Soil Carbon Storage and Its Determinants in Forest Fragments of Differentiated Patch Size

Abstract
:1. Introduction
2. Materials and Methods
3. Results
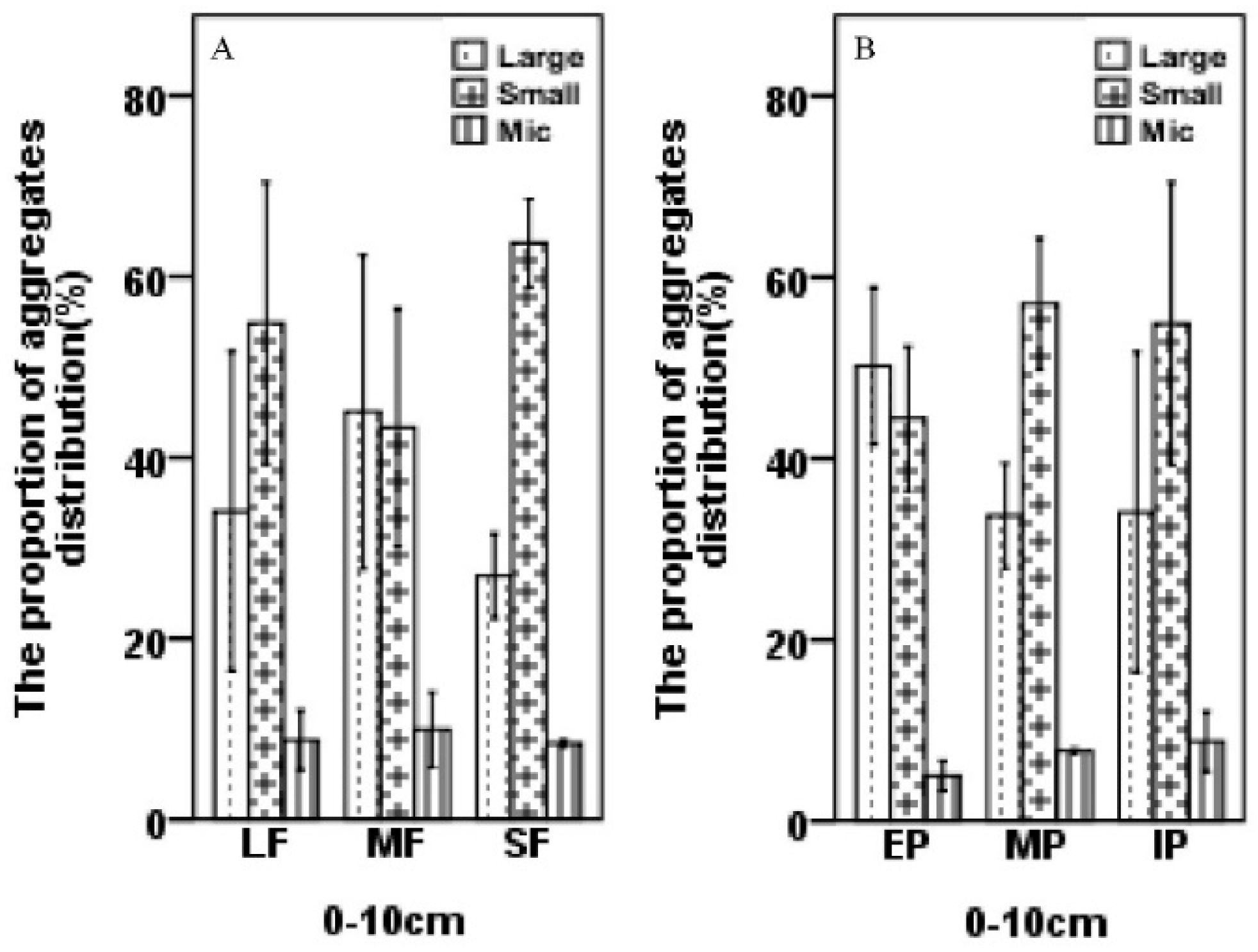
3.1. Characteristics of Three Sizes of Soil Aggregates within Forest Fragments
- The proportion of large and small macroaggregates differed among plot types.
- Microaggregates made up less than 10% of all aggregates, which was significantly lower (p < 0.05) than the proportions accounted for by small macroaggregates and large macroaggregates in all plots (Figure 1).
- The proportion of aggregates that were microaggregates increased while the proportion that were large macroaggregates decreased from edges to interior at 0–10 cm depth (Figure 1).
- The proportion of aggregates at the surface layer that were microaggregates decreased slightly as the patch size decreased.
3.2. Carbon Concentrations and Stocks in Soils
- Soil carbon concentration of all aggregates was in the order LF > MF > SF at 0–10 cm depth (Figure 2). Soil carbon concentrations of large macroaggregates (20.33 g/kg), small macroaggregates (18 g/kg), and microaggregates (16.13 g/kg) in SF were significantly (p < 0.05) lower than those in LF (58.6 g/kg, 58.27 g/kg, 62.33 g/kg, respectively) at the surface layer.
- The carbon concentration of aggregates was not different between size classes within plots (Figure 2).
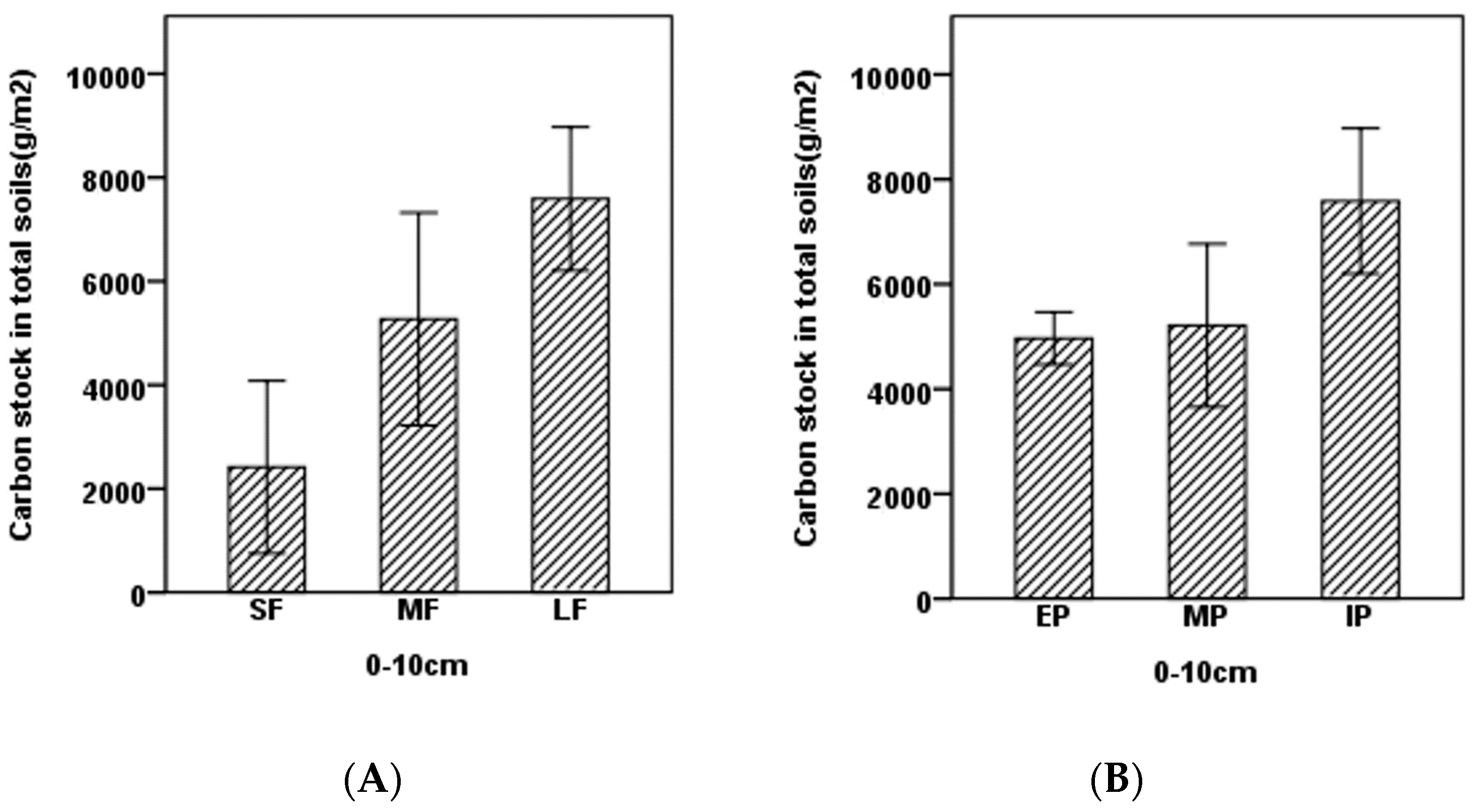
- The concentration of carbon in soils significantly increased as patch size increased and from forest edges to interiors in LF (Figure 3). The carbon concentrations increased from 18.07 g/kg to 57.3 g/kg in soils comparing SF to LF and from 33.97 g/kg to 57.07 g/kg from forest edges to interiors (Figure 3).
- Carbon concentration was significantly higher in LF than in SF, and higher in IP than in EP, respectively. The similarly trends were found in SOC of different type of plots (Figure 4).
3.3. Dynamics of Fine Roots and Litter Fall
- Fine root biomass storage was 1.62 Mg/ha in LF, which was significantly greater than that in MF (1.48 Mg/ha). Fine root production rate was 2.18, 1.43, and 1.06 Mg/ha∙y in LF, MF, and SF, respectively, during the study period (one year; Table 1).
- Fine root biomass storage and production rate decreased from LF to SF and from forest interiors to forest edges. However, decomposition rate increased from forest interiors to edges (Table 2). Similar trends were found in the dynamics of litter fall.
3.4. Contributions of Dynamics of Fine Roots and Litter Fall to Aggregate Carbon Concentration
- Fine root storage had the strongest impact on carbon concentration in large macroaggregates and microaggregates, with higher fine root biomass storage having a decreased carbon concentration. However, fine root decomposition rate had positive effects on carbon content in large macroaggregates (Table 3).
- For small macroaggregates, fine root production and decomposition rate had significantly positive effects, while fine root biomass storage, litter fall biomass storage, and litter fall decomposition had significantly negative effects on carbon concentration.
- In addition, fine root production rate had significantly positive effects on carbon concentrations of microaggregates. Generally, contributions of fine root dynamics to carbon concentration of these three aggregates were higher than litter fall dynamics.
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stockmann, U.; Padarian, J.; McBratney, A.; Minasny, B.; De Brogniez, D.; Montanarella, L.; Hong, S.Y.; Rawlins, B.G.; Field, D.J. Global soil organic carbon assessment. Glob. Food Secur. 2015, 6, 9–16. [Google Scholar] [CrossRef]
- Ye, L.; Tan, W.; Fang, L.; Ji, L.; Deng, H. Spatial analysis of soil aggregate stability in a small catchment of the Loess Plateau, China: II. Spatial prediction. Soil Tillage Res. 2019, 192, 1–11. [Google Scholar] [CrossRef]
- Gunina, A.; Kuzyakov, Y. Pathways of litter C by formation of aggregates and SOM density fractions: Implications from 13C natural abundance. Soil Biol. Biochem. 2014, 71, 95–104. [Google Scholar] [CrossRef]
- Yavitt, J.B.; Fahey, T.J.; Sherman, R.E.; Groffman, P.M. Lumbricid earthworm effects on incorporation of root and leaf litter into aggregates in a forest soil, New York State. Biogeochemistry 2015, 125, 261–273. [Google Scholar] [CrossRef]
- Xu, L.; Shi, Y.; Fang, H.; Zhou, G.; Xu, X.; Zhou, Y.; Tao, J.; Ji, B.; Xu, J.; Li, C.; et al. Vegetation carbon stocks driven by canopy density and forest age in subtropical forest ecosystems. Sci. Total Environ. 2018, 631, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Riitters, K.; Jones, K.B.; Wade, T.G.; Wickham, J.D. Distribution and Causes of Global Forest Fragmentation. Conserv. Ecol. 2003, 7, 1850–1851. [Google Scholar]
- Miller-Rushing, A.J.; Primack, R.B.; Devictor, V.; Corlett, R.T.; Cumming, G.S.; Loyola, R.; Maas, B.; Pejchar, L. How does habitat fragmentation affect biodiversity? A controversial question at the core of conservation biology. Biol. Conserv. 2019, 232, 271–273. [Google Scholar] [CrossRef]
- Song, W.; Lin, G.; Cao, H.; Ma, L. Beta diversity pattern and its driving forces of fengshui woods in South China. Bangladesh J. Bot. 2016, 45, 905–910. [Google Scholar]
- Villard, M.-A.; Metzger, J.P.; Saura, S. REVIEW: Beyond the fragmentation debate: A conceptual model to predict when habitat configuration really matters. J. Appl. Ecol. 2014, 51, 309–318. [Google Scholar] [CrossRef]
- Magnago, L.F.S.; Rocha, M.F.; Meyer, L.; Martins, S.V.; Meira-Neto, J.A.A. Microclimatic conditions at forest edges have significant impacts on vegetation structure in large Atlantic forest fragments. Biodivers. Conserv. 2015, 24, 2305–2318. [Google Scholar] [CrossRef]
- Ma, L.; Shen, C.; Lou, D.; Fu, S.; Guan, D. Patterns of ecosystem carbon density in edge-affected fengshui forests. Ecol. Eng. 2017, 107, 216–223. [Google Scholar] [CrossRef]
- Barros, H.S.; Fearnside, P.M. Soil carbon stock changes due to edge effects in central Amazon forest fragments. For. Ecol. Manag. 2016, 379, 30–36. [Google Scholar] [CrossRef]
- Baker, T.P.; Jordan, G.J.; Baker, S.C. Microclimatic edge effects in a recently harvested forest: Do remnant forest patches create the same impact as large forest areas? For. Ecol. Manag. 2016, 365, 128–136. [Google Scholar] [CrossRef]
- Pütz, S.; Groeneveld, J.; Henle, K.; Knogge, C.; Martensen, A.C.; Metz, M.; Metzger, J.P.; Ribeiro, M.C.; De Paula, M.D.; Huth, A. Long-term carbon loss in fragmented Neotropical forests. Nat. Commun. 2014, 5, 5037. [Google Scholar] [CrossRef]
- Malmivaara-Lämsä, M.; Hamberg, L.; Haapamäki, E.; Liski, J.; Kotze, D.J.; Lehvävirta, S.; Fritze, H. Edge effects and trampling in boreal urban forest fragments–Impacts on the soil microbial community. Soil Biol. Biochem. 2008, 40, 1612–1621. [Google Scholar] [CrossRef]
- Laurance, W.F.; Ferreira, L.V.; Merona, J.M.R.-D.; Laurance, S.G. Rain forest fragmentation and the dynamics of Amazonian tree communities. Ecology 1998, 79, 2032–2040. [Google Scholar] [CrossRef]
- Laurance, W.F.; Camargo, J.L.; Luizao, R.C.C.; Laurance, S.G.W.; Pimm, S.L.; Bruna, E.M.; Stouffer, P.C.; Williamson, G.B.; Benitez, J.; Vasconcelos, H.; et al. The fate of Amazonian forest fragments: A 32-year investigation. Biol. Conserv. 2013, 144, 56–67. [Google Scholar] [CrossRef]
- Ma, L.; Shen, C.; Lou, D.; Fu, S.; Guan, D. Ecosystem carbon storage in forest fragments of differing patch size. Sci. Rep. 2017, 7, 13173. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool. Soil Biol. Biochem. 2014, 68, 4–9. [Google Scholar] [CrossRef]
- Denef, K.; Zotarelli, L.; Boddey, R.M.; Six, J. Microaggregate-associated carbon as a diagnostic fraction for management-induced changes in soil organic carbon in two Oxisols. Soil Biol. Biochem. 2007, 39, 1165–1172. [Google Scholar] [CrossRef]
- Hontoria, C.; Gómez-Paccard, C.; Mariscal-Sancho, I.; Benito, M.; Pérez, J.; Espejo, R. Aggregate size distribution and associated organic C and N under different tillage systems and Ca-amendment in a degraded Ultisol. Soil Tillage Res. 2016, 160, 42–52. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, H.; Wang, X.; Chen, L.; Liu, M.; Li, H.; Sun, B. Nematodes and microbial community affect the sizes and turnover rates of organic carbon pools in soil aggregates. Soil Biol. Biochem. 2018, 119, 22–31. [Google Scholar] [CrossRef]
- Lorenz, K.; Preston, C.M.; Krumrei, S.; Feger, K.-H. Decomposition of needle/leaf litter from Scots pine, black cherry, common oak and European beech at a conurbation forest site. Eur. J. For. Res. 2004, 123, 177–188. [Google Scholar] [CrossRef]
- Hansson, K.; Helmisaari, H.-S.; Sah, S.P.; Lange, H. Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW Sweden. For. Ecol. Manag. 2013, 309, 58–65. [Google Scholar] [CrossRef]
- Sun, T.; Dong, L.; Mao, Z.; Li, Y. Fine root dynamics of trees and understorey vegetation in a chronosequence of Betula platyphylla stands. For. Ecol. Manag. 2015, 346, 1–9. [Google Scholar] [CrossRef]
- Matamala, R.; Gonzàlez-Meler, M.A.; Jastrow, J.D.; Norby, R.J.; Schlesinger, W.H. Impacts of Fine Root Turnover on Forest NPP and Soil C Sequestration Potential. Science 2003, 302, 1385–1387. [Google Scholar] [CrossRef]
- Terzaghi, M.; Di Iorio, A.; Montagnoli, A.; Baesso, B.; Scippa, G.S.; Chiatante, D. Forest canopy reduction stimulates xylem production and lowers carbon concentration in fine roots of European beech. For. Ecol. Manag. 2016, 379, 81–90. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Wang, J.; Shi, Z.; Lu, L.; Guo, W.; Jia, H.; Cai, D. Dynamics and speciation of organic carbon during decomposition of leaf litter and fine roots in four subtropical plantations of China. For. Ecol. Manag. 2013, 300, 43–52. [Google Scholar] [CrossRef]
- Tamura, M.; Suseela, V.; Simpson, M.; Powell, B.; Tharayil, N. Plant litter chemistry alters the content and composition of organic carbon associated with soil mineral and aggregate fractions in invaded ecosystems. Glob. Chang. Biol. 2017, 23, 4002–4018. [Google Scholar] [CrossRef]
- Del Giudice, R.; Lindo, Z. Short-term leaching dynamics of three peatland plant species reveals how shifts in plant communities may affect decomposition processes. Geoderma 2017, 285, 110–116. [Google Scholar] [CrossRef]
- Yan, J.; Wang, L.; Hu, Y.; Tsang, Y.F.; Zhang, Y.; Wu, J.; Fu, X.; Sun, Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma 2018, 319, 194–203. [Google Scholar] [CrossRef]
- Ma, L.; Huang, M.; Shen, Y.; Cao, H.; Wu, L.; Ye, H.; Lin, G.; Wang, Z. Species diversity and community structure in forest fragments of Guangzhou, South China. J. Trop. For. Sci. 2015, 27, 148–157. [Google Scholar]
- Katayama, A.; Kho, L.K.; Makita, N.; Kume, T.; Matsumoto, K.; Ohashi, M. Estimating Fine Root Production from Ingrowth Cores and Decomposed Roots in a Bornean Tropical Rainforest. Forests 2019, 10, 36. [Google Scholar] [CrossRef]
- Liao, Y.; McCormack, M.L.; Fan, H.; Wang, H.; Wu, J.; Tu, J.; Liu, W.; Guo, D. Relation of fine root distribution to soil C in a Cunninghamia lanceolata plantation in subtropical China. Plant Soil 2014, 381, 225–234. [Google Scholar] [CrossRef]
- R Team. R: A Language and Environmental for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Li, W.; Zheng, Z.C.; Li, T.X.; Liu, M.Y. Distribution characteristics of soil aggregates and its organic carbon in different tea plantation age. Acta Ecol. Sin. 2014, 34, 6326–6336. [Google Scholar]
- Zhao, D.; Xu, M.; Liu, G.; Ma, L.; Zhang, S.; Xiao, T.; Peng, G. Effect of vegetation type on microstructure of soil aggregates on the Loess Plateau, China. Agric. Ecosyst. Environ. 2017, 242, 1–8. [Google Scholar] [CrossRef]
- Qiu, L.; Wei, X.; Gao, J.; Zhang, X. Dynamics of soil aggregate-associated organic carbon along an afforestation chronosequence. Plant Soil 2015, 391, 237–251. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.; Paustian, K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; DeGryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Arai, M.; Miura, T.; Tsuzura, H.; Minamiya, Y.; Kaneko, N. Two-year responses of earthworm abundance, soil aggregates, and soil carbon to no-tillage and fertilization. Geoderma 2018, 332, 135–141. [Google Scholar] [CrossRef]
- Pulleman, M.; Six, J.; Uyl, A.; Marinissen, J.; Jongmans, A. Earthworms and management affect organic matter incorporation and microaggregate formation in agricultural soils. Appl. Soil Ecol. 2005, 29, 1–15. [Google Scholar] [CrossRef]
- Blazejewski, G.A.; Stolt, M.H.; Gold, A.J.; Groffman, P.M. Macro- and Micromorphology of Subsurface Carbon in Riparian Zone Soils. Soil Sci. Soc. Am. J. 2005, 69, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Liusui, Y.; Zhu, X.; Li, D.; Yan, C.; Sun, T.; Jia, H.; Zhao, X. Soil aggregate and intra-aggregate carbon fractions associated with vegetation succession in an alpine wetland of Northwest China. Catena 2019, 181, 104107. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K. Aggregate and Soil Organic Matter Dynamics under Conventional and No-Tillage Systems. Soil Sci. Soc. Am. J. 1999, 63, 1350. [Google Scholar] [CrossRef] [Green Version]
- Gentile, R.; Vanlauwe, B.; Six, J. Litter quality impacts short- but not long-term soil carbon dynamics in soil aggregate fractions. Ecol. Appl. 2011, 21, 695–703. [Google Scholar] [CrossRef]
- Zhong, X.-L.; Li, J.-T.; Li, X.-J.; Ye, Y.-C.; Liu, S.-S.; Hallett, P.D.; Ogden, M.R.; Naveed, M. Physical protection by soil aggregates stabilizes soil organic carbon under simulated N deposition in a subtropical forest of China. Geoderma 2017, 285, 323–332. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Reinmann, A.B.; Hutyra, L.R. Edge effects enhance carbon uptake and its vulnerability to climate change in temperate broadleaf forests. Proc. Natl. Acad. Sci. USA 2017, 114, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, P.; Junninen, K.; Linnakoski, R.; Edman, M.; Kouki, J. Climate and wood quality have decayer-specific effects on fungal wood decomposition. For. Ecol. Manag. 2016, 360, 341–351. [Google Scholar] [CrossRef]
- Otsing, E.; Barantal, S.; Anslan, S.; Koricheva, J.; Tedersoo, L. Litter species richness and composition effects on fungal richness and community structure in decomposing foliar and root litter. Soil Biol. Biochem. 2018, 125, 328–339. [Google Scholar] [CrossRef]
- Caplan, J.S.; Meiners, S.J.; Flores-Moreno, H.; McCormack, M.L. Fine-root traits are linked to species dynamics in a successional plant community. Ecology 2019, 100, e02588. [Google Scholar] [CrossRef] [PubMed]
- Ladegaard-Pedersen, P.; Elberling, B.; Vesterdal, L. Soil carbon stocks, mineralization rates, and CO2 effluxes under 10 tree species on contrasting soil types. Can. J. For. Res. 2005, 35, 1277–1284. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Factors causing variation in fine root biomass in forest ecosystems. For. Ecol. Manag. 2011, 261, 265–277. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, X.; Schmid, B.; Bruelheide, H.; Bu, W.; Ma, K. Positive effects of tree species richness on fine-root production in a subtropical forest in SE-China. J. Plant Ecol. 2017, 10, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Finér, L.; Domisch, T.; Dawud, S.M.; Raulund-Rasmussen, K.; Vesterdal, L.; Bouriaud, O.; Bruelheide, H.; Jaroszewicz, B.; Selvi, F.; Valladares, F. Conifer proportion explains fine root biomass more than tree species diversity and site factors in major European forest types. For. Ecol. Manag. 2017, 406, 330–350. [Google Scholar] [CrossRef]
- Wurzburger, N.; Wright, S.J. Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest. Ecology 2015, 96, 2137–2146. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Feinstein, L.M.; Kershner, M.W.; Blackwood, C.B. Aggregated and complementary: Symmetric proliferation, overyielding, and mass effects explain fine-root biomass in soil patches in a diverse temperate deciduous forest landscape. New Phytol. 2015, 205, 731–742. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Large Forest | Medium Forest | Small Forest | |
|---|---|---|---|
| Fine root biomass storage | 1.62 ± 0.53 | 1.48 ± 0.24 | 1.60 ± 0.30 |
| Fine root production rate | 2.18 ± 0.42 | 1.43 ± 0.52 | 1.06 ± 0.30 |
| Fine root decomposition rate | 42.13 ± 4.02 | 56.37 ± 1.38 | 48.49 ± 6.26 |
| Litter falls biomass storage | 508 ± 52.66 | 433 ± 42.85 | 327 ± 35.31 |
| Litter falls production rate | 640 ± 14.24 | 555 ± 24.82 | 444 ± 42.37 |
| Litter falls decomposition rate | 45.09 ± 7.38 | 31.40 ± 1.12 | 51.32 ± 12.13 |
| Inner Plots | Middle Plots | Edge Plots | |
|---|---|---|---|
| Fine root biomass storage | 1.62 ± 0.53 | 1.14 ± 0.45 | 0.88 ± 0.18 |
| Fine root production rate | 2.18 ± 0.42 | 1.47 ± 0.39 | 1.03 ± 0.11 |
| Fine root decomposition rate | 42.13 ± 4.02 | 45.25 ± 7.07 | 49.16 ± 6.11 |
| Litter falls biomass storage | 508 ± 52.66 | 426 ± 63.69 | 301 ± 75.76 |
| Litter falls production rate | 640 ± 14.24 | 526 ± 11.55 | 490 ± 28.67 |
| Litter falls decomposition rate | 45.09 ± 7.38 | 50.05 ± 11.16 | 55.24 ± 3.44 |
| Variables | LA | SA | MA | |||
|---|---|---|---|---|---|---|
| Fine root biomass storage | −0.61 | *** | −0.37 | *** | −0.51 | *** |
| Fine root production rate | 0.67 | 1.6 | *** | 1.3 | *** | |
| Fine root decomposition rate | 0.02 | ** | 0.23 | *** | 0.09 | |
| Litter falls biomass storage | 0.39 | −0.56 | *** | −0.24 | *** | |
| Litter falls production rate | 0.11 | 0.08 | 0.11 | |||
| Litter falls decomposition rate | −0.02 | −0.14 | *** | −0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, C.; Ma, L.; Hu, J.; Huang, L.; Chen, Y.; Guan, D. Soil Carbon Storage and Its Determinants in Forest Fragments of Differentiated Patch Size. Forests 2019, 10, 1044. https://doi.org/10.3390/f10111044
Shen C, Ma L, Hu J, Huang L, Chen Y, Guan D. Soil Carbon Storage and Its Determinants in Forest Fragments of Differentiated Patch Size. Forests. 2019; 10(11):1044. https://doi.org/10.3390/f10111044
Chicago/Turabian StyleShen, Chunyu, Lei Ma, Jiaxi Hu, Liyang Huang, Yujuan Chen, and Dongsheng Guan. 2019. "Soil Carbon Storage and Its Determinants in Forest Fragments of Differentiated Patch Size" Forests 10, no. 11: 1044. https://doi.org/10.3390/f10111044