for the characterization of conditions in different microhabitats.
Only a small number of studies have focused on the soil pH and microbial response to N deposition in urban wetlands [
2,
11]. In addition, the mechanisms of N deposition influencing soil microorganisms require more clarification. In this study, we sought to answer the following questions: (i) what are the main characteristics of soil microhabitats affected by N deposition, (ii) are the composition and structure of microbial communities affected by N deposition in the soil and, if so, which microbial species are most affected, and (iii) what are the main characteristics of microhabitat-microbial interactions that influence changes in microbial composition based on species-specific requirements following N deposition? We examined variations in the composition of the microbial community in topsoil (0–20 cm) and how they respond to soil pH and soil physical characteristics (soil water content, soil specific gravity, and bulk density) and to soil particle characteristics (clay, silt, sand, and specific surface area) under SND. Species-specific compositions of bacteria and fungi were evaluated by high-throughput sequencing of the 16S ribosomal RNA (rRNA) gene and ribosomal ITS, respectively. Overall, our study provides basic information for the forest management of N deposition and clarifies the features of species-specific microhabitats in urban wetlands.
2. Materials and Methods
2.1. Overview of the Study Area
The experimental site was Xixi National Wetland Park in the western suburb of Hangzhou, Zhejiang Province, China. Its total area is 16.15 km
2. The dike-pond is a typical ecosystem in the delta zone of the Yangtze River, where aquaculture in ponds and cultivation of economically important tree species on dikes has occurred for thousands of years, fertilizing the manure and pond sediment [
33]. The region has a monsoon climate in the northern margin of the subtropical zone. The average annual temperature in the western area of Hangzhou is 16.4 °C, the frost-free period is 240 days, and annual precipitation is 1100–1600 mm. The daily average temperature, daily average humidity, and rainfall in the Xixi Wetland begins to rise from March to May, reaches a peak in June to August, and decreases in September to October [
12]. The main soil type is paddy soil. A peat layer can be seen at an average depth of 60–100 cm.
2.2. Field Experiment and Soil Sampling
The experimental location was selected in Feijiatang Nature Reserve of the Xixi Wetland (120°03′ E, 30°15′ N). Phyllostachys violascens is the main economic species in the compound dike-pond production system of the Xixi Wetland due to its edible shoots. In recent decades, to better protect wetlands from being taken over by urban expansion, Ph. violascens forests have been left to grow naturally in the reserve. The dike width of the Ph. violascens forest ranges from 4 m to 8 m. A number of herbaceous plant species can be found in the undergrowth, including Ophiopogon japonicus, Cardamine hirsuta, Pteris multifida, Torilis scabra, Mazus japonicus, Veronica peregrina, Lygodium japonicum, Oxalis corniculata, and Digitaria violascens. The dike surface is 60–80 cm away from the water surface. The density, height, and diameter at breast height (DBH) of Ph. violascens are 2.6–3.1 stalks·m2, 3.3–5.7 m, and 2.1–4.8 cm, respectively. The light density under the forest is 73.0–89.4% of full sunlight. Initial nutrient concentrations of experimental soils such as soil organic carbon, total nitrogen, and total phosphorus are 30.01–40.57 g·kg−1, 1.28–4.07 g·kg−1, and 57.66–112.08 mg·kg−1.
Based on the actual amount of N deposition in subtropical China (30–37 kg·hm
−2 a
−1) [
34] and its increasing trend [
5,
7], four levels of SND were derived: control (CK, 0 kg·hm
−2·a
−1), low N (N30, 30 kg·hm·hm
−2·a
−1), medium N (N60, 60 kg·hm
−2·a
−1), and high N (N90, 90 kg·hm
−2·a
−1). Three 3 m × 3 m replicate plots were set up at each of the four SND levels for a total of 12 plots established in this study. There was an interval of 30–50 m between each plot as a buffer to prevent interaction.
SND spraying began in October 2016 and ended in September 2018. The annual N application rate was 12 equal applications, once per month. The specific application method was as follows: at the beginning of each month, quantitative ammonium nitrate (NH
4NO
3) was fully dissolved in 2.7 L pure water and uniformly sprayed on the surface of each plot of forest land using an electric back sprayer (equivalent to a 3 mm increase in annual precipitation). The control plot was sprayed with 2.7 L pure water without N [
7]. In October 2018, after surface litter and impurities were removed, soil samples were collected from a depth of 0–20 cm by auger boring at three randomly selected points in each plot. Subsequently, a total of 36 soil samples were collected. Soil samples were stored at 4 °C in a sample box and transported to the laboratory within 6 h of collection. The samples were sieved (2 mm) for homogenization and removal of visible roots. Three equal portions of the sieved soil samples were retrieved. One portion of the sieved sample was stored at −78 °C for DNA extraction and subsequent 16S and ITS sequencing. Another portion was air-dried for particle-size analysis, and the remainder was used for the determination of soil pH and physical parameters.
2.3. Measurement of Soil Microhabitat Properties
Soil pH was measured in a 1:2.5 (
w/
v) soil:water extract using a glass LE410 pH electrode (FiveEasy™ pH meter; Mettler-Toledo AG, Zurich, Switzerland). The particle-size distribution was measured using a Malvern Mastersizer 3000 laser diffractometer (Malvern Instruments, Malvern, UK), which enabled rapid measurement of the volume percentage of particles in 100 size classes within a 0.01–3500 μm range, giving an output of specific surface area (SSA) data. Macroscopic traces of organic matter were removed from representative subsamples before being dampened by the dropwise addition of a standard chemical solution (40 g/L solution of sodium hexametaphosphate, (NaPO
3)
6, in distilled water) for aggregate dispersal. The mixture was fully reacted by agitation for 16 h and analyzed in a deionized water suspension with no sonication. A standard range of textural parameters was calculated, including the percentage of clay (<2 μm), fine silt (FS, 2–20 μm), coarse silt (CS, 20–63 μm), and sand class sizes (>63 μm) following the taxonomy guidelines of the U.S. Department of Agriculture [
28]. The Malvern instrumentation was regularly calibrated using latex beads of known size.
The soil water content (SWC) was determined by the oven-dry weight method. The soil-specific gravity (SSG) was measured by the pycnometer method. Soil bulk density (BD) was determined by the core method with the undisturbed soil samples collected in 100 cm
3 steel cylinders and calculated as the oven-dry mass of the sample volume [
12].
2.4. DNA Extraction, Polymerase Chain Reaction, and Illumina HiSeq Sequencing
DNA was extracted using a HiPure soil DNA kit B (Magen, Guangzhou, China). The hypervariable V3/V4 regions of the bacterial 16S rRNA gene were amplified using the forward primer 5′-CCTACGGGNGGCWGCAG-3′ and reverse primer 5′-GACTACHVGGGTATCTAATCC-3′ [
35]. The ITS2 regions of the fungal rRNA gene were amplified using the forward primer 5′-GCATCGATGAAGAACGCAGC-3′ and reverse primer 5′-TCCTCCGCTTATTGATATGC-3′ [
36].
The polymerase chain reaction (PCR) was set up in a volume of 25 µL as follows: 12.5 µL of 2 × KAPA HiFi HotStart ReadyMix (KAPA Biosystems, Wilmington, MA, USA), 1 µL of each primer, 5.5 µL of PCR-grade water, and 5 µL of DNA template. The PCR conditions were as follows: initial denaturation at 95 °C for 3 min, followed by 25 cycles (30 cycles for ITS) of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s, with a final extension at 72 °C for 5 min. A specific tag sequence was added to the samples using the following PCR mixture (50 µL): 25 µL of 2 × KAPA HiFi HotStart ReadyMix, 2 µL of universal primer, 2 µL of barcode primer, 16 µL of PCR-grade water, and 5 µL of diluted template. The PCR conditions were as follows: initial denaturation at 95 °C for 3 min, followed by eight cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s, with a final extension at 72 °C for 5 min. The concentration of PCR products in each sample was measured using a NanoDrop ND1000 spectrophotometer (Wilmington, DE, USA) after gel extraction.
Library construction and Illumina HiSeq 2500 sequencing were performed by Shanghai Xiangyin Biological Technology Co., Ltd. (Shanghai, China).
2.5. Data Processing and Statistical Analysis
The obtained DNA sequence reads were trimmed using PANDAseq. After trimming, chimeras were removed using USEARCH 8.0 [
37] and sample sequences were combined using QIIME_1.9.1 software [
38]. An operational taxonomic unit (OTU) was defined as a set of DNA sequences with a similarity of >97%. OTUs were extracted using the Greengenes v13_08 database and classified by species. OTUs representing less than 0.005% of all sequences were removed before analysis. Diversity indexes were calculated using the QIIME software.
One-way ANOVA was used to test for significant differences among the different levels of SND. Multiple comparative analyses of OTU frequencies (α = 0.05) were conducted in Data Processing System (DPS) (17.10 for Windows, Zhejiang University, Hangzhou, China) using the Benjamini–Hochberg method to control the familywise error [
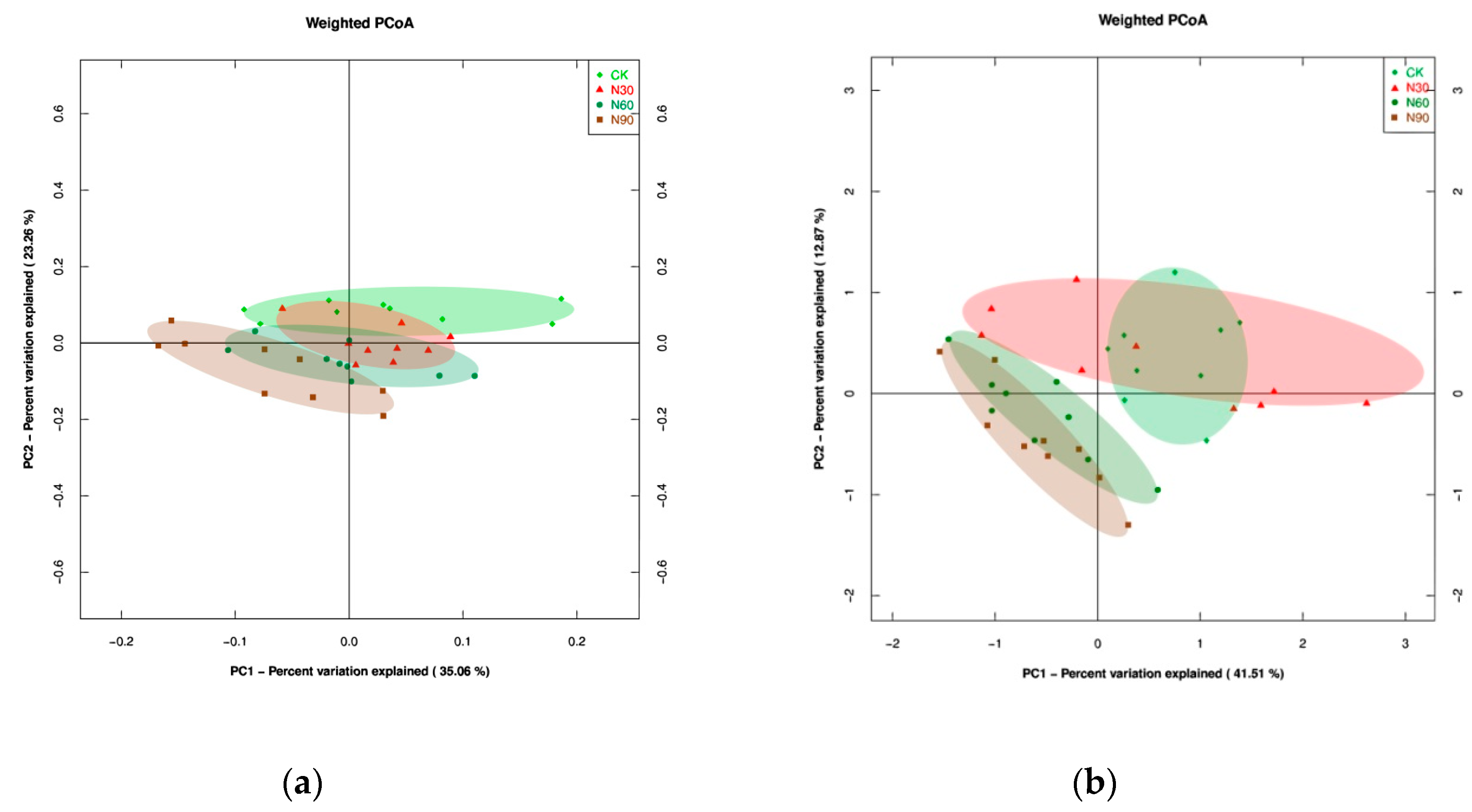
39]. Principal coordinates analysis (PCoA), redundancy analysis (RDA), nonparametric multivariate analysis of variance (PERMANOVA) of the microbial composition, as well as alpha-diversity type complexity analysis (including analysis of Chaol, Observed species (OS), and Shannon and Simpson index) were conducted at the phylum level using the “vegan” and “ggplot” package in R [
40,
41]. The fungi/bacteria ratio (F/B) was calculated based on absolute quantity (i.e., observed species) [
18]. Linear discriminant analysis coupled with effect size measurements (LefSe) was conducted using the Kruskal–Wallis rank sum test in “stats” package in R [
42].
5. Conclusions
In summary, our study showed that bacteria and fungi respond differently to SND levels, which are associated with a series of microhabitat changes (i.e., in soil pH, clay content, sand content, and SSA). Bacterial abundance in taxonomic groups below the class level declined with an increase in SND levels, with a significant difference found at the highest SND level. In contrast, the threshold of fungal responses was found to occur at a medium SND level. Species-specificity was found to be influenced by the pH value for both bacteria and fungi. Bacteria demonstrated a negative correlation with clay content and SSA, while fungi were negatively correlated with SSG. These findings suggest that the compositions of the bacterial and fungal communities had a significant interaction with different types of microaggregates. Thus, the selective effects of microhabitats induced by SND strongly contribute to niche separation. Based on these results, we recommend that the observation of microbial niche changes and indicator clades should be used as an important component in wetland forest management programs.






{kind=link}
{kind=link}
{kind=link}