Variation in Carbon Concentration and Allometric Equations for Estimating Tree Carbon Contents of 10 Broadleaf Species in Natural Forests in Northeast China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Biomass Measurements in the Field
2.3. Measurements of the Carbon Concentration and Carbon Stock
2.4. Effects of Species, Tree Sizes, and Tissues on Carbon Concentration
2.5. Additive Carbon Equations
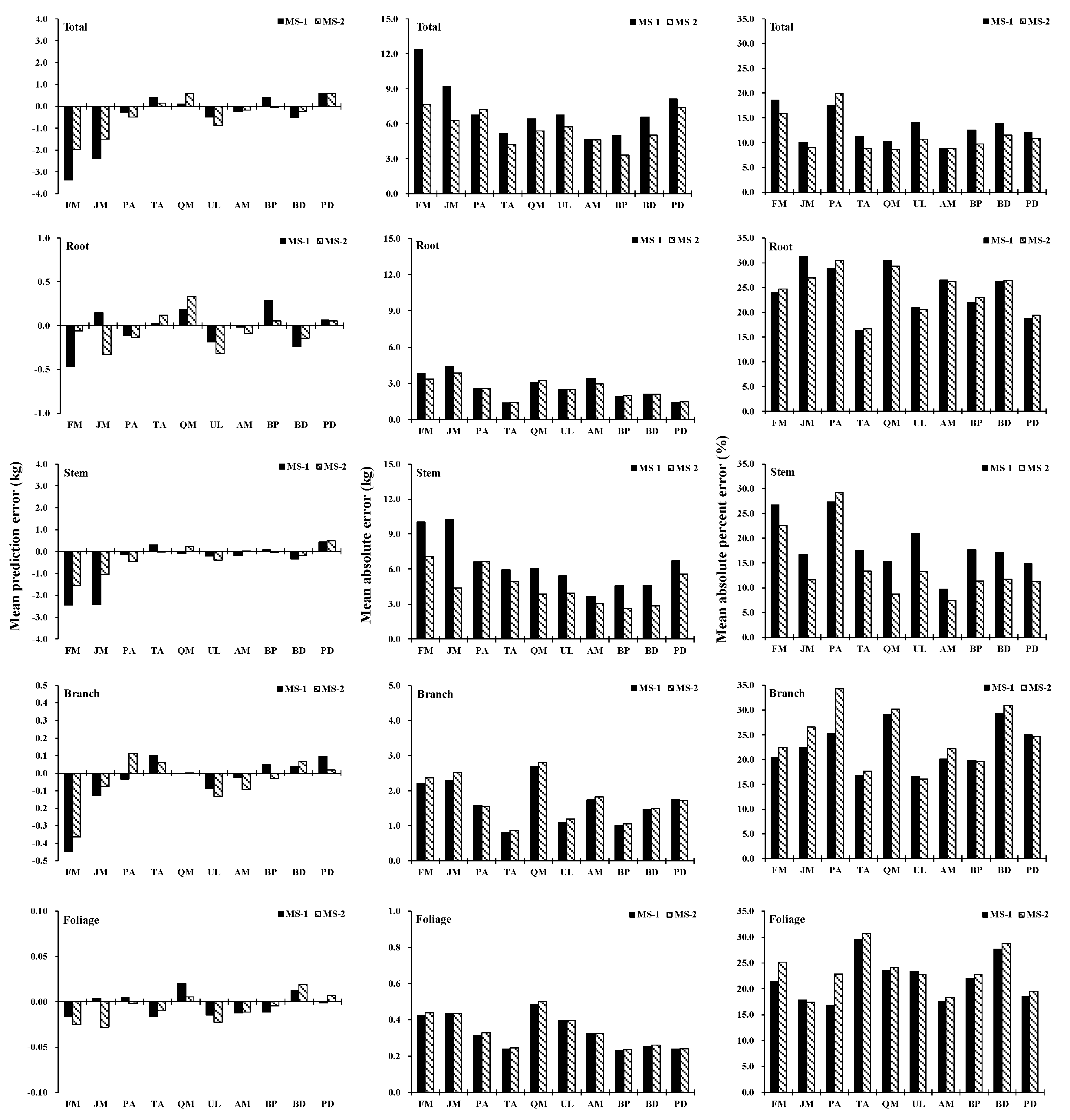
2.6. Assessment and Validation of the Model
3. Results
3.1. Variation of Carbon Concentration
3.2. Allometric Equations of Carbon and Validation of Models
3.3. Carbon Partitioning
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Statistical Yearbook: World Food and Agriculture; FAO: Rome, Italy, 2013. [Google Scholar]
- Canadell, J.G.; Le Quéré, C.; Raupach, M.R.; Field, C.B.; Buitenhuis, E.T.; Ciais, P.; Conway, T.J.; Gillett, N.P.; Houghton, R.; Marland, G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc. Natl. Acad. Sci. USA 2007, 104, 18866–18870. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Wang, C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- State Forestry Administration the Eighth Forest Resource Survey Report. 2014. Available online: http://211.167.243.162:8085/8/index.html (accessed on 14 October 2019).
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef]
- Foster, J.R.; Finley, A.O.; D’amato, A.W.; Bradford, J.B.; Banerjee, S. Predicting tree biomass growth in the temperate–boreal ecotone: Is tree size, age, competition, or climate response most important? Glob. Chang. Biol. 2016, 22, 2138–2151. [Google Scholar] [CrossRef]
- Kachamba, D.; Eid, T.; Gobakken, T. Above-and belowground biomass models for trees in the miombo woodlands of Malawi. Forests 2016, 7, 38. [Google Scholar] [CrossRef]
- Kapinga, K.; Syampungani, S.; Kasubika, R.; Yambayamba, A.M.; Shamaoma, H. Species-specific allometric models for estimation of the above-ground carbon stock in miombo woodlands of Copperbelt Province of Zambia. For. Ecol. Manag. 2018, 417, 184–196. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Allometry and partitioning of individual tree biomass and carbon of Abies nephrolepis Maxim in northeast China. Scand. J. For. Res. 2016, 31, 399–411. [Google Scholar] [CrossRef]
- Gower, S.T.; Kucharik, C.J.; Norman, J.M. Direct and indirect estimation of leaf area index, fAPAR, and net primary production of terrestrial ecosystems. Remote Sens. Environ. 1999, 70, 29–51. [Google Scholar] [CrossRef]
- Losi, C.J.; Siccama, T.G.; Condit, R.; Morales, J.E. Analysis of alternative methods for estimating carbon stock in young tropical plantations. For. Ecol. Manag. 2003, 184, 355–368. [Google Scholar] [CrossRef]
- Neumann, M.; Moreno, A.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Achten, W.M.; Thivolle-Cazat, A.; Bronisz, K. Comparison of carbon estimation methods for European forests. For. Ecol. Manag. 2016, 361, 397–420. [Google Scholar] [CrossRef]
- Houghton, R.A. Converting terrestrial ecosystems from sources to sinks of carbon. Ambio 1996, 25, 267–272. [Google Scholar]
- Gower, S.; Krankina, O.; Olson, R.; Apps, M.; Linder, S.; Wang, C. Net primary production and carbon allocation patterns of boreal forest ecosystems. Ecol. Appl. 2001, 11, 1395–1411. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C.; Wang, X.; Quan, X. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar] [CrossRef]
- Castaño-Santamaría, J.; Bravo, F. Variation in carbon concentration and basic density along stems of sessile oak (Quercus petraea (Matt.) Liebl.) and Pyrenean oak (Quercus pyrenaica Willd.) in the Cantabrian Range (NW Spain). Ann. For. Sci. 2012, 69, 663–672. [Google Scholar] [CrossRef]
- Wang, X.W.; Weng, Y.H.; Liu, G.F.; Krasowski, M.J.; Yang, C.P. Variations in carbon concentration, sequestration and partitioning among Betula platyphylla provenances. For. Ecol. Manag. 2015, 358, 344–352. [Google Scholar] [CrossRef]
- Martin, A.R.; Gezahegn, S.; Thomas, S.C. Variation in carbon and nitrogen concentration among major woody tissue types in temperate trees. Can. J. For. Res. 2015, 45, 744–757. [Google Scholar] [CrossRef]
- Gao, B.; Taylor, A.R.; Chen, H.Y.; Wang, J. Variation in total and volatile carbon concentration among the major tree species of the boreal forest. For. Ecol. Manag. 2016, 375, 191–199. [Google Scholar] [CrossRef]
- Elias, M.; Potvin, C. Assessing inter-and intra-specific variation in trunk carbon concentration for 32 neotropical tree species. Can. J. For. Res. 2003, 33, 1039–1045. [Google Scholar] [CrossRef]
- Mello, A.A.D.; Nutto, L.; Weber, K.S.; Sanquetta, C.E.; Matos, J.L.M.D.; Becker, G. Individual biomass and carbon equations for Mimosa scabrella Benth. (Bracatinga) in Southern Brazil. Silva Fenn. 2012, 46, 333–343. [Google Scholar] [CrossRef]
- Zhu, H.; Weng, Y.; Zhang, H.; Meng, F.; Major, J. Comparing fast-and slow-growing provenances of Picea koraiensis in biomass, carbon parameters and their relationships with growth. For. Ecol. Manag. 2013, 307, 178–185. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Additive biomass equations based on different dendrometric variables for two dominant species (Larix gmelini Rupr. and Betula platyphylla Suk.) in natural forests in the eastern Daxing’an Mountains, Northeast China. Forests 2018, 9, 261. [Google Scholar] [CrossRef]
- Zhao, D.; Kane, M.; Markewitz, D.; Teskey, R.; Clutter, M. Additive tree biomass equations for midrotation loblolly pine plantations. For. Sci. 2015, 61, 613–623. [Google Scholar] [CrossRef]
- Kralicek, K.; Huy, B.; Poudel, K.P.; Temesgen, H.; Salas, C. Simultaneous estimation of above-and below-ground biomass in tropical forests of Viet Nam. For. Ecol. Manag. 2017, 390, 147–156. [Google Scholar] [CrossRef]
- Zhou, X.; Brandle, J.R.; Schoeneberger, M.M.; Awada, T. Developing above-ground woody biomass equations for open-grown, multiple-stemmed tree species: Shelterbelt-grown Russian-olive. Ecol. Model. 2007, 202, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.; Murphy, S.; Volkova, L.; Weston, C.; Fairman, T.; Li, Y.; Law, R.; Norris, J.; Lei, X.; Caccamo, G. Additive biomass equations based on complete weighing of sample trees for open eucalypt forest species in south-eastern Australia. For. Ecol. Manag. 2015, 349, 106–121. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. A three-step proportional weighting system of nonlinear biomass equations. For. Sci. 2015, 61, 35–45. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar]
- Parresol, B.R. Additivity of nonlinear biomass equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Tang, S.; Li, Y.; Wang, Y. Simultaneous equations, error-in-variable models, and model integration in systems ecology. Ecol. Model. 2001, 142, 285–294. [Google Scholar] [CrossRef]
- Bi, H.; Turner, J.; Lambert, M.J. Additive biomass equations for native eucalypt forest trees of temperate Australia. Trees 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Zhao, D.; Westfall, J.; Coulston, J.W.; Lynch, T.B.; Bullock, B.P.; Montes, C.R. Additive biomass equations for slash pine trees: Comparing three modeling approaches. Can. J. For. Res. 2019, 49, 27–40. [Google Scholar] [CrossRef]
- Finney, D. On the distribution of a variate whose logarithm is normally distributed. Suppl. J. R. Stat. Soc. 1941, 7, 155–161. [Google Scholar] [CrossRef]
- Baskerville, G. Use of logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Clifford, D.; Cressie, N.; England, J.R.; Roxburgh, S.H.; Paul, K.I. Correction factors for unbiased, efficient estimation and prediction of biomass from log–log allometric models. For. Ecol. Manag. 2013, 310, 375–381. [Google Scholar] [CrossRef]
- Zianis, D.; Xanthopoulos, G.; Kalabokidis, K.; Kazakis, G.; Ghosn, D.; Roussou, O. Allometric equations for aboveground biomass estimation by size class for Pinus brutia Ten. Trees growing in North and South Aegean Islands, Greece. Eur. J. For. Res. 2011, 130, 145–160. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. A compatible system of biomass equations for three conifer species in Northeast, China. For. Ecol. Manag. 2014, 329, 306–317. [Google Scholar] [CrossRef]
- Meng, S.; Liu, Q.; Zhou, G.; Jia, Q.; Zhuang, H.; Zhou, H. Aboveground tree additive biomass equations for two dominant deciduous tree species in Daxing’anling, northernmost China. J. For. Res. 2017, 22, 233–240. [Google Scholar] [CrossRef]
- Paul, K.I.; Radtke, P.J.; Roxburgh, S.H.; Larmour, J.; Waterworth, R.; Butler, D.; Brooksbank, K.; Ximenes, F. Validation of allometric biomass models: How to have confidence in the application of existing models. For. Ecol. Manag. 2018, 412, 70–79. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/ETS 9.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Fu, L.; Sun, W.; Wang, G. A climate-sensitive aboveground biomass model for three larch species in northeastern and northern China. Trees 2017, 31, 557–573. [Google Scholar] [CrossRef]
- Zeng, W.; Duo, H.; Lei, X.; Chen, X.; Wang, X.; Pu, Y.; Zou, W. Individual tree biomass equations and growth models sensitive to climate variables for Larix spp. in China. Eur. J. For. Res. 2017, 136, 233–249. [Google Scholar] [CrossRef]
- Thomas, S.; Malczewski, G. Wood carbon content of tree species in Eastern China: Interspecific variability and the importance of the volatile fraction. J. Environ. Manag. 2007, 85, 659–662. [Google Scholar] [CrossRef]
- Lamlom, S.; Savidge, R. A reassessment of carbon content in wood: Variation within and between 41 North American species. Biomass Bioenergy 2003, 25, 381–388. [Google Scholar] [CrossRef]
- Ngo, K.M.; Turner, B.L.; Muller-Landau, H.C.; Davies, S.J.; Larjavaara, M.; bin Nik Hassan, N.F.; Lum, S. Carbon stocks in primary and secondary tropical forests in Singapore. For. Ecol. Manag. 2013, 296, 81–89. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, F.; Liu, S.; Liu, Y.; Li, C. Variations of carbon stock with forest types in subalpine region of southwestern China. For. Ecol. Manag. 2013, 300, 88–95. [Google Scholar] [CrossRef]
- Diédhiou, I.; Diallo, D.; Mbengue, A.; Hernandez, R.; Bayala, R.; Diémé, R.; Diédhiou, P.; Sène, A. Allometric equations and carbon stocks in tree biomass of Jatropha curcas L. in Senegal’s Peanut Basin. Glob. Ecol. Conserv. 2017, 9, 61–69. [Google Scholar] [CrossRef]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Strong, W.; Roi, G.L. Root-system morphology of common boreal forest trees in Alberta, Canada. Can. J. For. Res. 1983, 13, 1164–1173. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.; Ehleringer, J.; Mooney, H.; Sala, O.; Schulze, E.-D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | DF | Type III SS | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Tree size (D) | 1 | 24.16 | 24.16 | 5.42 | 0.0200 |
| Tree species | 9 | 603.19 | 67.02 | 15.05 | <0.0001 |
| Tissue | 3 | 854.96 | 284.99 | 63.99 | <0.0001 |
| Tree size × Tissue | 9 | 273.65 | 30.41 | 6.83 | <0.0001 |
| Tree Species | N | Branch | Foliage | Root | Stem | WMCC |
|---|---|---|---|---|---|---|
| FM | 24 | 45.70 ± 2.83 | 44.49 ± 1.91 | 44.11 ± 2.92 | 44.82 ± 3.06 | 44.75 ± 2.93 |
| JM | 30 | 45.05 ± 1.90 | 46.85 ± 1.93 | 42.89 ± 1.69 | 44.95 ± 2.46 | 44.58 ± 2.04 |
| PA | 18 | 43.63 ± 2.31 | 43.67 ± 1.76 | 42.47 ± 2.88 | 44.16 ± 2.22 | 43.70 ± 2.25 |
| TA | 38 | 43.97 ± 2.43 | 45.24 ± 2.30 | 43.57 ± 2.39 | 45.18 ± 2.25 | 44.73 ± 2.06 |
| QM | 64 | 44.91 ± 2.10 | 46.70 ± 2.12 | 44.06 ± 2.36 | 45.68 ± 2.13 | 45.25 ± 2.02 |
| UL | 40 | 44.26 ± 1.53 | 42.87 ± 1.54 | 43.07 ± 1.69 | 43.85 ± 1.91 | 43.67 ± 1.62 |
| AM | 46 | 44.07 ± 2.27 | 44.37 ± 2.02 | 43.19 ± 1.89 | 44.20 ± 2.25 | 43.94 ± 2.01 |
| BP | 66 | 46.17 ± 1.77 | 48.68 ± 2.09 | 45.46 ± 1.77 | 46.35 ± 1.87 | 46.18 ± 1.64 |
| BD | 52 | 45.92 ± 1.85 | 46.43 ± 2.04 | 44.99 ± 1.94 | 45.70 ± 2.09 | 45.56 ± 1.91 |
| PD | 54 | 44.53 ± 1.99 | 45.92 ± 2.46 | 43.37 ± 2.03 | 44.40 ± 1.88 | 44.28 ± 1.81 |
| All species | 432 | 44.94 ± 2.20 | 45.90 ± 2.66 | 43.93 ± 2.27 | 45.09 ± 2.28 | 44.84 ± 2.11 |
| Tree Species | Carbon Components | βi0 | βi1 | RMSE | Weight Function | |||
|---|---|---|---|---|---|---|---|---|
| Estimate | SE | Estimate | SE | |||||
| FM | Root | −4.3993 | 0.3836 | 2.5020 | 0.1221 | 0.9268 | 4.5548 | D2.4537 |
| Stem | −2.2940 | 0.2322 | 2.1752 | 0.0753 | 0.9150 | 12.9900 | D2.2577 | |
| Branch | −6.2638 | 0.3550 | 2.9343 | 0.1114 | 0.9385 | 2.7533 | D2.8112 | |
| Foliage | −5.3096 | 0.4059 | 2.1160 | 0.1308 | 0.9307 | 0.5116 | D2.1436 | |
| Total | - | - | - | - | 0.9431 | 17.4863 | D2.2123 | |
| JM | Root | −3.4686 | 0.3393 | 2.0564 | 0.1068 | 0.8948 | 5.5046 | D2.9440 |
| Stem | −3.6363 | 0.1651 | 2.5117 | 0.0547 | 0.9539 | 13.9442 | D3.8552 | |
| Branch | −4.2657 | 0.2768 | 2.2587 | 0.0839 | 0.9549 | 3.0605 | D1.9283 | |
| Foliage | −5.5766 | 0.2931 | 2.1833 | 0.0930 | 0.9677 | 0.5337 | D3.2885 | |
| Total | - | - | - | - | 0.9808 | 13.4931 | D3.4722 | |
| PA | Root | −6.4318 | 0.4733 | 3.0452 | 0.1441 | 0.9766 | 2.9545 | D2.0757 |
| Stem | −3.3025 | 0.1385 | 2.3845 | 0.0417 | 0.9756 | 7.1162 | D2.8811 | |
| Branch | −6.2062 | 0.4173 | 2.8708 | 0.1273 | 0.9806 | 1.8019 | D2.0720 | |
| Foliage | −5.7706 | 0.4033 | 2.2266 | 0.1265 | 0.9644 | 0.4015 | D2.7516 | |
| Total | - | - | - | - | 0.9895 | 8.1229 | D2.9551 | |
| TA | Root | −3.2098 | 0.2114 | 1.9424 | 0.0721 | 0.9720 | 1.7613 | D2.8887 |
| Stem | −3.5676 | 0.1580 | 2.4640 | 0.0501 | 0.9686 | 8.2212 | D2.4344 | |
| Branch | −5.7017 | 0.2577 | 2.5094 | 0.0853 | 0.9663 | 1.3210 | D3.6814 | |
| Foliage | −5.1279 | 0.3364 | 1.8247 | 0.1125 | 0.8780 | 0.3153 | D2.3317 | |
| Total | - | - | - | - | 0.9870 | 7.3417 | D1.9310 | |
| QM | Root | −4.1592 | 0.1892 | 2.3883 | 0.0621 | 0.9555 | 4.3850 | D3.6476 |
| Stem | −3.0136 | 0.1422 | 2.3729 | 0.0451 | 0.9785 | 8.5595 | D2.7019 | |
| Branch | −6.6852 | 0.2577 | 3.1627 | 0.0797 | 0.9759 | 3.8246 | D3.7733 | |
| Foliage | −6.6988 | 0.2607 | 2.5843 | 0.0802 | 0.9489 | 0.7517 | D2.1625 | |
| Total | - | - | - | - | 0.9922 | 9.3531 | D2.5420 | |
| UL | Root | −3.2591 | 0.1909 | 2.0468 | 0.0643 | 0.9446 | 3.3534 | D2.8270 |
| Stem | −2.6275 | 0.1185 | 2.1730 | 0.0374 | 0.9703 | 7.2734 | D2.7464 | |
| Branch | −3.2156 | 0.1607 | 1.8316 | 0.0535 | 0.9567 | 1.3939 | D2.6379 | |
| Foliage | −3.9191 | 0.2446 | 1.6018 | 0.0844 | 0.8991 | 0.4876 | D2.6514 | |
| Total | - | - | - | - | 0.9805 | 8.9256 | D2.8154 | |
| AM | Root | −4.8306 | 0.3060 | 2.6609 | 0.0965 | 0.9558 | 4.0845 | D2.1791 |
| Stem | −2.8834 | 0.1263 | 2.3046 | 0.0409 | 0.9817 | 5.3065 | D2.1708 | |
| Branch | −4.2090 | 0.2139 | 2.3003 | 0.0724 | 0.9483 | 2.2505 | D2.8763 | |
| Foliage | −4.2266 | 0.1870 | 1.7472 | 0.0663 | 0.9218 | 0.4071 | D2.9753 | |
| Total | - | - | - | - | 0.9905 | 6.7462 | D2.4949 | |
| BP | Root | −4.0412 | 0.1659 | 2.3718 | 0.0583 | 0.9637 | 2.9315 | D3.6368 |
| Stem | −2.7296 | 0.1158 | 2.2856 | 0.0407 | 0.9644 | 6.9291 | D2.8397 | |
| Branch | −6.0092 | 0.2256 | 2.8747 | 0.0760 | 0.9798 | 1.5945 | D3.6122 | |
| Foliage | −6.3597 | 0.1641 | 2.4766 | 0.0566 | 0.9714 | 0.3290 | D3.0989 | |
| Total | - | - | - | - | 0.9876 | 7.1944 | D2.3971 | |
| BD | Root | −3.8799 | 0.1525 | 2.2312 | 0.0518 | 0.9108 | 3.2188 | D2.2107 |
| Stem | −3.1879 | 0.1703 | 2.4001 | 0.0599 | 0.9603 | 6.8378 | D3.6068 | |
| Branch | −8.3881 | 0.3189 | 3.6647 | 0.1025 | 0.9659 | 2.5285 | D3.6079 | |
| Foliage | −8.0584 | 0.2529 | 3.0287 | 0.0799 | 0.9793 | 0.3108 | D1.4856 | |
| Total | - | - | - | - | 0.9715 | 10.1206 | D3.6981 | |
| PD | Root | −4.3300 | 0.2395 | 2.2614 | 0.0762 | 0.9606 | 1.8136 | D2.2476 |
| Stem | −2.8292 | 0.1402 | 2.2754 | 0.0463 | 0.9563 | 8.8689 | D2.9923 | |
| Branch | −7.5074 | 0.3793 | 3.1670 | 0.1185 | 0.9420 | 2.2657 | D3.9500 | |
| Foliage | −6.8948 | 0.2619 | 2.4573 | 0.0824 | 0.9300 | 0.3662 | D2.2230 | |
| Total | - | - | - | - | 0.9673 | 11.1664 | D2.5572 | |
| Tree Species | Carbon Components | βi0 | βi1 | βi2 | RMSE | Weight Function | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SE | Estimate | SE | Estimate | SE | |||||
| FM | Root | −3.9956 | 0.6335 | 2.2747 | 0.1376 | 0.1004 | 0.2629 | 0.9443 | 3.9741 | D2.0463 |
| Stem | −3.2245 | 0.3059 | 1.6765 | 0.0607 | 0.8291 | 0.1256 | 0.9706 | 7.6390 | D1.9876 | |
| Branch | −7.3358 | 0.5465 | 2.8620 | 0.1254 | 0.4330 | 0.2388 | 0.9372 | 2.7832 | D2.7448 | |
| Foliage | −4.5477 | 0.6954 | 2.0263 | 0.1575 | −0.1628 | 0.2959 | 0.9344 | 0.4978 | D1.8113 | |
| Total | - | - | - | - | - | - | 0.9804 | 10.2714 | D2.1491 | |
| JM | Root | −3.0664 | 0.4009 | 2.5876 | 0.1889 | −0.7066 | 0.2311 | 0.9097 | 5.0990 | D2.6508 |
| Stem | −3.9598 | 0.1280 | 1.8806 | 0.0593 | 0.7856 | 0.0707 | 0.9899 | 6.5391 | D2.8925 | |
| Branch | −3.8308 | 0.2301 | 2.2356 | 0.1147 | −0.1199 | 0.1394 | 0.9585 | 2.9341 | D1.7883 | |
| Foliage | −5.4919 | 0.3287 | 2.3766 | 0.1565 | −0.2361 | 0.1906 | 0.9679 | 0.5322 | D2.7763 | |
| Total | - | - | - | - | - | - | 0.9915 | 8.9693 | D2.6857 | |
| PA | Root | −6.2320 | 0.6971 | 2.9456 | 0.2139 | 0.0434 | 0.3526 | 0.9764 | 2.9710 | D2.2485 |
| Stem | −3.0940 | 0.2925 | 2.4544 | 0.0915 | −0.1513 | 0.1472 | 0.9755 | 7.1248 | D2.2018 | |
| Branch | −5.6096 | 0.6007 | 2.7677 | 0.1845 | −0.0899 | 0.3022 | 0.9829 | 1.6899 | D1.8890 | |
| Foliage | −4.8826 | 0.4613 | 2.3390 | 0.2023 | −0.4366 | 0.3015 | 0.9702 | 0.3678 | D1.9366 | |
| Total | - | - | - | - | - | - | 0.9880 | 8.6846 | D2.3215 | |
| TA | Root | −3.3346 | 0.3365 | 1.8780 | 0.1274 | 0.1138 | 0.2220 | 0.9699 | 1.8267 | D3.0437 |
| Stem | −4.5319 | 0.2987 | 2.1628 | 0.0888 | 0.6881 | 0.1772 | 0.9774 | 6.9783 | D2.4724 | |
| Branch | −5.6928 | 0.4856 | 2.5542 | 0.1647 | −0.0513 | 0.3005 | 0.9686 | 1.2752 | D3.2538 | |
| Foliage | −5.0719 | 0.5696 | 1.8170 | 0.2026 | −0.0126 | 0.3619 | 0.8808 | 0.3117 | D2.2758 | |
| Total | - | - | - | - | - | - | 0.9906 | 6.2356 | D2.0373 | |
| QM | Root | −3.8662 | 0.2290 | 2.5715 | 0.0990 | −0.3216 | 0.1586 | 0.9561 | 4.3558 | D2.9477 |
| Stem | −3.9306 | 0.1226 | 2.0347 | 0.0426 | 0.7199 | 0.0695 | 0.9894 | 6.0027 | D2.7199 | |
| Branch | −6.6321 | 0.3426 | 3.1306 | 0.1060 | 0.0172 | 0.1639 | 0.9756 | 3.8468 | D2.7882 | |
| Foliage | −6.6655 | 0.3530 | 2.6626 | 0.1117 | −0.1021 | 0.1702 | 0.9507 | 0.7381 | D1.8370 | |
| Total | - | - | - | - | - | - | 0.9943 | 7.9764 | D2.5228 | |
| UL | Root | −3.4129 | 0.2592 | 2.1852 | 0.1136 | −0.0981 | 0.1772 | 0.9414 | 3.4469 | D2.5519 |
| Stem | −3.8518 | 0.1737 | 1.9719 | 0.0592 | 0.6701 | 0.0903 | 0.9834 | 5.4327 | D2.3251 | |
| Branch | −3.2943 | 0.2284 | 1.9281 | 0.0923 | −0.0789 | 0.1423 | 0.9503 | 1.4939 | D2.2346 | |
| Foliage | −3.8016 | 0.3880 | 1.7543 | 0.1647 | −0.2134 | 0.2565 | 0.8935 | 0.5008 | D2.1660 | |
| Total | - | - | - | - | - | - | 0.9838 | 8.1408 | D2.2144 | |
| AM | Root | −3.9510 | 0.2955 | 2.7922 | 0.0853 | −0.4747 | 0.1223 | 0.9641 | 3.6834 | D1.7510 |
| Stem | −3.6194 | 0.1403 | 2.1589 | 0.0486 | 0.4375 | 0.0827 | 0.9867 | 4.5312 | D2.6590 | |
| Branch | −3.9286 | 0.3449 | 2.2380 | 0.1104 | −0.0329 | 0.1789 | 0.9530 | 2.1452 | D2.0332 | |
| Foliage | −4.2369 | 0.3343 | 1.6296 | 0.1183 | 0.1351 | 0.2048 | 0.9270 | 0.3935 | D2.2499 | |
| Total | - | - | - | - | - | - | 0.9906 | 6.7104 | D2.3593 | |
| BP | Root | −4.0713 | 0.4800 | 2.3894 | 0.1698 | −0.0005 | 0.2997 | 0.9668 | 2.8031 | D3.2843 |
| Stem | −4.1802 | 0.1955 | 1.7812 | 0.0583 | 1.0230 | 0.1105 | 0.9902 | 3.6256 | D2.5802 | |
| Branch | −5.9972 | 0.7607 | 2.9277 | 0.2163 | −0.0561 | 0.4189 | 0.9788 | 1.6346 | D3.5408 | |
| Foliage | −6.1326 | 0.3590 | 2.4996 | 0.0978 | −0.1040 | 0.1938 | 0.9727 | 0.3215 | D2.2578 | |
| Total | - | - | - | - | - | - | 0.9953 | 4.4225 | D2.1757 | |
| BD | Root | −4.0287 | 0.2430 | 2.2069 | 0.1334 | 0.0778 | 0.1857 | 0.9122 | 3.1936 | D2.3814 |
| Stem | −4.1736 | 0.1466 | 1.8585 | 0.0614 | 0.9411 | 0.0871 | 0.9864 | 3.9996 | D3.1062 | |
| Branch | −8.6425 | 0.4104 | 3.7298 | 0.1669 | 0.0178 | 0.2210 | 0.9692 | 2.4030 | D3.3696 | |
| Foliage | −8.1679 | 0.3071 | 3.0751 | 0.1215 | −0.0133 | 0.1593 | 0.9799 | 0.3064 | D1.2775 | |
| Total | - | - | - | - | - | - | 0.9826 | 7.9073 | D3.3496 | |
| PD | Root | −4.3908 | 0.4537 | 2.1979 | 0.1090 | 0.0875 | 0.2109 | 0.9607 | 1.8118 | D2.1666 |
| Stem | −4.1757 | 0.2161 | 1.9245 | 0.0594 | 0.8179 | 0.1128 | 0.9687 | 7.5064 | D2.9923 | |
| Branch | −7.0421 | 0.8779 | 3.3141 | 0.2075 | −0.3094 | 0.4057 | 0.9426 | 2.2546 | D3.2213 | |
| Foliage | −6.1852 | 0.4526 | 2.5739 | 0.1114 | −0.3613 | 0.2111 | 0.9338 | 0.3562 | D2.1713 | |
| Total | - | - | - | - | - | - | 0.9722 | 10.3088 | D2.1058 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Liu, Y.; Zhang, L.; Xie, L.; Li, F. Variation in Carbon Concentration and Allometric Equations for Estimating Tree Carbon Contents of 10 Broadleaf Species in Natural Forests in Northeast China. Forests 2019, 10, 928. https://doi.org/10.3390/f10100928
Dong L, Liu Y, Zhang L, Xie L, Li F. Variation in Carbon Concentration and Allometric Equations for Estimating Tree Carbon Contents of 10 Broadleaf Species in Natural Forests in Northeast China. Forests. 2019; 10(10):928. https://doi.org/10.3390/f10100928
Chicago/Turabian StyleDong, Lihu, Yongshuai Liu, Lianjun Zhang, Longfei Xie, and Fengri Li. 2019. "Variation in Carbon Concentration and Allometric Equations for Estimating Tree Carbon Contents of 10 Broadleaf Species in Natural Forests in Northeast China" Forests 10, no. 10: 928. https://doi.org/10.3390/f10100928