Dry Season Irrigation Promotes Leaf Growth in Eucalyptus urophylla × E. grandis under Fertilization

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Plant Material
2.2. Experimental Method
2.3. Index Measure
2.4. Statistical Analysis
3. Results
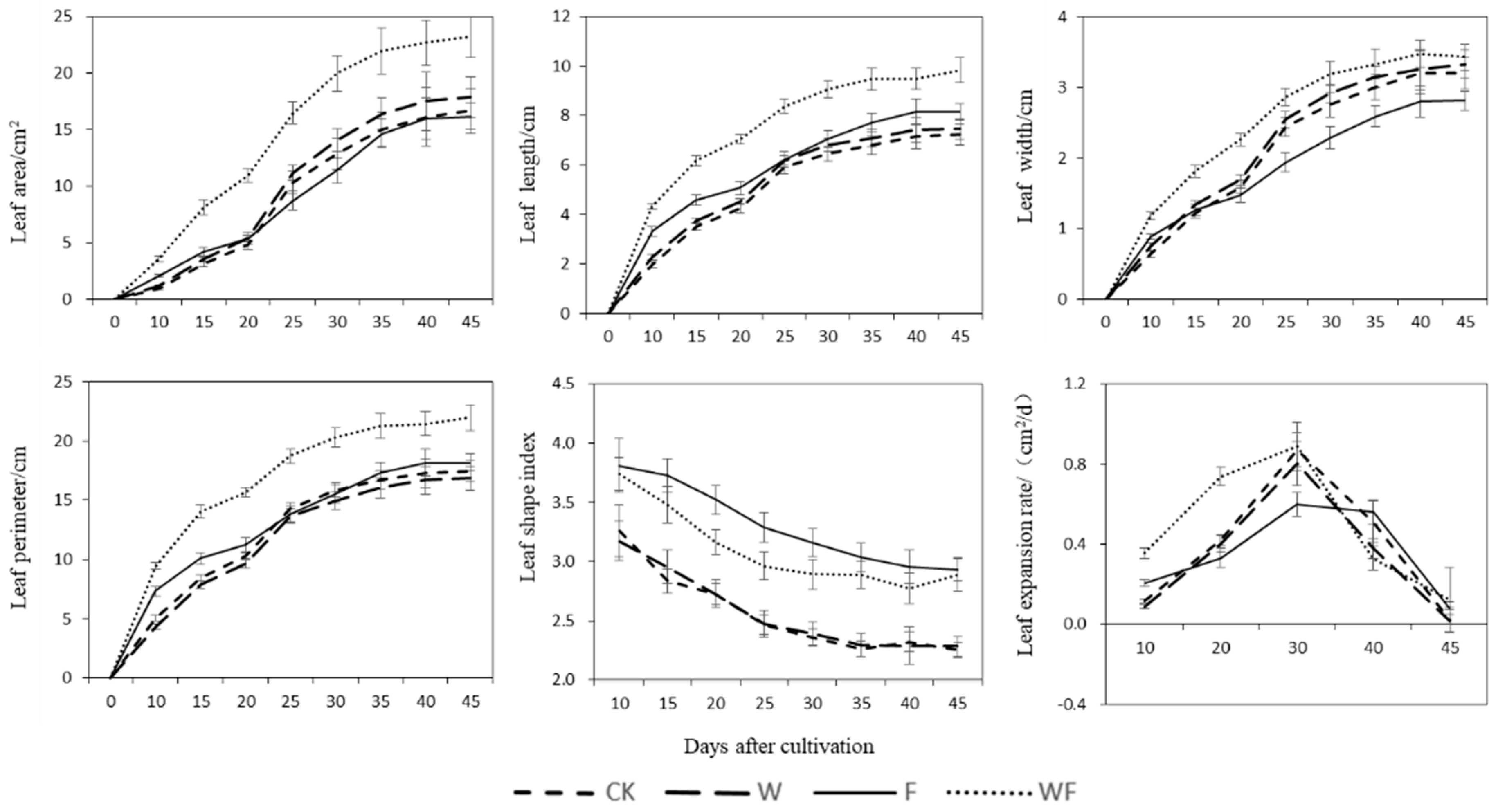
3.1. Changes in Leaf Growth Trend
3.2. Two-Factor Analysis of the Leaf Index in Dry Season Irrigation and Fertilization
3.3. Leaf Logistic Model Fitting
4. Discussion
4.1. Effects of Water and Fertilizer on Leaf Growth
4.2. Leaf Shape Variation
4.3. Leaf Expansion Rate and Model Fitting
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavenderbares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821. [Google Scholar] [CrossRef]
- Grier, C.G.; Running, S.W. Leaf area of mature northwestern coniferous forests: Relation to site water balance. Ecology 1977, 58, 893–899. [Google Scholar] [CrossRef]
- Wellstein, C.; Poschlod, P.; Gohlke, A.; Chelli, S.; Campetella, G.; Rosbakh, S.; Canullo, R.; Kreyling, J.; Jentsch, A.; Beierkuhnlein, C. Effects of extreme drought on specific leaf area of grassland species: A meta-analysis of experimental studies in temperate and sub-Mediterranean systems. Glob. Chang. Biol. 2017, 541, 516. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N. Shaping up: The genetic control of leaf shape. Curr. Opin. Plant Biol. 2004, 7, 65–72. [Google Scholar]
- Barkoulas, M.; Galinha, C.; Grigg, S.P.; Tsiantis, M. From genes to shape: Regulatory interactions in leaf development. Curr. Opin. Plant Biol. 2007, 10, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Traiser, C.; Klotz, S.; Uhl, D.; Mosbrugger, V. Environmental signals from leaves—A physiognomic analysis of European vegetation. New Phytol. 2005, 166, 465–484. [Google Scholar] [CrossRef] [PubMed]
- Roche, P.; Díaz-Burlinson, N.; Gachet, S. Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? Plant Ecol. 2004, 174, 37–48. [Google Scholar] [CrossRef]
- Yang, H.Y.; Wei, H.Y.; Sang, M.J.; Shang, Z.H.; Mao, Y.J.; Wang, X.R.; Liu, F.; Gu, W. Phenotypic Plasticity of Schisandra sphenanthera Leaf and the Effect of Environmental Factors on Leaf Phenotype. Chin. Bull. Bot. 2016, 51, 322–334. [Google Scholar]
- Nicotra, A.B.; Cosgrove, M.J.; Cowling, A.; Schlichting, C.D.; Jones, C.S. Leaf Shape Linked to Photosynthetic Rates and Temperature Optima in South African Pelargonium Species. Oecologia 2008, 154, 625–635. [Google Scholar] [CrossRef]
- Wang, C.; Liu, J.; Xiao, H.; Du, D. Response of Leaf Functional Traits of Cerasus yedoensis (Mats.) Yü Li to Serious Insect Attack. Pol. J. Environm. Stud. 2016, 25, 333–339. [Google Scholar] [CrossRef]
- Martinez, K.A.; Fridley, J.D. Acclimation of leaf traits in seasonal light environments: Are non-native species more plastic? J. Ecol. 2018. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Penuelas, J.L.; Jacobs, D.F. Nitrogen nutrition and drought hardening exert opposite effects on the stress tolerance of Pinus pinea L. seedlings. Tree Physiol. 2013, 33, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Y.; Wang, M.; Yang, D.; Song, J.; Zhang, W.W.; Han, S.J.; Hao, G.Y. Responses of hydraulics at the whole-plant level to simulated nitrogen deposition of different levels in Fraxinus mandshurica. Tree Physiol. 2016, 36, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Albaugh, T.J.; Allen, H.L.; Dougherty, P.M.; Kress, L.W.; King, J.S. Leaf area and above- and belowground growth responses of loblolly pine to nutrient and water additions. For. Sci. 1998, 44, 317–328. [Google Scholar]
- Lower, S.S.; Orians, C.M. Soil nutrients and water availability interact to influence willow growth and chemistry but not leaf beetle performance. Entomol. Exp. Appl. 2003, 107, 69–79. [Google Scholar] [CrossRef]
- Maire, G.L.; Marsden, C.; Verhoef, W.; Ponzoni, F.J.; Seen, D.L.; Bégué, A.; Stape, J.; Nouvellon, Y. Leaf area index estimation with MODIS reflectance time series and model inversion during full rotations of Eucalyptus, plantations. Remote Sens. Environ. 2011, 115, 586–599. [Google Scholar] [CrossRef]
- Macfarlane, C.; Coote, M.; White, D.A.; Adams, M.A. Photographic exposure affects indirect estimation of leaf area in plantations of Eucalyptus globulus Labill. Agric. For. Meteorol. 2000, 100, 155–168. [Google Scholar] [CrossRef]
- Diao, J.; Lei, X.D.; Hong, L.X.; Kong, J.T.; Qiang, S. Single leaf area estimation models based on the leaf weight of Eucalyptus in southern china. J. For. Res. 2010, 21, 73–76. [Google Scholar] [CrossRef]
- Nouvellon, Y.; Laclau, J.P.; Epron, D.; Kinana, A.; Mabiala, A.; Roupsard, O.; Bonnefond, J.M.; Maire, G.; Marsden, C.; Bontemps, D.; et al. Within-stand and seasonal variations of specific leaf area in a clonal Eucalyptus plantation in the Republic of Congo. For. Ecol. Manag. 2010, 259, 1796–1807. [Google Scholar] [CrossRef]
- Li, L.L.; Li, M.; Zheng, R.M.; Cheng, X.P. Response of Eucalyptus Leaf Area and Biomass to Wet and Dry Season. Sci. Technol. Eng. 2016, 16, 167–170. [Google Scholar]
- Silva, W.D.; Silva, A.A.D.; Sediyama, T.; Cardoso, A.A. Biomass and leaf area of Eucalyptus citriodora and Eucalyptus grandis seedlings as affected by water content in soil and association with Brachiaria brizantha. In Pesquisa Agropecuaria Gaucha; 1999. Available online: www.fepagro.rs.gov.br/upload/1398909545_art_04.pdf (accessed on 20 November 2018).
- Aspinwall, M.J.; Drake, J.E.; Campany, C.; Varhammar, A.; Ghannoum, O.; Tissue, D.T.; Reich, P.B.; Tjoelker, M.G. Convergent acclimation of leaf photosynthesis and respiration to prevailing ambient temperatures under current and warmer climates in Eucalyptus tereticornis. New Phytol. 2016, 212, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Crous, K.Y.; Ósvaldsson, A.; Ellsworth, D.S. Is phosphorus limiting in a mature Eucalyptus woodland? Phosphorus fertilisation stimulates stem growth. Plant Soil 2015, 391, 293–305. [Google Scholar] [CrossRef]
- Wu, J.W.; He, Q.; Li, J.Y.; Wang, J.H.; Su, Y.; Wang, L.P.; Dong, J.L.; Bai, J.J. Dynamic changes of foliage growth of Catalpa bungei clones under different nitrogen exponential fertilizations. J. Beijing For. Univ. 2015, 37, 19–28. [Google Scholar]
- Qiu, Q. Interactive Effects of Light with Water and Fertilizer on the Growth and Physiological Characteristics of Catalpa bungei Clones Seedlings. Master’s Thesis, South China Agriculture University, Guangzhou, China, 2016. [Google Scholar]
- Lei, X.D.; Li, Y.C.; Xiang, W. Individual basal area growth model using multi-level linear mixed model with repeated measures. Sci. Silva. Sin. 2009, 45, 74–80. [Google Scholar]
- Sack, L.; Scoffoni, C.; Mckown, A.D.; Frole, K.; Rawls, M.; Havran, J.C.; Tran, H.; Tran, H. Developmentally based scaling of leaf venation architecture explains global ecological patterns. Nat. Commun. 2012, 3, 837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, P.M.; Bonetta, D.; Tsukaya, H.; Dengler, R.E.; Dengler, N.G. Cell Cycling and Cell Enlargement in Developing Leaves of Arabidopsis. Dev. Biol. 1999, 215, 0–419. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Ortiz-Monasterio, J.I.; Asner, G.P.; Naylor, R.L.; Falcon, W.P. Combining field surveys, remote sensing, and regression trees to understand yield variations in an irrigated wheat landscape. Agron. J. 2005, 97, 241–249. [Google Scholar]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Thiec, D.L.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides× Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Nat. Sci. Mater. Int. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Zhu, G.L.; Wei, X.Z. Leaf morphological plasticity of Ziziphus jujuba var. spinosa in response to natural drought gradient ecotopes. Acta Phytoecol. Sin. 2016, 36, 6178–6187. [Google Scholar]
- Klich, M.G. Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity. Environ. Exp. Bot. 2000, 44, 171–183. [Google Scholar] [CrossRef]
- Gates, D.M. Biophysical Ecology; Springer: New York, NY, USA, 1980; 611p. [Google Scholar]
- Yates, M.J.; Verboom, G.A.; Rebelo, A.G.; Cramer, M.D. Ecophysiological significance of leaf size variation in Proteaceae from the Cape Floristic Region. Funct. Ecol. 2010, 24, 485–492. [Google Scholar] [CrossRef]
- Martorell, C.; Ezcurra, E. The narrow-leaf syndrome: A functional and evolutionary approach to the form of fog-harvesting rosette plants. Oecologia 2007, 151, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Q.; Feng, M.; Li, Z.J. Investigation of bud burst, shoot growth and leaf expansion in Populus euphratica of different ages. Acta Ecol. Sin. 2015, 35, 1198–1207. [Google Scholar]
- Sadras, V.O.; Villalobos, F.J.; Fereres, E.; Wolfe, D.W. Leaf responses to soil water deficits: Comparative sensitivity of leaf expansion rate and leaf conductance in field-grown sunflower (Helianthus annuus, L.). Plant Soil 1993, 153, 189–194. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of, variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2010, 182, 565–588. [Google Scholar] [CrossRef]
- Tardieu, F. Modelling leaf expansion in a fluctuating environment: Are changes in specific leaf area a consequence of changes in expansion rate? New Phytol. 2010, 143, 33–43. [Google Scholar] [CrossRef]
- Zhu, X.B.; Liu, Y.M.; Sun, S.C. Leaf expansion of the dominant woody species of three deciduous oak forests in Nanjing, east China. Acta Phytoecol. Sin. 2005, 29, 128–136. [Google Scholar]
- Goldstein, G.; Bucci, S.J.; Scholz, F.G. Why do trees adjust water relations and hydraulic architecture in response to nutrient availability? Tree Physiol. 2013, 33, 238–240. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, K.W.; Abrams, M.D.; Schultz, J.C. The impact of water and nutrient deficiencies on the growth, gas exchange and water relations of red oak and chestnut oak. Tree Physiol. 1992, 11, 271. [Google Scholar] [CrossRef]
- Shen, A.; Liu, C. Effects of different application rate of urea on the growth of rice and N fertilizer utilization ratio under water leakage and non-leakage conditions. Chin. J. Riceence 1997, 11, 231–237. [Google Scholar]
- Mcdonald, P.G.; Fonseca, C.R.; Westoby, M. Leaf-Size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades? Funct. Ecol. 2010, 17, 50–57. [Google Scholar] [CrossRef]
- Santini, B.A.; Hodgson, J.G.; Thompson, K.; Wilson, P.J.; Band, S.R.; Jones, G.; Charles, M.; Bogaard, A.; Palmer, C.; Rees, M. The triangular seed mass-leaf area relationship holds for annual plants and is determined by habitat productivity. Funct. Ecol. 2017, 31, 1770–1779. [Google Scholar] [CrossRef] [Green Version]
- Milla, R.; Reich, P.B. Multi-trait interactions, not phylogeny, fine-tune leaf size reduction with increasing altitude. Ann. Bot. 2011, 107, 455. [Google Scholar] [CrossRef] [PubMed]
- Drewry, D.T.; Kumar, P.; Long, S.; Bernacchi, C.; Liang, X.Z.; Sivapalan, M. Ecohydrological responses of dense canopies to environmental variability: 1. interplay between vertical structure and photosynthetic pathway. J. Geophys. Res. Biogeosci. 2015, 115, 985–989. [Google Scholar] [CrossRef]
- Drewry, D.T.; Kumar, P.; Long, S.; Bernacchi, C.; Liang, X.Z.; Sivapalan, M. Ecohydrological responses of dense canopies to environmental variability: 2. Role of acclimation under elevated CO2. J. Geophys. Res. Biogeosci. 2015, 115, 6291–6297. [Google Scholar] [CrossRef]
- Srinivasan, V.; Kumar, P.; Long, S.P. Decreasing, not increasing, leaf area will raise crop yields under global atmospheric change. Glob. Chang. Biol. 2017, 23, 1626–1635. [Google Scholar] [CrossRef]
- Gholz, H.L. Environmental limits on aboveground net primary production, leaf area, and biomass in vegetation zones of the Pacific Northwest. Ecology 1982, 63, 469–481. [Google Scholar] [CrossRef]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, A.; Pretzsch, H.; Ruiz-peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Fertilizer (Eucalyptus Fertilizer) | Irrigation Time(s) | Irrigation Time |
|---|---|---|---|
| CK | - | - | - |
| W | - | 8 h/week | October to March of the following year |
| F | base fertilizer and top dressing | - | - |
| WF | base fertilizer and top dressing | 8 h/week | October to March of the following year |
| Treatment | Number | Height/cm | Clear Bole Height/cm | Crown Length/cm | Crown Width/m | Ground Diameter/mm | Diameter at Breast Height (DBH)/mm | Branching Quantity |
|---|---|---|---|---|---|---|---|---|
| CK | 70 | 150.36 ± 8.38 b | 21.77 ± 1.08 b | 128.59 ± 8.50 b | 1.18 × 1.16 b | 22.72 ± 1.36 b | 14.73 ± 1.33 b | 31.56 ± 3.07 b |
| W | 84 | 168.37 ± 6.87 b | 26.65 ± 1.10 a | 141.71 ± 6.83 b | 1.23 × 1.24 b | 24.25 ± 1.15 b | 15.29 ± 0.90 b | 35.00 ± 2.53 b |
| F | 66 | 454.12 ± 10.78 a | 21.17 ± 1.10 b | 432.95 ± 10.73 a | 2.17 × 2.13 a | 58.10 ± 1.54 a | 41.10 ± 1.30 a | 67.08 ± 3.58 a |
| WF | 71 | 438.80 ± 8.67 a | 22.87 ± 1.15 b | 415.93 ± 9.04 a | 2.16 × 2.12 a | 55.72 ± 1.45 a | 39.38 ± 1.31 a | 64.92 ± 2.45 a |
| Index | W | LA | LP | R | Leaf Shape Index (L/W) |
|---|---|---|---|---|---|
| Leaf length (L) | 0.644 ** | 0.880 ** | 0.988 ** | 0.749 ** | 0.370 * |
| Leaf width (W) | 0.916 ** | 0.751 ** | 0.918 ** | −0.452 ** | |
| Leaf area (LA) | 0.940 ** | 0.933 ** | −0.086 | ||
| Leaf perimeter (LP) | 0.831 ** | 0.23 | |||
| Leaf expansion rate (R) | −0.259 |
| Source of Difference | Height/cm | Clear Bole Height/cm | Crown Length/cm | SN Crown Width/m | EW Crown Width/m | Ground Diameter/mm | Diameter at Breast Height (DBH)/mm | Branching Quantity |
|---|---|---|---|---|---|---|---|---|
| Water | 0.013 | 8.548 ** | 0.233 | 0.007 | 0.251 | 0.179 | 0.105 | 0.046 |
| ertilizer | 1133.549 ** | 3.699 | 1127.112 ** | 566.200 ** | 560.166 ** | 614.645 ** | 196.417 ** | 121.021 ** |
| Water × Fertilizer | 2.787 | 1.889 | 2.186 | 0.204 | 0.434 | 1.721 | 0.399 | 0.886 |
| Source of Difference | Leaf Length (L) | Leaf Width (W) | Leaf Area (LA) | Leaf Perimeter (LP) | Leaf Expansion Rate (R) | Leaf Shape Index (L/W) | Leaf Length × Width (LW) |
|---|---|---|---|---|---|---|---|
| Water | 2.971 | 1.551 | 2.780 | 2.934 | 3.006 | 0.013 | 2.843 |
| Fertilizer | 15.438 ** | 0.556 | 1.920 | 9.639 ** | 0.016 | 45.287 ** | 2.181 |
| Water × Fertilizer | 5.124 * | 3.746 | 5.494 * | 5.420 * | 6.409 * | 0.144 | 5.338 * |
| Treatment | Regressive Equation | R2 (COD) | F | Vmax/(cm2 · Day−1) | t1/Day | t2/Day | t3/Day | Peak Days/Day |
|---|---|---|---|---|---|---|---|---|
| CK | Y = 18.064/(1 + 101.652 × exp(−0.199*x)) | 0.996 | 1694.95 ** | 0.90 | 16.61 | 23.22 | 29.84 | 13.24 |
| W | Y = 16.706/(1 + 110.307 × exp(−0.201*x)) | 0.996 | 1389.47 ** | 0.84 | 16.85 | 23.40 | 29.95 | 13.10 |
| F | Y = 17.608/(1 + 32.718 × exp(−0.139*x)) | 0.994 | 1166.41 ** | 0.61 | 15.62 | 25.09 | 34.57 | 18.95 |
| WF | Y = 23.604/(1 + 28.321 × exp(−0.167*x)) | 0.996 | 1976.4 ** | 0.99 | 12.14 | 20.02 | 27.91 | 15.77 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Truong, T.V.; He, Q.; Hua, L.; Su, Y.; Li, J. Dry Season Irrigation Promotes Leaf Growth in Eucalyptus urophylla × E. grandis under Fertilization. Forests 2019, 10, 67. https://doi.org/10.3390/f10010067
Yu F, Truong TV, He Q, Hua L, Su Y, Li J. Dry Season Irrigation Promotes Leaf Growth in Eucalyptus urophylla × E. grandis under Fertilization. Forests. 2019; 10(1):67. https://doi.org/10.3390/f10010067
Chicago/Turabian StyleYu, Fei, Thuy Van Truong, Qian He, Lei Hua, Yan Su, and Jiyue Li. 2019. "Dry Season Irrigation Promotes Leaf Growth in Eucalyptus urophylla × E. grandis under Fertilization" Forests 10, no. 1: 67. https://doi.org/10.3390/f10010067