Effects of Fertilization Ratios and Frequencies on the Growth and Nutrient Uptake of Magnolia wufengensis (Magnoliaceae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Field
2.2. Materials
2.3. Experimental Design
2.3.1. Effects of Different NPK Ratios on the Seedling Growth of M. wufengensis
2.3.2. Effects of Fertilization Frequencies on M. wufengensis Seedling Growth
2.4. Methods to Measure Physiological Parameters
2.4.1. Height and Root Collar Diameter of Seedlings
2.4.2. Determination of Chlorophyll Ccontent
2.4.3. Determination of Biomass
2.4.4. Determination of N, P, and K
2.5. Data Processing and Analysis
2.5.1. Calculation of Nutrient Parameters
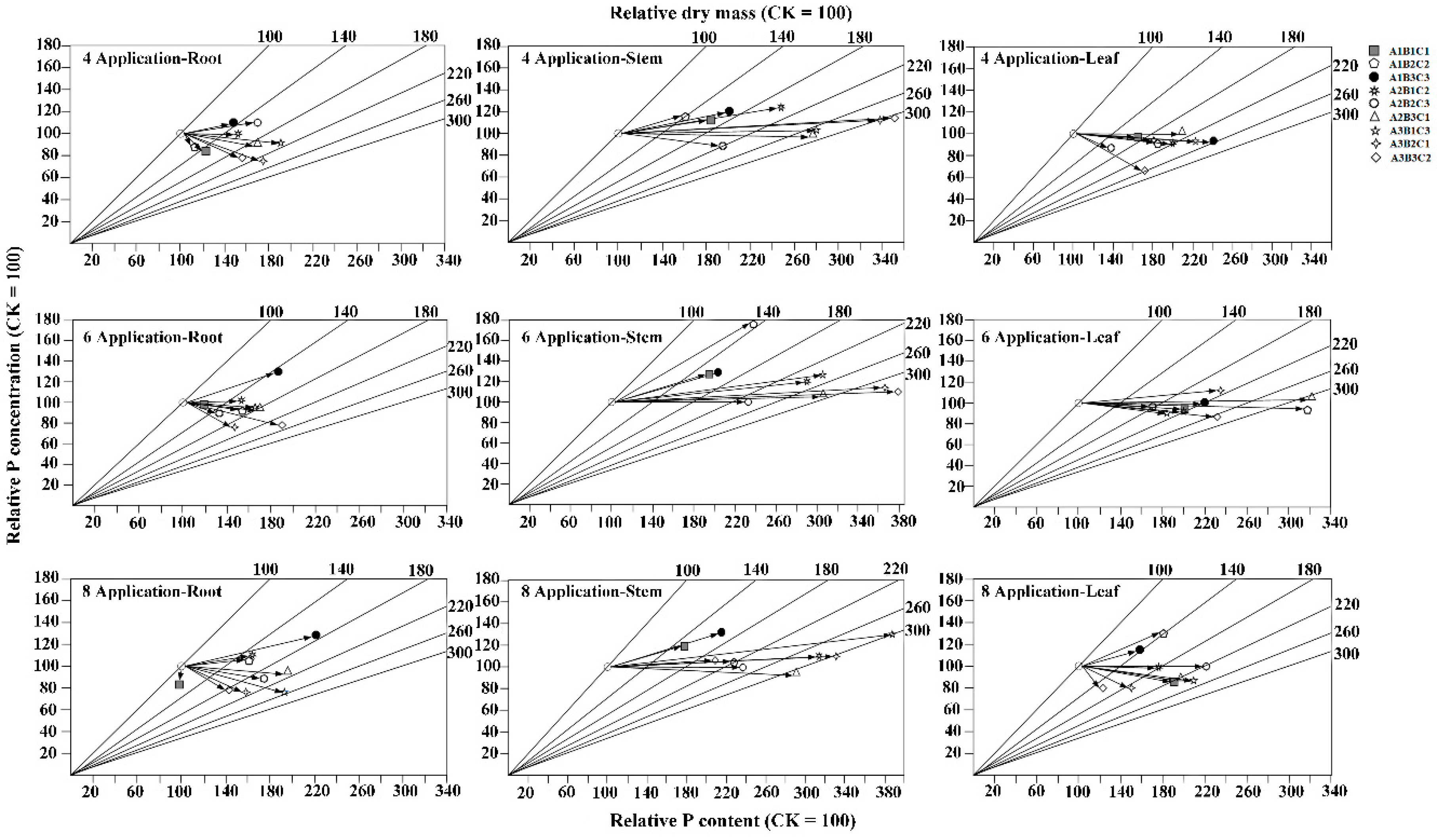
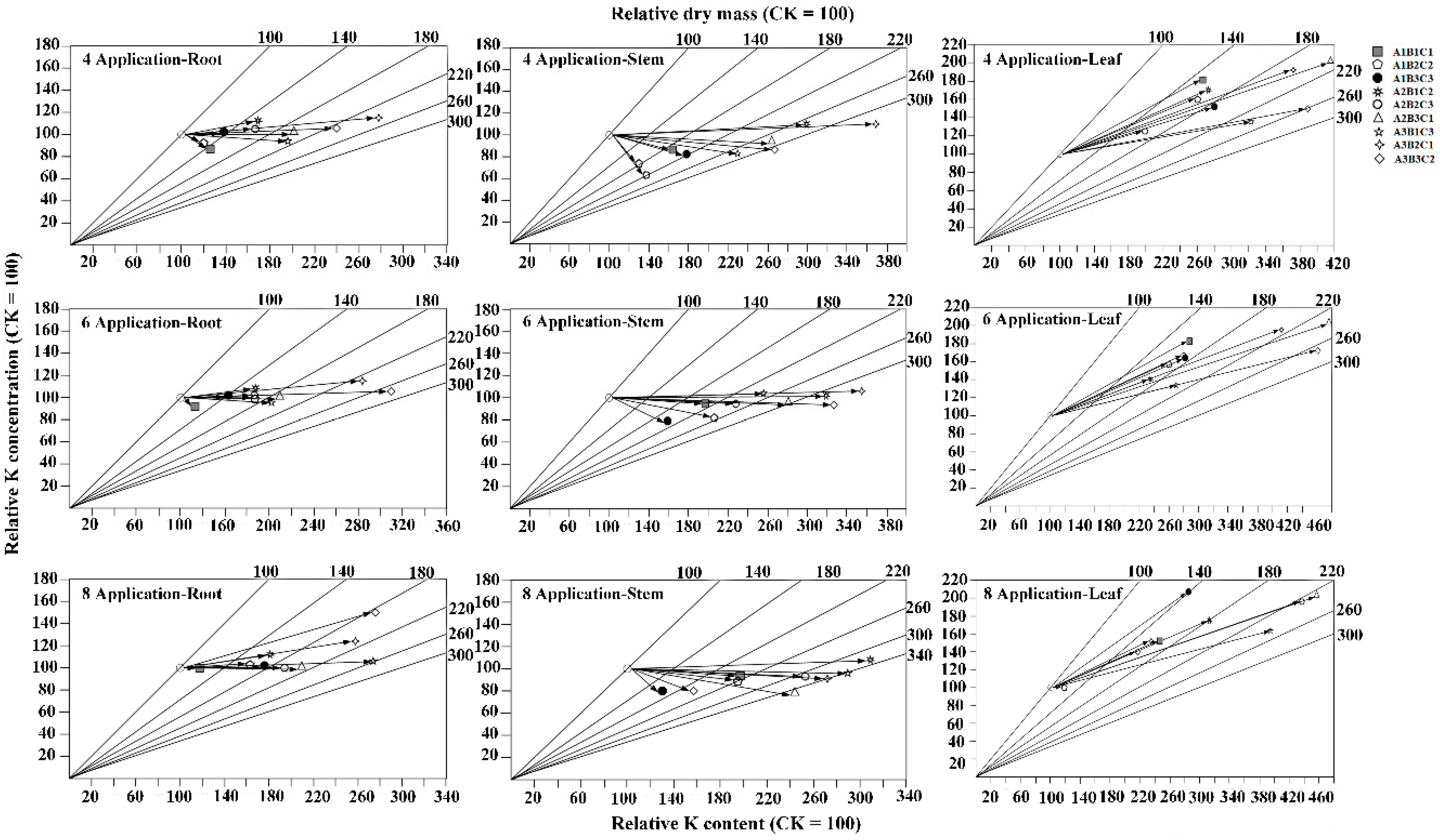
2.5.2. Vector Analysis
2.5.3. Seedling Quality Index (QI)
2.5.4. Data analysis
3. Results
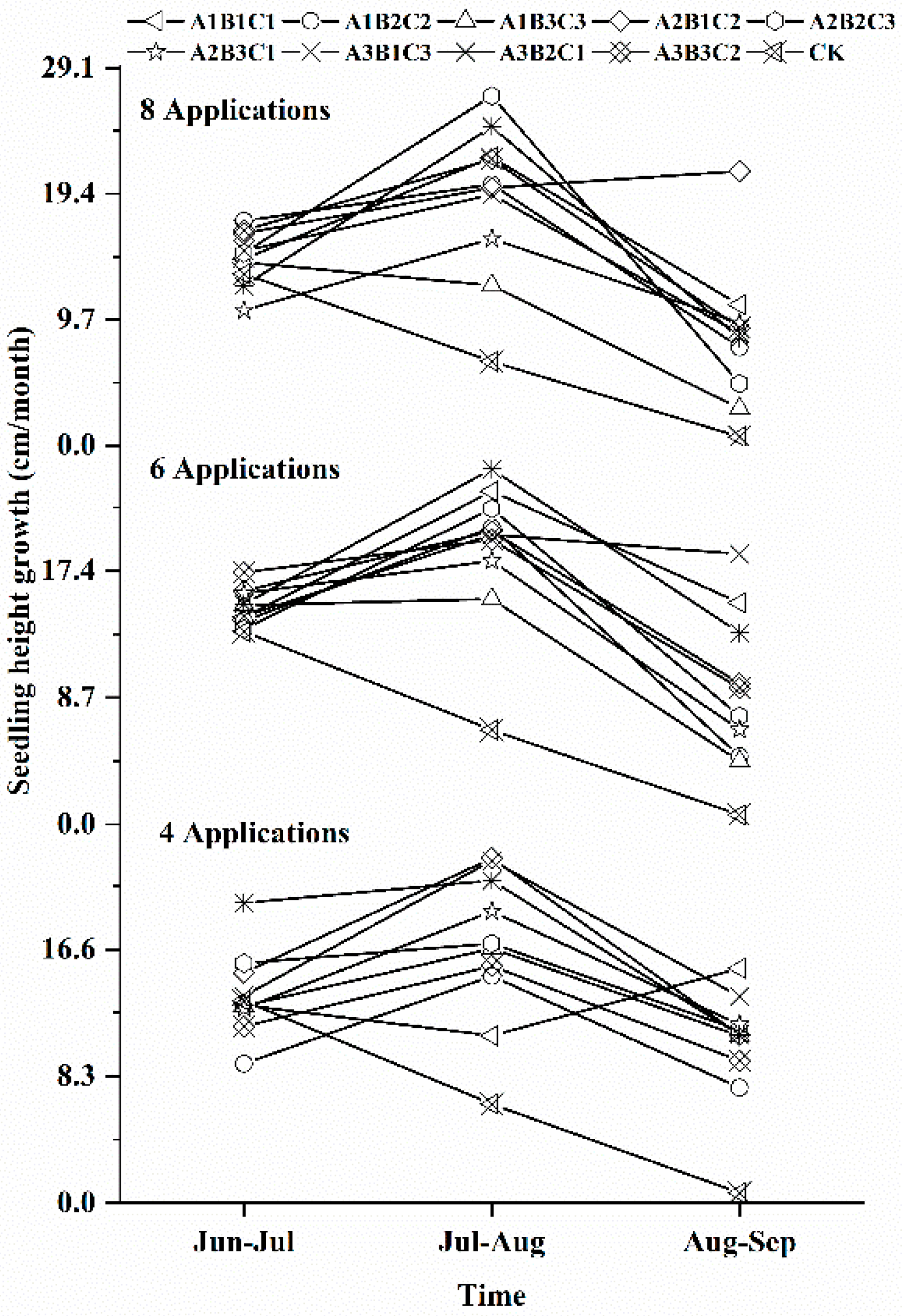
3.1. Effects of Different Fertilization Ratios and Frequencies on Seedling Height Growth
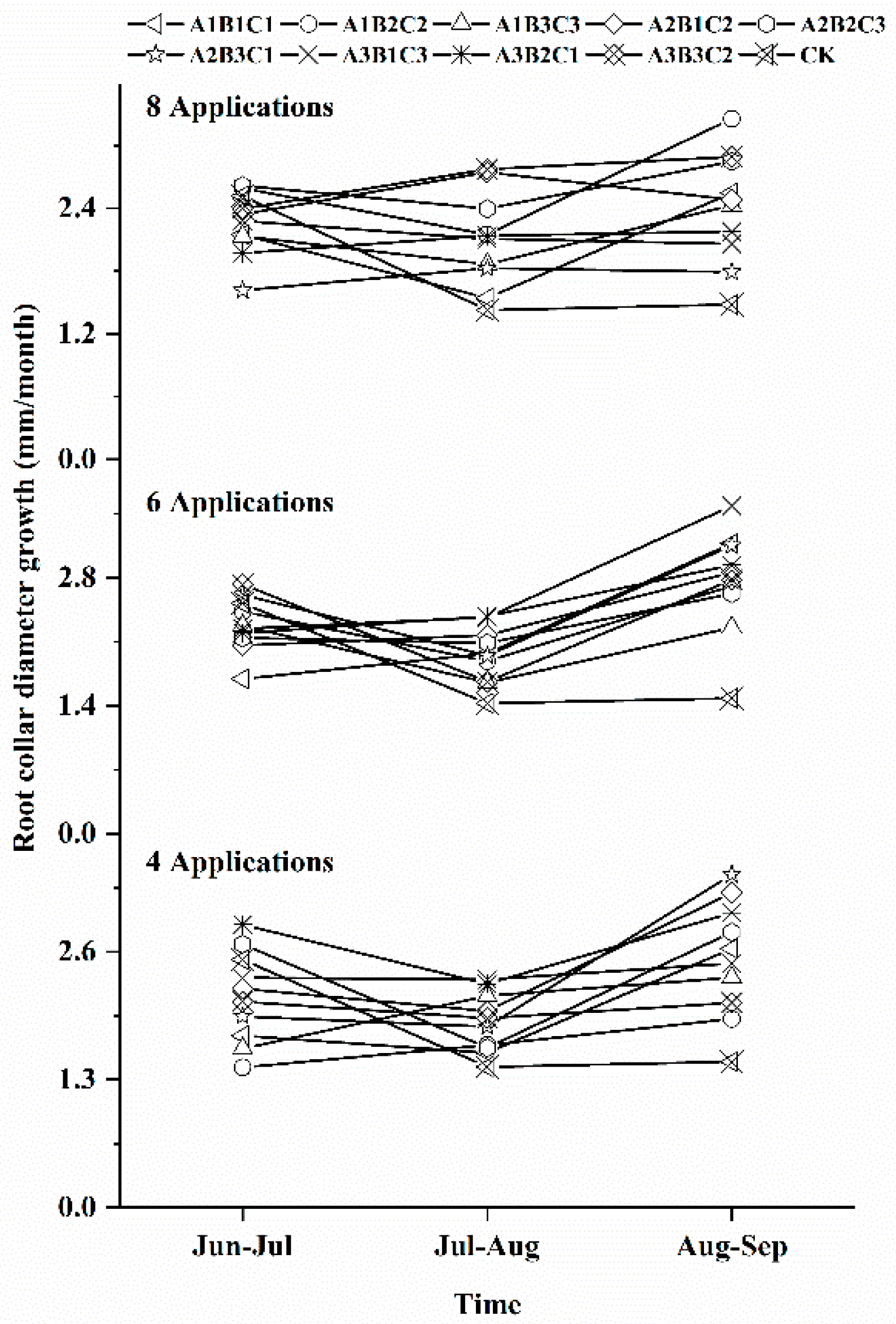
3.2. Effects of Different Fertilization Ratios and Frequencies on Seedling Root Collar Diameter
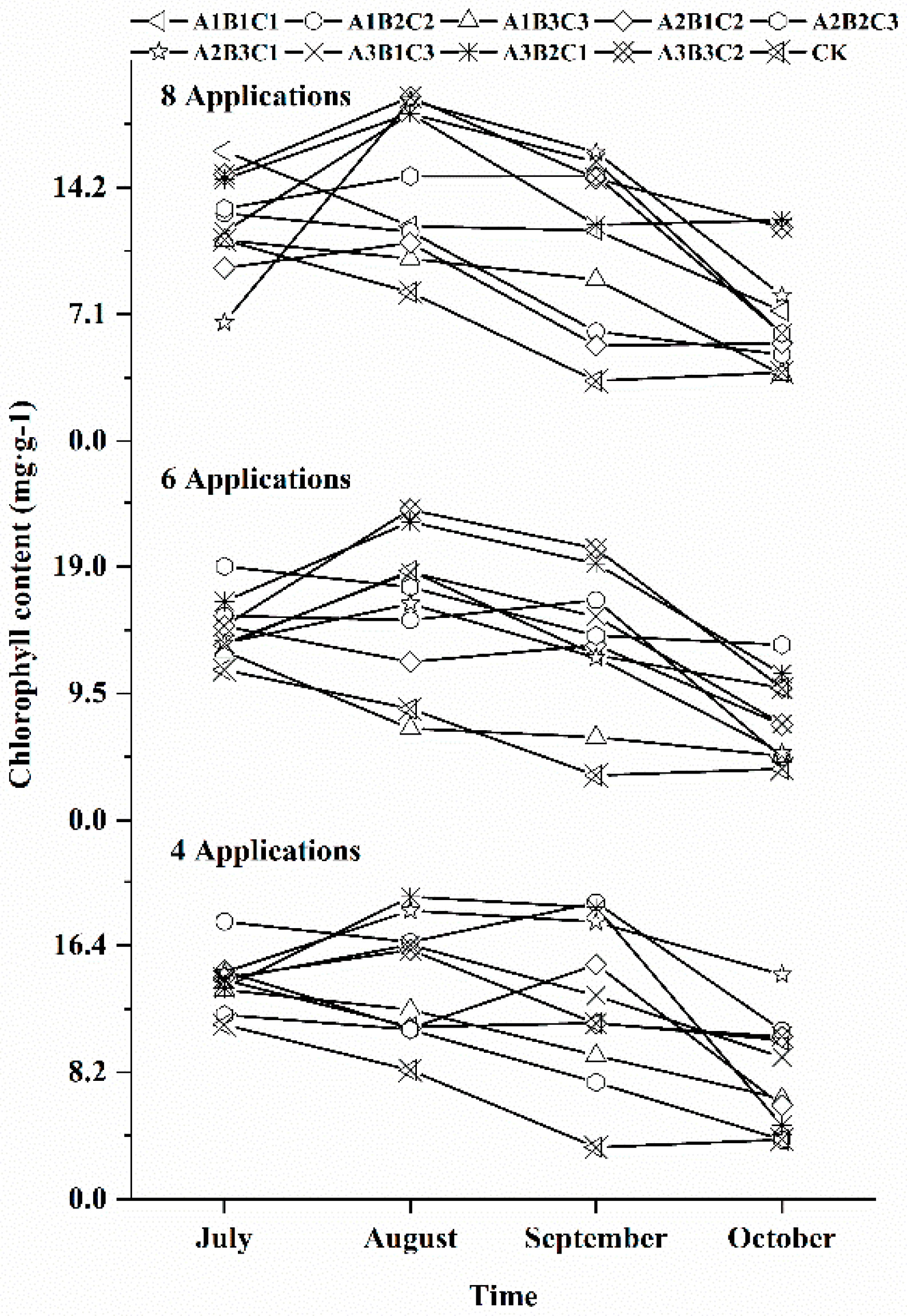
3.3. Effects of Different Fertilization Ratios and Frequencies on the Chlorophyll Content in Seedling Leaves
3.4. Effects of Different Fertilization Ratios and Frequencies on Seedling Bbiomass
3.4.1. Effects on Seedling Root Biomass
3.4.2. Effects on Seedling Stem Biomass
3.4.3. Effects on Seedling Leaf Biomass
3.4.4. Effects on Whole Seedling Biomass
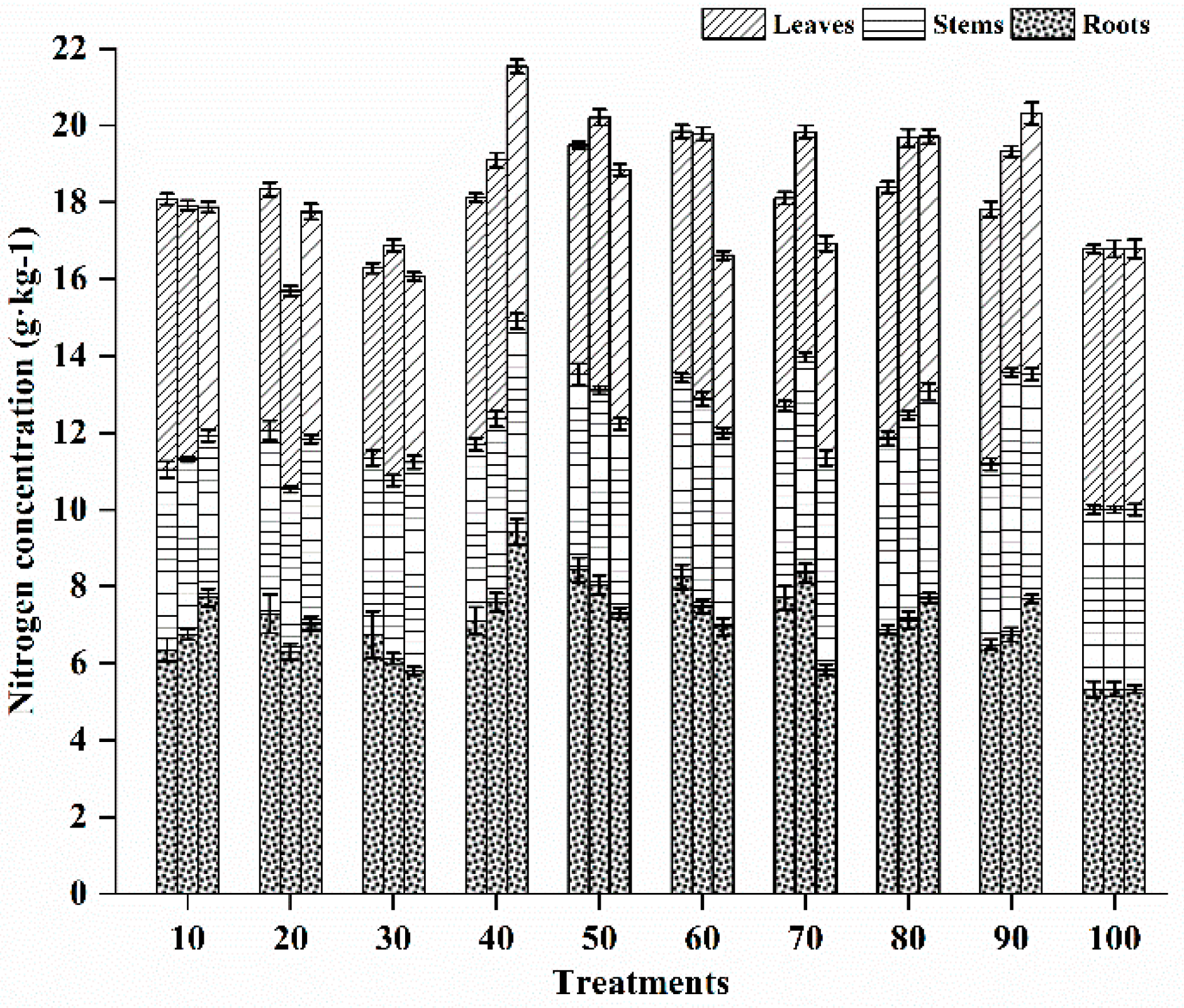
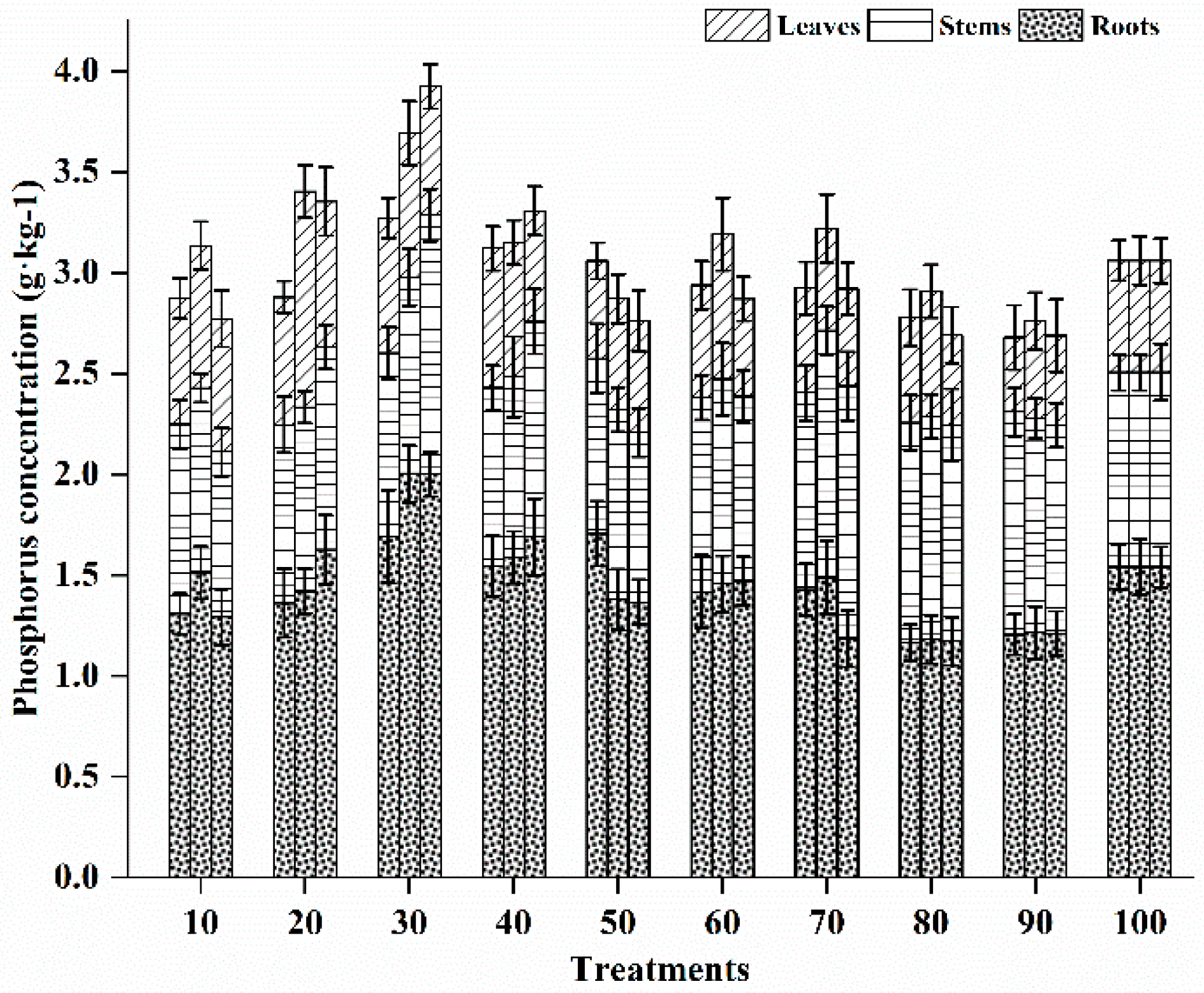
3.5. Effects of Different Fertilization Ratios and Frequencies on Nutrient Satus in Seedling Tissues
3.5.1. Nitrogen Status in Different Vegetative Tissues
3.5.2. Phosphorus Status in Different Vegetative Tissues
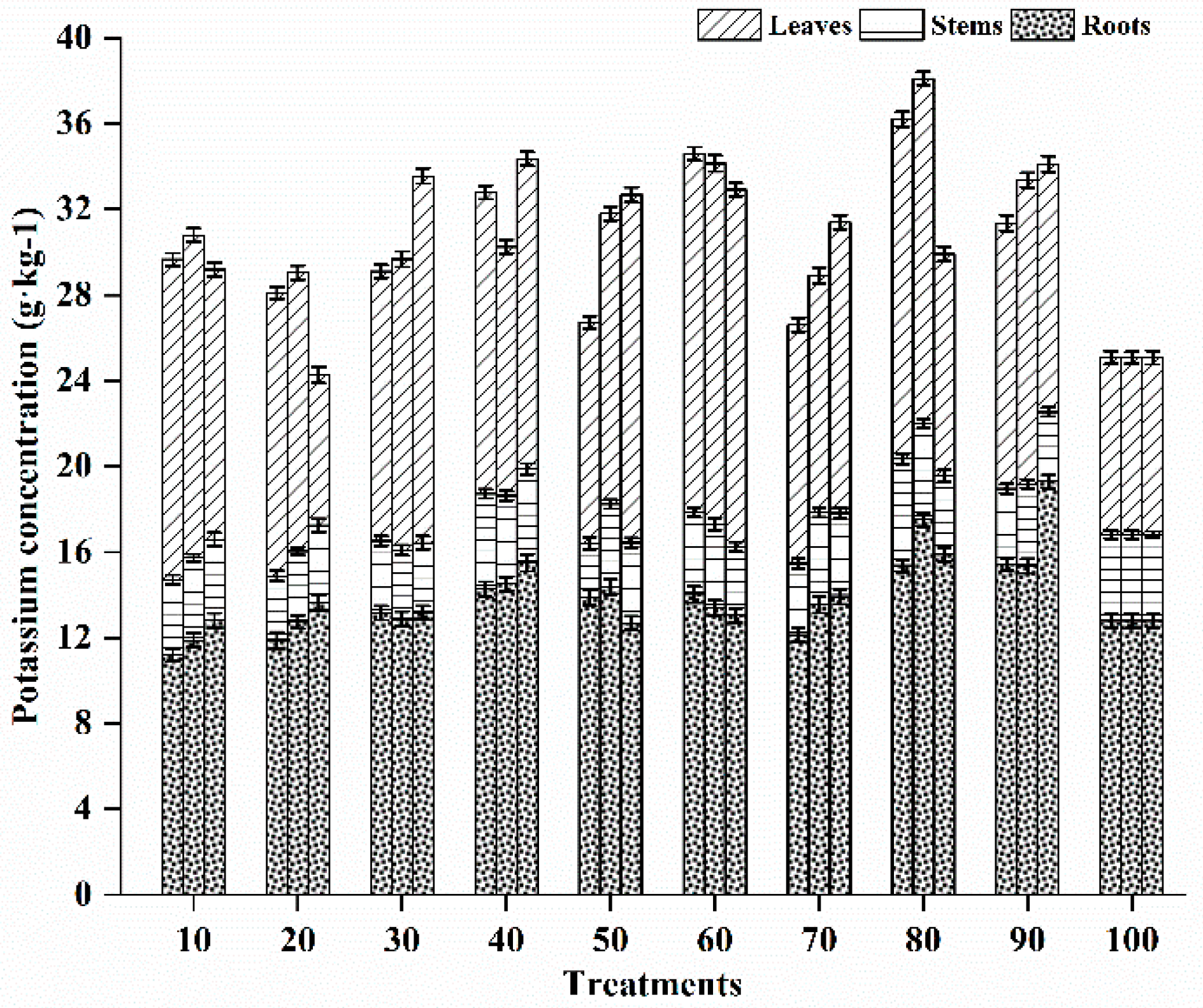
3.5.3. Potassium Status in Different Vegetative Tissues
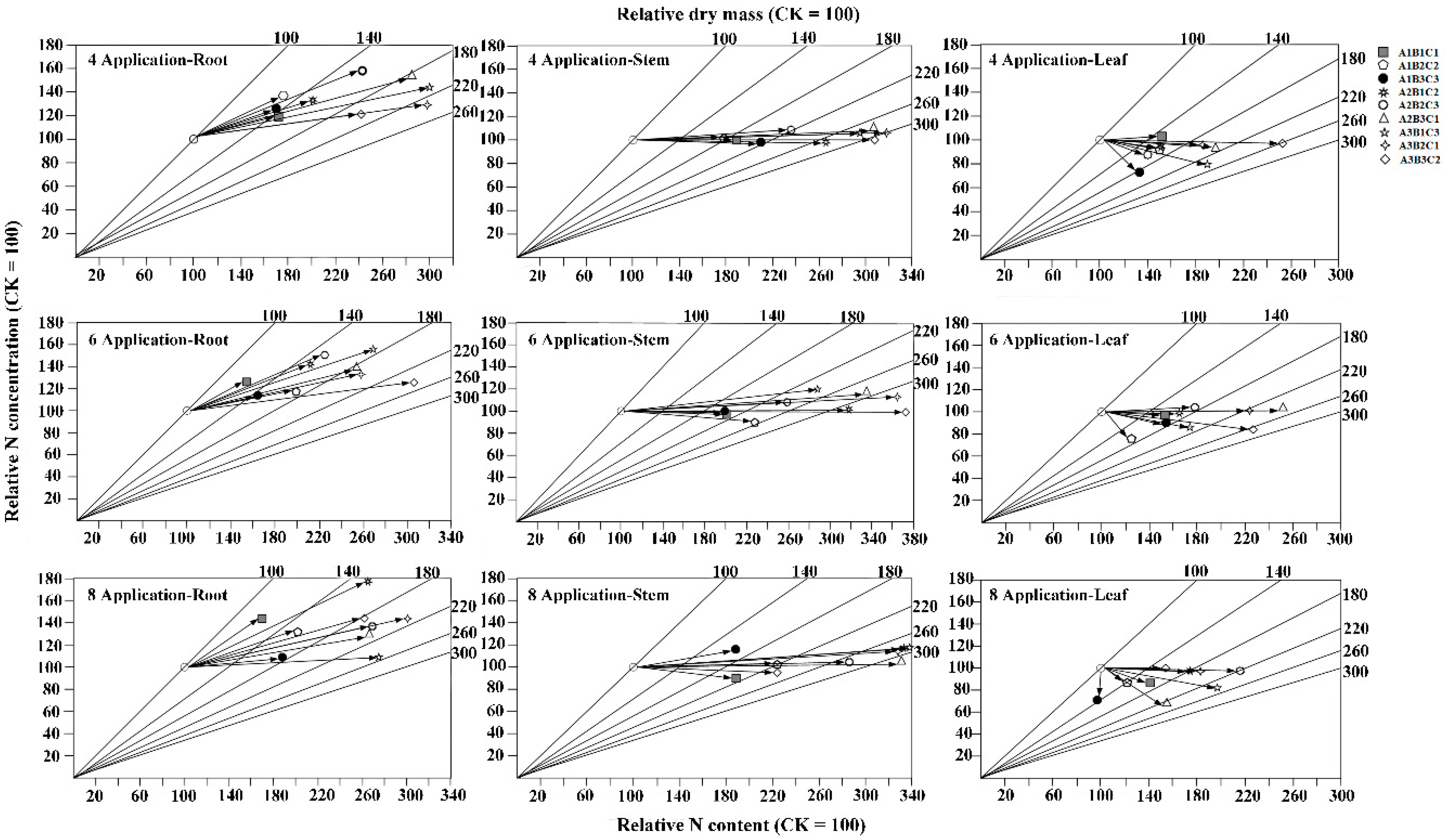
3.6. Effects of Different Fertilization Ratios and Frequencies on Nutrient Transport Efficiency
3.7. Effects of Different Fertilization Ratios and Frequencies on Nutrient Uptake Efficiency
3.8. Effects of Different Fertilization Ratios and Frequencies on Soil Nutrient Contents
3.9. Comprehensive Analysis
4. Discussion
4.1. Fertilization Promotes Plant Growth
4.2. Effects of Fertilization on the Chlorophyll Content in Magnolia wufengensis
4.3. Effects of Fertilization on Nutrient Status in Magnolia wufengensis
4.4. Effects of Fertilization on Nutrient Uptake and Transport Efficiency and Soil Nutrient Contents
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Principal Components | Score Formula |
|---|---|
| 1 | Y = 0.229X1 + 0.226X2 + 0.273X3 + 0.311X4 + 0.300X5 + 0.320X6 + 0.183X7 − 0.264X8 + 0.175X9 + 0.183X10 − 0.264X11 + 0.175X12 + 0.104X13 − 0.231X14 − 0.150X15 − 0.060X16 − 0.097X17 − 0.145X18 − 0.019X19 + 0.282X20 + 0.252X21 |
| 2 | Y = − 0.017X1 + 0.086X2 + 0.233X3 + 0.134X4 + 0.196X5 + 0.215X6 − 0.326X7 + 0.256X8 + 0.147X9 − 0.325X10 + 0.256X11 + 0.146X12 − 0.432X13 − 0.329X14 − 0.174X15 − 0.153X16 + 0.144X17 − 0.033X18 + 0.056X19 + 0.166X20 + 0.221X21 |
| 3 | Y = 0.039X1 − 0.086X2 − 0.138X3 + 0.019X4 − 0.073X5 − 0.084X6 + 0.301X7 + 0.153X8 + 0.386X9 + 0.301X10 + 0.153X11 + 0.386X12 − 0.240X13 + 0.099X14 − 0.350X15 − 0.382X16 + 0.262X17 − 0.067X18 + 0.133X19 − 0.027X20 − 0.077X21 |
| 4 | Y = − 0.276X1 − 0.109X2 + 0.158X3 − 0.105X4 − 0.105X5 + 0.026X6 − 0.036X7 − 0.093X8 + 0.309X9 − 0.036X10 − 0.093X11 + 0.310X12 − 0.004X13 − 0.028X14 + 0.226X15 + 0.353X16 − 0.474X17 + 0.375X18 + 0.330X19 + 0.038X20 + 0.036X21 |
| 5 | Y = − 0.137X1 + 0.192X2 + 0.097X3 + 0.018X4 − 0.042X5 + 0.049X6 + 0.263X7 + 0.292X8 − 0.195X9 + 0.264X10 + 0.293X11 − 0.195X12 + 0.148X13 + 0.264X14 − 0.352X15 + 0.113X16 − 0.019X17 + 0.049X18 + 0.500X19 + 0.217X20 + 0.130X21 |
| 6 | Y = − 0.530X1 − 0.189X2 + 0.150X3 + 0.315X4 − 0.083X5 + 0.171X6 − 0.031X7 + 0.047X8 + 0.017X9 − 0.031X10 + 0.047X11 + 0.017X12 − 0.027X13 + 0.027X14 − 0.178X15 + 0.029X16 + 0.130X17 + 0.499X18 + 0.132X19 − 0.135X20 − 0.428X21 |
| Frequencies | Treatments | Numbering | F1 | Ranking | F2 | Ranking | F3 | Ranking | F4 | Ranking | F5 | Ranking | F6 | Ranking | F | Ranking |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 Applications | A1B1C1 | 1 | 35.1 | 25 | 9.3 | 24 | 15.7 | 15 | −0.2 | 7 | 3.9 | 13 | 31.4 | 25 | 95.2 | 25 |
| A1B2C2 | 2 | 32.7 | 26 | 8.6 | 27 | 15.1 | 21 | 1.0 | 4 | 5.1 | 3 | 27.2 | 26 | 89.8 | 27 | |
| A1B3C3 | 3 | 36.1 | 23 | 11.3 | 15 | 14.6 | 23 | −1.5 | 11 | 3.8 | 14 | 32.3 | 22 | 96.5 | 23 | |
| A2B1C2 | 4 | 42.8 | 14 | 11.2 | 16 | 17.4 | 6 | −5.3 | 26 | 1.2 | 27 | 43.2 | 4 | 110.6 | 13 | |
| A2B2C3 | 5 | 36.4 | 21 | 9.4 | 23 | 14.7 | 22 | −1.3 | 10 | 5.8 | 1 | 31.5 | 24 | 96.5 | 24 | |
| A2B3C1 | 6 | 43.7 | 10 | 13.1 | 10 | 17.7 | 3 | −0.2 | 5 | 4.1 | 11 | 36.0 | 17 | 114.3 | 9 | |
| A3B1C3 | 7 | 45.0 | 8 | 13.5 | 9 | 13.3 | 26 | −4.9 | 25 | 4.5 | 7 | 40.6 | 10 | 112.0 | 11 | |
| A3B2C1 | 8 | 45.2 | 7 | 15.2 | 4 | 16.7 | 9 | 1.4 | 2 | 3.8 | 15 | 38.5 | 13 | 120.8 | 4 | |
| A3B3C2 | 9 | 47.2 | 4 | 14.4 | 5 | 15.2 | 20 | −4.7 | 24 | 2.6 | 21 | 42.9 | 5 | 117.7 | 6 | |
| 6 Applications | A1B1C1 | 10 | 37.5 | 19 | 9.1 | 25 | 17.2 | 8 | −3.5 | 15 | 1.6 | 26 | 38.4 | 14 | 100.3 | 19 |
| A1B2C2 | 11 | 37.8 | 18 | 12.8 | 11 | 13.8 | 25 | −2.1 | 13 | 3.7 | 16 | 34.6 | 19 | 100.5 | 18 | |
| A1B3C3 | 12 | 35.4 | 24 | 11.1 | 17 | 15.3 | 18 | −0.9 | 8 | 4.1 | 12 | 32.3 | 21 | 97.4 | 20 | |
| A2B1C2 | 13 | 43.2 | 12 | 10.9 | 19 | 16.5 | 10 | −6.1 | 27 | 2.4 | 23 | 43.2 | 3 | 110.2 | 14 | |
| A2B2C3 | 14 | 40.9 | 16 | 9.7 | 21 | 17.5 | 4 | −3.9 | 19 | 2.8 | 19 | 38.7 | 12 | 105.7 | 17 | |
| A2B3C1 | 15 | 44.7 | 9 | 13.8 | 7 | 17.3 | 7 | −1.0 | 9 | 4.5 | 6 | 36.0 | 16 | 115.5 | 7 | |
| A3B1C3 | 16 | 42.4 | 15 | 11.3 | 14 | 15.7 | 16 | −4.1 | 21 | 4.4 | 9 | 38.8 | 11 | 108.5 | 15 | |
| A3B2C1 | 17 | 47.4 | 3 | 14.3 | 6 | 18.8 | 2 | −1.8 | 12 | 2.5 | 22 | 41.0 | 7 | 122.3 | 3 | |
| A3B3C2 | 18 | 51.1 | 1 | 15.9 | 2 | 15.9 | 13 | −4.4 | 22 | 2.9 | 17 | 46.2 | 1 | 127.6 | 1 | |
| 8 Applications | A1B1C1 | 19 | 37.1 | 20 | 8.7 | 26 | 16.1 | 12 | −3.8 | 18 | 2.3 | 24 | 36.4 | 15 | 96.7 | 22 |
| A1B2C2 | 20 | 36.2 | 22 | 9.6 | 22 | 12.9 | 27 | −4.1 | 20 | 5.4 | 2 | 34.2 | 20 | 94.2 | 26 | |
| A1B3C3 | 21 | 32.6 | 27 | 12.8 | 13 | 16.2 | 11 | 4.6 | 1 | 4.8 | 4 | 25.8 | 27 | 96.7 | 21 | |
| A2B1C2 | 22 | 43.6 | 11 | 10.2 | 20 | 19.3 | 1 | −4.4 | 23 | 2.2 | 25 | 42.8 | 6 | 113.7 | 10 | |
| A2B2C3 | 23 | 43.1 | 13 | 13.5 | 8 | 15.9 | 14 | −0.2 | 6 | 4.5 | 8 | 34.8 | 18 | 111.7 | 12 | |
| A2B3C1 | 24 | 46.3 | 5 | 15.3 | 3 | 15.5 | 17 | −2.8 | 14 | 2.7 | 20 | 41.0 | 8 | 118.1 | 5 | |
| A3B1C3 | 25 | 48.5 | 2 | 16.1 | 1 | 14.4 | 24 | −3.6 | 17 | 2.9 | 18 | 45.0 | 2 | 123.3 | 2 | |
| A3B2C1 | 26 | 45.3 | 6 | 12.8 | 12 | 15.3 | 19 | −3.6 | 16 | 4.2 | 10 | 40.6 | 9 | 114.6 | 8 | |
| A3B3C2 | 27 | 39.5 | 17 | 11.1 | 18 | 17.5 | 5 | 1.4 | 3 | 4.8 | 5 | 31.9 | 23 | 106.1 | 16 |
References
- Ma, L.; Luorong, W.; He, S.; Liu, X.; Wang, X. A new species of magnolia (magnoliaceae) from Hubei, China. Bull. Bot. Res. 2006, 26, 4–7. [Google Scholar]
- Sang, Z. Diversity Analysis of the Floral Traits and Study on the Drought Resistance of magnolia wufengensis. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2011. [Google Scholar]
- Ma, L.; Luorong, W.; He, S.; Liu, X.; Wang, X. A new variety of magnolia (magnoliaceae) from Hubei, China. Bull. Bot. Res. 2006, 26, 516–519. [Google Scholar]
- Pennazio, S. Mineral nutrition of plants: A short history of plant physiology. Riv. Biol. 2005, 98, 215–236. [Google Scholar] [PubMed]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: New York, NY, USA, 2011; Volume 24. [Google Scholar]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. In Advances in Ecological Research; Fitter, A.H., Raffaelli, D.G., Eds.; Academic Press: New York, NY, USA, 1999; Volume 30, pp. 1–67. [Google Scholar]
- Luo, J.; Qin, J.; He, F.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Net fluxes of ammonium and nitrate in association with h+ fluxes in fine roots of populus popularis. Planta 2013, 237, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, H.; Zhao, H.; Korpelainen, H.; Li, C. Sexually different physiological responses of Populus cathayana to nitrogen and phosphorus deficiencies. Tree Physiol. 2014, 34, 343–354. [Google Scholar] [CrossRef]
- Marschner, H. Mineral nutrition of higher plants. J. Ecol. 1995, 76, 1250. [Google Scholar]
- Gan, H.; Jiao, Y.; Jia, J.; Wang, X.; Li, H.; Shi, W.; Peng, C.; Polle, A.; Luo, Z.B. Phosphorus and nitrogen physiology of two contrasting poplar genotypes when exposed to phosphorus and/or nitrogen starvation. Tree Physiol. 2016, 36, 22–38. [Google Scholar] [CrossRef]
- Burman, U.; Garg, B.K.; Kathju, S. Effect of phosphorus application on clusterbean under different intensities of water stress. J. Plant Nutr. 2009, 32, 668–680. [Google Scholar] [CrossRef]
- Dos Santos, M.G.; Ribeiro, R.V.; de Oliveira, R.F.; Machado, E.C.; Pimentel, C. The role of inorganic phosphate on photosynthesis recovery of common bean after a mild water deficit. Plant Sci. 2006, 170, 659–664. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Christina, M.; Le Maire, G.; Battie-Laclau, P.; Nouvellon, Y.; Bouillet, J.P.; Jourdan, C.; de Moraes Goncalves, J.L.; Laclau, J.P. Measured and modeled interactive effects of potassium deficiency and water deficit on gross primary productivity and light-use efficiency in Eucalyptus grandis plantations. Glob. Chang. Biol. 2015, 21, 2022–2039. [Google Scholar] [CrossRef] [PubMed]
- Armengaud, P.; Breitling, R.; Amtmann, A. The potassium-dependent transcriptome of arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef] [PubMed]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Principles of Plant Nutrition; International Potash Institute: Basel, Switzerland, 1982; pp. 639–655. [Google Scholar]
- Glass, A.D.M. Nitrogen use efficiency of crop plants: Physiological constraints upon nitrogen absorption. Crit. Rev. Plant Sci. 2003, 22, 453–470. [Google Scholar] [CrossRef]
- Raun, W.R.; Solie, J.B.; Johnson, G.V.; Stone, M.L.; Mullen, R.W.; Freeman, K.W.; Thomason, W.E.; Lukina, E.V. Improving nitrogen use efficiency in cereal grain production with optical sensing and variable rate application. Agron. J. 2002, 94, 815–820. [Google Scholar] [CrossRef]
- Bussi, C.; Amiot, M.J. Effects of nitrogen and potassium fertilization on the growth, yield and pitburn of apricot (cv. Bergeron). J. Hortic. Sci. Biotechnol. 1998, 73, 387–392. [Google Scholar] [CrossRef]
- Bussi, C.; Besset, J.; Girard, T. Effects of fertilizer rates and dates of application on apricot (cv bergeron) cropping and pitburn. Sci. Hortic. 2003, 98, 139–147. [Google Scholar] [CrossRef]
- Chu, H.; Lin, X.; Fujii, T.; Morimoto, S.; Yagi, K.; Hu, J.; Zhang, J. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem. 2007, 39, 2971–2976. [Google Scholar] [CrossRef]
- Kanchikerimath, M.; Singh, D. Soil organic matter and biological properties after 26 years of maize–wheat–cowpea cropping as affected by manure and fertilization in a cambisol in semiarid region of india. Agric. Ecosyst. Environ. 2001, 86, 155–162. [Google Scholar] [CrossRef]
- Cai, Z.C.; Qin, S.W. Dynamics of crop yields and soil organic carbon in a long-term fertilization experiment in the huang-huai-hai plain of china. Geoderma 2006, 136, 708–715. [Google Scholar] [CrossRef]
- Osmond, D.L.; Riha, S.J. Nitrogen fertilizer requirements for maize produced in the tropics: A comparison of three computer-based recommendation systems. Agric. Syst. 1996, 50, 37–50. [Google Scholar] [CrossRef]
- Konde, N.M.; Kanase, N.M.; Jadhao, S.M.; Goud, V.V. Yield targetting equation for soybean with conjoint use of manure and chemical fertilizer based on fertility gradient approach. Ann. Plant Physiol. 2009, 23, 210–214. [Google Scholar]
- Abbasi, M.K.; Tahir, M.M.; Rahim, N. Effect of n fertilizer source and timing on yield and n use efficiency of rainfed maize (Zea mays L.) in kashmir–pakistan. Geoderma 2013, 195–196, 87–93. [Google Scholar] [CrossRef]
- Van Eekeren, N.; de Boer, H.; Hanegraaf, M.; Bokhorst, J.; Nierop, D.; Bloem, J.; Schouten, T.; de Goede, R.; Brussaard, L. Ecosystem services in grassland associated with biotic and abiotic soil parameters. Soil Biol. Biochem. 2010, 42, 1491–1504. [Google Scholar] [CrossRef]
- De Boer, H.C.; Deru, J.G.C.; Hoekstra, N.J.; van Eekeren, N. Strategic timing of nitrogen fertilization to increase root biomass and nitrogen-use efficiency of Lolium perenne L. Plant Soil 2016, 407, 81–90. [Google Scholar] [CrossRef]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef]
- Ruiz Diaz, D.A.; Sawyer, J.E. Plant-available nitrogen from poultry manure as affected by time of application. Agron. J. 2008, 100, 1318. [Google Scholar] [CrossRef]
- Randall, G.W.; Vetsch, J.A. Corn production on a subsurface-drained mollisol as affected by fall versus spring application of nitrogen and nitrapyrin. Agron. J. 2005, 97, 472–478. [Google Scholar] [CrossRef]
- Rozas, H.R.S.; Echeverría, H.E.; Barbieri, P.A. Nitrogen balance as affected by application time and nitrogen fertilizer rate in irrigated no-tillage maize. Agron. J. 2004, 96, 1622. [Google Scholar] [CrossRef]
- Rozas, H.S.; Echeverría, H.; Andrade, F.; Studdert, G. Effect of urease inhibitor and fertilization time on nitrogen uptake and maize grain yield under no-tillage. Rev. Fac. Agron. La Plata 1997, 102, 129–136. [Google Scholar]
- Garrido-Lestache, E.; López-Bellido, R.J.; López-Bellido, L. Durum wheat quality under mediterranean conditions as affected by n rate, timing and splitting, n form and s fertilization. Eur. J. Agron. 2005, 23, 265–278. [Google Scholar] [CrossRef]
- Zebarth, B.; Leclerc, Y.; Moreau, G. Rate and timing of nitrogen fertilization of russet burbank potato: Nitrogen use efficiency. Can. J. Plant Sci. 2004, 84, 845–854. [Google Scholar] [CrossRef]
- Yang, L.; Pan, C.; Shao, S.; Tao, C.; Wang, W.; Ying, Y. Effects of pp_(333) and drought stress on the activity, photosynthetic characteristics, and non-structural carbohydrates of phyllostachys edulis seedlings. Acta Ecol. Sin. 2018, 39, 46–59. [Google Scholar]
- Haase, D.L.; Rose, R. Vector analysis and its use for interpreting plant nutrient shifts in response to silvicultural trea. For. Sci. 1995, 41, 54–66. [Google Scholar]
- Imo, M.; Timmer, V.R. Vector diagnosis of nutrient dynamics in mesquite seedlings. For. Sci. 1997, 43, 268–273. [Google Scholar]
- Chapin, F.S.; Vitousek, P.M.; Cleve, K.V. The nature of nutrient limitation in plant communities. Am. Nat. 1986, 127, 48–58. [Google Scholar] [CrossRef]
- Li, J.; Guo, Q.; Zhang, J.; Korpelainen, H.; Li, C. Effects of nitrogen and phosphorus supply on growth and physiological traits of two Larix species. Environ. Exp. Bot. 2016, 130, 206–215. [Google Scholar] [CrossRef]
- Rothstein, D.E.; Zak, D.R.; Pregitzer, K.S.; Curtis, P.S. Kinetics of nitrogen uptake by Populus tremuloides in relation to atmospheric co2 and soil nitrogen availability. Tree Physiol. 2000, 20, 265–270. [Google Scholar] [CrossRef]
- Ibrahim, L.; Proe, M.F.; Cameron, A.D. Interactive effects of nitrogen and water availabilities on gas exchange and whole-plant carbon allocation in poplar. Tree Physiol. 1998, 18, 481–487. [Google Scholar] [CrossRef]
- Gordon, W.S.; Jackson, R.B. Nutrient concentrations in fine roots. Ecology 2000, 81, 275–280. [Google Scholar] [CrossRef]
- Wang, J.R.; Hawkins, C.D.; Letchford, T. Relative growth rate and biomass allocation of paper birch (Betula pap). Can. J. For. Res. 1998, 28, 44–55. [Google Scholar] [CrossRef]
- Proe, M.F.; Millard, P. Effect of p supply upon seasonal growth and internal cycling of p in sitka spruce (Picea sitchensis (bong.)carr.) seedlings. Plant Soil 1995, 168/169, 313–317. [Google Scholar] [CrossRef]
- Deng, S.; Yin, Q.; Zhang, S.; Shi, K.; Jia, Z.; Ma, L. Drip irrigation affects the morphology and distribution of olive roots. HortScience 2017, 52, 1298–1306. [Google Scholar] [CrossRef]
- Klemens, J.A. Biomass allocation and growth of tropical dry forest tree seedlings across light and soil gradients. Ph.D. Thesis, University of Pennsylvania, Philadelphia, PA, USA, 2003. [Google Scholar]
- van der Werf, A.; Nagel, O.W. Carbon allocation to shoots and roots in relation to nitrogen supply is mediated by cytokinins and sucrose: Opinion. Plant Soil 1996, 185, 21–32. [Google Scholar] [CrossRef]
- Bekele, A.; Hudnall, W.H.; Tiarks, A.E. Response of densely stocked loblolly pine (Pinus taeda L.) to applied nitrogen and phosphorus. South. J. Appl. For. 2003, 27, 180–189. [Google Scholar]
- Coops, N.C.; Stone, C.; Culvenor, D.S.; Chisholm, L. Assessment of crown condition in eucalypt vegetation by remotely sensed optical indices. J. Environ. Qual. 2004, 33, 956. [Google Scholar] [CrossRef]
- Curran, P.J.; Dungan, J.L.; Gholz, H.L. Exploring the relationship between reflectance red edge and chlorophyll content in slash pine. Tree Physiol. 1991, 7, 33–48. [Google Scholar] [CrossRef]
- Moran, J.A.; Mitchell, A.K.; Goodmanson, G.; Stockburger, K.A. Differentiation among effects of nitrogen fertilization treatments on conifer seedlings by foliar reflectance: A comparison of methods. Tree Physiol. 2000, 20, 1113–1120. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology; Springer-Verlag: Berlin, Germany, 1982; pp. 1217–1220. [Google Scholar]
- Duru, M. Evaluation of chlorophyll meter to assess nitrogen status of cocksfoot sward. J. Plant Nutr. 2002, 25, 275–286. [Google Scholar] [CrossRef]
- Rao, I.M.; Terry, N. Leaf phosphate status, photosynthesis, and carbon partitioning in sugar beet. Plant Physiol. 1989, 90, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Liu, G. Effects of Balanced Fertilization on Mulberry Yield and Quality and Its Fertilization Model. Ph.D. Thesis, Southwest University, Chongqing, China, 2012. [Google Scholar]
- Machler, N. Influence of inorganic-phosphate on photosynthesis of wheat chloroplasts 1 photosynthesis and assimilate export at 5-degrees-c and 25-degrees-c. Biotechnol. Bioeng. 1984, 108, 481–487. [Google Scholar]
- Wu, C.; Wang, Z.; Sun, H.; Guo, S. Effects of different concentrations of nitrogen and phosphorus on chlorophyll biosynthesis, chlorophyll a fluorescence, and photosynthesis in Larix olgensis seedlings. Front. For. China 2006, 1, 170–175. [Google Scholar] [CrossRef]
- Wu, T.G.; Wu, M.; Liu, L.; Xiao, J.H.; He, J.S.; Han, X.G. Seasonal variations of leaf nitrogen and phosphorus stoichiometry of three herbaceous species in hangzhou bay coastal wetlands, China. Chin. J. Plant Ecol. 2010, 34, 23–28. [Google Scholar]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef]
- Lovett Doust, J.; Lovett Doust, L. Plant reproductive ecology: Patterns and strategies. Evolution 1988, 93, 646–647. [Google Scholar]
- He, J.S.; Wang, L.; Flynn, D.F.; Wang, X.; Ma, W.; Fang, J. Leaf nitrogen:Phosphorus stoichiometry across chinese grassland biomes. Oecologia 2008, 155, 301–310. [Google Scholar] [CrossRef]
- Li, J. Functional Analysis of Potassium Channel Osaktl and Its Regulators in Rice k+ Uptake. Ph.D. Thesis, China Agricultural University, Guangzhou, China, 2014. [Google Scholar]
- Tester, M.; Leigh, R.A. Partitioning of nutrient transport processes in roots. J. Exp. Bot. 2001, 52, 445–457. [Google Scholar] [CrossRef]
- Güsewell, S. High nitrogen: Phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytol. 2005, 166, 537. [Google Scholar] [CrossRef]
- Ingestad, T.; Lund, A.B. Theory and techniques for steady state mineral nutrition and growth of plants. Scand. J. For. Res. 1986, 1, 439–453. [Google Scholar] [CrossRef]
- Suriyagoda, L.D.B.; Ryan, M.H.; Renton, M.; Lambers, H. Above- and below-ground interactions of grass and pasture legume species when grown together under drought and low phosphorus availability. Plant Soil 2011, 348, 281–297. [Google Scholar] [CrossRef]
- Evans, H.J.; Wildes, R.A. Potassium and its role in enzyme activation. In Proceedings of the 8th Colloquium of the International Potash Institute, Uppsala, Sweden, 14–17 June 1971. [Google Scholar]
- Wyn Jones, R.G.; Pollard, A. Proteins, enzymes and inorganic ions. Encycl. Plant Physiol. New 1983, 92, 528–562. [Google Scholar]









| BD (g·cm−3) | FC (%) | PH | OM (g·kg−1) | TN (g·kg−1) | AP (g·kg−1) | AK (g·kg−1) | Sand (%) | Silt (%) | Clay (%) | ST |
|---|---|---|---|---|---|---|---|---|---|---|
| 1.61 | 16.9 | 7.86 | 19.23 | 0.37 | 0.47 | 4.08 | 62.51 | 36.89 | 0.60 | loam |
| Treatment | N (mg·Plant−1) | P (mg·Plant−1) | K (mg·Plant−1) |
|---|---|---|---|
| A1B1C1 | 160 | 80 | 80 |
| A1B2C2 | 160 | 160 | 160 |
| A1B3C3 | 160 | 320 | 320 |
| A2B1C2 | 320 | 80 | 160 |
| A2B2C3 | 320 | 160 | 320 |
| A2B3C1 | 320 | 320 | 80 |
| A3B1C3 | 480 | 80 | 320 |
| A3B2C1 | 480 | 160 | 80 |
| A3B3C2 | 480 | 320 | 160 |
| CK | 0 | 0 | 0 |
| Source of Variation | Degree of Freedom | Height | Root Collar Diameter | Chlorophyll |
|---|---|---|---|---|
| Fertilizer ratio (F) | 9 | <0.001 | <0.05 | <0.001 |
| Application frequency (A) | 2 | 0.087 | 0.058 | <0.001 |
| Time (T) | 3 | <0.001 | <0.001 | <0.001 |
| F × A | 18 | <0.05 | <0.05 | <0.001 |
| F × T | 27 | <0.001 | <0.05 | <0.001 |
| A × T | 6 | 0.933 | 0.288 | <0.001 |
| F × A × T | 54 | 0.995 | 0.459 | <0.001 |
| Treatments | 4 Applications | 6 Applications | 8 Applications |
|---|---|---|---|
| A1B1C1 | 48.10 ± 1.7985Bb | 61.22 ± 1.4316Abc | 58.52 ± 3.0447Aab |
| A1B2C2 | 41.36 ± 1.4511Bc | 52.12 ± 0.9281Ad | 52.82 ± 3.5309Abc |
| A1B3C3 | 50.34 ± 2.7899Ab | 49.70 ± 3.6003Ad | 37.26 ± 1.6238Bd |
| A2B1C2 | 68.42 ± 1.4302Aa | 67.60 ± 2.5977Aab | 66.84 ± 3.0336Aa |
| A2B2C3 | 47.98 ± 2.6299Bb | 61.40 ± 1.0569Abc | 52.44 ± 2.9115Bbc |
| A2B3C1 | 54.74 ± 1.7299Ab | 55.52 ± 2.2739Acd | 62.60 ± 3.6212Aa |
| A3B1C3 | 62.50 ± 1.1983Aa | 61.30 ± 2.5108Abc | 66.94 ± 2.4281Aa |
| A3B2C1 | 54.70 ± 2.0594Bb | 63.10 ± 1.1261Aab | 61.62 ± 2.7838Aa |
| A3B3C2 | 66.18 ± 3.4773Aa | 69.34 ± 2.4817Aa | 48.94 ± 1.3692Bc |
| CK | 34.24 ± 2.4667Ad | 34.24 ± 2.4667Ae | 34.24 ± 2.4667Ad |
| Treatments | 4 Applications | 6 Applications | 8 Applications |
|---|---|---|---|
| A1B1C1 | 9.750 ± 0.3677Aa | 9.440 ± 0.4802Ab | 10.008 ± 0.5918Abc |
| A1B2C2 | 8.952 ± 0.6038Bab | 10.614 ± 0.2990Aab | 11.190 ± 0.6157Aab |
| A1B3C3 | 10.632 ± 0.7294Aa | 9.754 ± 0.7879Ab | 9.000 ± 0.5486Acd |
| A2B1C2 | 10.096 ± 0.6715Aa | 10.784 ± 0.4578Aab | 9.640 ± 0.4816Abc |
| A2B2C3 | 9.914 ± 0.5593Ba | 9.996 ± 0.3668Bab | 12.012 ± 0.8459Aa |
| A2B3C1 | 10.152 ± 0.7193Aa | 11.126 ± 0.5824Aab | 11.048 ± 0.6771Aab |
| A3B1C3 | 10.786 ± 0.4820Aa | 10.374 ± 0.6372Aab | 9.454 ± 0.2535Abcd |
| A3B2C1 | 10.744 ± 0.7254Aa | 11.672 ± 0.3132Aa | 10.980 ± 0.7005Aab |
| A3B3C2 | 10.518 ± 0.6523Aa | 11.138 ± 0.5604Aab | 10.768 ± 0.5180Aabc |
| CK | 7.674 ± 0.5501Ab | 7.674 ± 0.5501Ac | 7.674 ± 0.5501Ad |
| Source of Variation | Degree of Freedom | Total Biomass | Root Biomass | Stem Biomass | Leaf Biomass |
|---|---|---|---|---|---|
| Fertilizer ratio (F) | 9 | <0.001 | <0.001 | <0.001 | <0.001 |
| Application frequency (A) | 2 | 0.784 | 0.322 | 0.110 | 0.596 |
| F × A | 18 | <0.05 | 0.167 | <0.05 | 0.333 |
| Treatments | 4 Applications | 6 Applications | 8 Applications | |||
|---|---|---|---|---|---|---|
| Root Biomass (g·Plant−1) | Percentage of Total Plants | Root Biomass (g·Plant−1) | Percentage of Total Plants | Root Biomass (g·Plant−1) | Percentage of Total Plants | |
| A1B1C1 | 9.79 ± 1.06Acde | 50.2% | 8.26 ± 0.66Acd | 44.0% | 7.95 ± 0.68Acd | 42.5% |
| A1B2C2 | 8.71 ± 0.30Ade | 47.3% | 11.38 ± 1.44Abc | 48.4% | 10.37 ± 1.16Abcd | 49.9% |
| A1B3C3 | 9.12 ± 0.94Ade | 44.4% | 9.68 ± 1.27Abcd | 47.5% | 11.57 ± 1.59Abc | 57.3% |
| A2B1C2 | 10.24 ± 0.81Abcde | 44.8% | 10.07 ± 1.73Abcd | 41.7% | 10.15 ± 0.58Abcd | 43.0% |
| A2B2C3 | 10.40 ± 2.03Abcde | 48.8% | 10.09 ± 1.03Abcd | 45.9% | 13.30 ± 1.01Aab | 48.3% |
| A2B3C1 | 12.46 ± 1.78Aabcd | 47.0% | 12.23 ± 1.64Abc | 44.1% | 13.89 ± 1.48Aab | 47.3% |
| A3B1C3 | 14.02 ± 1.53ABab | 48.6% | 11.64 ± 1.51Cbc | 47.8% | 17.04 ± 1.49Aa | 52.3% |
| A3B2C1 | 15.69 ± 1.07Aa | 52.3% | 13.05 ± 1.38Aab | 45.8% | 14.10 ± 1.48Aab | 49.9% |
| A3B3C2 | 13.48 ± 1.03Aabc | 45.3% | 16.33 ± 1.28Aa | 48.0% | 12.35 ± 1.82Ab | 55.0% |
| CK | 6.77 ± 0.79Ae | 54.2% | 6.77 ± 0.79Ad | 54.2% | 6.77 ± 0.79Ad | 54.2% |
| Treatments | 4 Applications | 6 Applications | 8 Applications | |||
|---|---|---|---|---|---|---|
| Stem Biomass (g·Plant−1) | Percentage of Total Plants | Stem Biomass (g·Plant−1) | Percentage of Total Plants | Stem Biomass (g·Plant−1) | Percentage of Total Plants | |
| A1B1C1 | 5.82 ± 0.60Ab | 29.9% | 6.36 ± 0.25Ac | 33.9% | 6.50 ± 0.37Abc | 34.8% |
| A1B2C2 | 5.42 ± 0.34Ab | 29.5% | 7.79 ± 1.39Abc | 33.1% | 6.76 ± 0.51Ab | 32.5% |
| A1B3C3 | 6.62 ± 0.67Ab | 32.2% | 6.21 ± 0.88Ac | 30.5% | 5.00 ± 0.31Ac | 24.8% |
| A2B1C2 | 8.38 ± 0.49Aa | 36.7% | 9.66 ± 0.65Aab | 40.0% | 8.78 ± 0.72Aa | 37.2% |
| A2B2C3 | 6.70 ± 0.56Bb | 31.5% | 7.38 ± 0.24ABbc | 33.6% | 8.40 ± 0.52Aa | 30.5% |
| A2B3C1 | 8.58 ± 0.53Aa | 32.3% | 8.98 ± 1.28Aab | 32.4% | 9.50 ± 0.44Aa | 32.4% |
| A3B1C3 | 8.56 ± 0.50Aa | 29.6% | 7.42 ± 0.80Abc | 30.4% | 9.24 ± 0.59Aa | 28.4% |
| A3B2C1 | 9.24 ± 0.59Aa | 30.8% | 9.88 ± 0.41Aab | 34.7% | 9.24 ± 0.77Aa | 32.7% |
| A3B3C2 | 9.47 ± 0.64Aa | 31.8% | 10.64 ± 0.71Aa | 31.3% | 6.04 ± 0.61Bbc | 26.9% |
| CK | 3.09 ± 0.33Ac | 24.7% | 3.09 ± 0.33Ad | 24.7% | 3.09 ± 0.33Ad | 24.7% |
| Treatments | 4 Applications | 6 Applications | 8 Applications | |||
|---|---|---|---|---|---|---|
| Leaf Biomass (g·Plant−1) | Percentage of Total Plants | Leaf Biomass (g·Plant−1) | Percentage of Total Plants | Leaf Biomass (g·Plant−1) | Percentage of Total Plants | |
| A1B1C1 | 3.87 ± 0.42Abc | 19.9% | 4.16 ± 0.49Acd | 22.2% | 4.24 ± 0.42Abcd | 22.7% |
| A1B2C2 | 4.27 ± 0.41Abc | 23.2% | 4.35 ± 0.59Acd | 18.5% | 3.67 ± 0.56Acd | 17.6% |
| A1B3C3 | 4.82 ± 0.32Aabc | 23.4% | 4.51 ± 0.77Abcd | 22.1% | 3.61 ± 0.37Acd | 17.9% |
| A2B1C2 | 4.22 ± 0.93Abc | 18.5% | 4.39 ± 0.91Acd | 18.2% | 4.69 ± 0.63Aabc | 19.8% |
| A2B2C3 | 4.19 ± 0.38Bbc | 19.7% | 4.49 ± 0.61ABbcd | 20.5% | 5.83 ± 0.43Aab | 21.2% |
| A2B3C1 | 5.49 ± 0.41Aab | 20.7% | 6.54 ± 0.45Aab | 23.6% | 5.95 ± 0.58Aab | 20.3% |
| A3B1C3 | 6.29 ± 1.15Aab | 21.8% | 5.31 ± 0.65Aabc | 21.8% | 6.32 ± 0.82Aa | 19.4% |
| A3B2C1 | 5.08 ± 0.51Aabc | 16.9% | 5.55 ± 0.79Aabc | 19.5% | 4.93 ± 0.93Aabc | 17.4% |
| A3B3C2 | 6.81 ± 1.48Aa | 22.9% | 7.06 ± 0.50Aa | 20.8% | 4.07 ± 0.47Abcd | 18.1% |
| CK | 2.63 ± 0.60Ac | 21.1% | 2.63 ± 0.60Ad | 21.1% | 2.63 ± 0.60Ad | 21.1% |
| Treatments | 4 Applications | 6 Applications | 8 Applications |
|---|---|---|---|
| A1B1C1 | 19.48 ± 1.90Ad | 18.78 ± 0.68Acd | 18.70 ± 1.35Ad |
| A1B2C2 | 18.40 ± 0.63Ad | 23.53 ± 2.30Abc | 20.80 ± 1.73Ad |
| A1B3C3 | 20.56 ± 1.62Acd | 20.40 ± 2.46Ac | 20.18 ± 1.85Ad |
| A2B1C2 | 22.85 ± 1.71Abcd | 24.12 ± 3.08Abc | 23.63 ± 1.31Abcd |
| A2B2C3 | 21.30 ± 1.92Bcd | 21.96 ± 1.44Bbc | 27.53 ± 1.74Aabc |
| A2B3C1 | 26.52 ± 2.59Aabc | 27.74 ± 2.63Aab | 29.34 ± 1.98Aab |
| A3B1C3 | 28.87 ± 2.83Aab | 24.37 ± 2.65Abc | 32.60 ± 2.79Aa |
| A3B2C1 | 30.02 ± 2.00Aa | 28.49 ± 2.08Aab | 28.27 ± 2.89Aabc |
| A3B3C2 | 29.77 ± 2.54ABa | 34.03 ± 2.31Aa | 22.46 ± 2.85Bcd |
| CK | 12.50 ± 1.49Ae | 12.50 ± 1.49Ad | 12.50 ± 1.49Ae |
| Fertilization Frequencies | Treatments | Transport Efficiency | |||||
|---|---|---|---|---|---|---|---|
| N | Ranking | P | Ranking | K | Ranking | ||
| 4 Applications | A1B1C1 | 16.7% | 7 | 11.1% | 8 | 23.2% | 5 |
| A1B2C2 | 18.4% | 4 | 11.4% | 7 | 16.7% | 7 | |
| A1B3C3 | 5.1% | 10 | 26.6% | 1 | 20.9% | 6 | |
| A2B1C2 | 17.0% | 5 | 20.8% | 2 | 36.2% | 3 | |
| A2B2C3 | 25.8% | 2 | 18.4% | 3 | 11.0% | 8 | |
| A2B3C1 | 28.0% | 1 | 13.7% | 5 | 43.7% | 2 | |
| A3B1C3 | 16.9% | 6 | 13.1% | 6 | 10.5% | 9 | |
| A3B2C1 | 18.7% | 3 | 7.5% | 9 | 50.4% | 1 | |
| A3B3C2 | 15.0% | 8 | 3.7% | 10 | 30.2% | 4 | |
| CK | 8.4% | 9 | 18.4% | 3 | 4.1% | 10 | |
| 6 Applications | A1B1C1 | 15.7% | 7 | 21.3% | 6 | 27.9% | 5 |
| A1B2C2 | 1.3% | 10 | 31.7% | 2 | 20.6% | 8 | |
| A1B3C3 | 9.0% | 8 | 42.8% | 1 | 23.2% | 7 | |
| A2B1C2 | 23.3% | 6 | 21.9% | 5 | 25.7% | 6 | |
| A2B2C3 | 30.4% | 1 | 11.1% | 9 | 32.1% | 4 | |
| A2B3C1 | 27.7% | 3 | 23.5% | 4 | 41.8% | 2 | |
| A3B1C3 | 28.0% | 2 | 24.6% | 3 | 20.1% | 9 | |
| A3B2C1 | 27.0% | 4 | 12.5% | 8 | 58.3% | 1 | |
| A3B3C2 | 24.8% | 5 | 6.8% | 10 | 38.6% | 3 | |
| CK | 8.4% | 9 | 18.4% | 7 | 4.1% | 10 | |
| 8 Applications | A1B1C1 | 15.3% | 5 | 7.2% | 7 | 21.2% | 8 |
| A1B2C2 | 14.6% | 6 | 29.8% | 2 | 0.8% | 10 | |
| A1B3C3 | 3.7% | 10 | 51.8% | 1 | 39.4% | 3 | |
| A2B1C2 | 39.0% | 1 | 27.9% | 3 | 42.7% | 1 | |
| A2B2C3 | 21.6% | 4 | 6.9% | 8 | 35.7% | 5 | |
| A2B3C1 | 7.3% | 9 | 11.1% | 6 | 36.7% | 4 | |
| A3B1C3 | 9.3% | 7 | 13.0% | 5 | 30.4% | 6 | |
| A3B2C1 | 27.2% | 3 | 4.1% | 9 | 24.3% | 7 | |
| A3B3C2 | 31.1% | 2 | 4.0% | 10 | 41.7% | 2 | |
| CK | 8.4% | 8 | 18.4% | 4 | 4.1% | 9 | |
| Fertilization Frequencies | Treatments | Nutrient Uptake Efficiency | |||||
|---|---|---|---|---|---|---|---|
| N | Ranking | P | Ranking | K | Ranking | ||
| 4 Applications | A1B1C1 | 57.9% | 4 | −23.6% | 9 | 57.3% | 2 |
| A1B2C2 | 73.9% | 2 | −22.6% | 8 | 37.7% | 5 | |
| A1B3C3 | −54.4% | 9 | 26.5% | 1 | 50.3% | 3 | |
| A2B1C2 | 30.2% | 5 | 3.8% | 2 | 48.2% | 4 | |
| A2B2C3 | 72.9% | 3 | −0.1% | 3 | 10.2% | 8 | |
| A2B3C1 | 83.7% | 1 | −7.6% | 5 | 59.5% | 1 | |
| A3B1C3 | 19.8% | 7 | −4.3% | 4 | 4.8% | 9 | |
| A3B2C1 | 25.7% | 6 | −8.8% | 6 | 34.8% | 6 | |
| A3B3C2 | 13.8% | 8 | −11.9% | 7 | 19.6% | 7 | |
| 6 Applications | A1B1C1 | 47.6% | 6 | 9.3% | 3 | 71.5% | 1 |
| A1B2C2 | −91.9% | 9 | 42.8% | 2 | 49.6% | 4 | |
| A1B3C3 | −17.3% | 8 | 78.9% | 1 | 57.5% | 2 | |
| A2B1C2 | 60.6% | 3 | 5.7% | 5 | 32.4% | 7 | |
| A2B2C3 | 95.4% | 1 | −11.8% | 9 | 42.0% | 5 | |
| A2B3C1 | 82.0% | 2 | 8.2% | 4 | 56.7% | 3 | |
| A3B1C3 | 55.7% | 4 | 5.0% | 6 | 12.0% | 9 | |
| A3B2C1 | 52.5% | 5 | −4.8% | 7 | 40.7% | 6 | |
| A3B3C2 | 45.2% | 7 | −9.4% | 8 | 25.9% | 8 | |
| 8 Applications | A1B1C1 | 44.6% | 3 | −36.0% | 9 | 51.4% | 3 |
| A1B2C2 | 37.1% | 6 | 36.7% | 2 | −10.1% | 9 | |
| A1B3C3 | −68.2% | 9 | 49.2% | 1 | 55.0% | 2 | |
| A2B1C2 | 136.9% | 1 | 15.4% | 3 | 58.0% | 1 | |
| A2B2C3 | 52.7% | 4 | −18.6% | 8 | 47.5% | 5 | |
| A2B3C1 | −16.8% | 7 | −11.9% | 7 | 49.0% | 4 | |
| A3B1C3 | −4.6% | 8 | −4.3% | 4 | 19.7% | 7 | |
| A3B2C1 | 53.1% | 5 | −11.6% | 5 | 15.1% | 8 | |
| A3B3C2 | 65.7% | 2 | −11.6% | 5 | 28.2% | 6 | |
| Fertilization Frequencies | Treatments | Soil Nutrient Contents | ||
|---|---|---|---|---|
| N | P | K | ||
| 4 Applications | A1B1C1 | 2.76 ± 0.257ab | 1.45 ± 0.019a | 5.06 ± 0.101ab |
| A1B2C2 | 2.56 ± 0.015ab | 1.50 ± 0.015a | 5.01 ± 0.299ab | |
| A1B3C3 | 2.93 ± 0.188ab | 1.59 ± 0.034a | 4.83 ± 0.133ab | |
| A2B1C2 | 2.56 ± 0.107ab | 1.59 ± 0.009a | 4.49 ± 0.278ab | |
| A2B2C3 | 2.64 ± 0.053ab | 1.36 ± 0.266a | 4.72 ± 0.328ab | |
| A2B3C1 | 2.52 ± 0.097b | 0.66 ± 0.011b | 4.30 ± 0.089b | |
| A3B1C3 | 2.65 ± 0.008ab | 0.83 ± 0.019b | 4.30 ± 0.123b | |
| A3B2C1 | 2.96 ± 0.163a | 0.84 ± 0.017b | 6.22 ± 0.160a | |
| A3B3C2 | 2.68 ± 0.057ab | 0.92 ± 0.018b | 4.07 ± 0.176b | |
| CK | 2.62 ± 0.051ab | 0.89 ± 0.074b | 4.95 ± 0.112ab | |
| 6 Applications | A1B1C1 | 2.34 ± 0.027b | 1.00 ± 0.192b | 5.14 ± 0.112a |
| A1B2C2 | 4.28 ± 0.577a | 1.49±0.009a | 4.66 ± 0.276ab | |
| A1B3C3 | 2.84 ± 0.048b | 1.48 ± 0.018a | 5.07 ± 0.05a | |
| A2B1C2 | 2.52 ± 0.055b | 1.58 ± 0.007a | 4.72 ± 0.116ab | |
| A2B2C3 | 2.71 ± 0.054b | 1.61 ± 0.027a | 4.22 ± 0.067bc | |
| A2B3C1 | 2.50 ± 0.148b | 0.73± 0.025c | 3.98 ± 0.175c | |
| A3B1C3 | 2.82 ± 0.036b | 0.77 ± 0.028c | 4.40 ± 0.177bc | |
| A3B2C1 | 2.63 ± 0.086b | 0.86 ± 0.014bc | 4.07 ± 0.269c | |
| A3B3C2 | 2.89 ± 0.082b | 0.87 ± 0.010bc | 4.70 ± 0.056ab | |
| CK | 2.62 ± 0.051b | 0.89 ± 0.074bc | 4.95 ± 0.112a | |
| 8 Applications | A1B1C1 | 2.60 ± 0.082b | 1.49 ± 0.042b | 4.70 ± 0.154a |
| A1B2C2 | 4.53 ± 0.157a | 1.49 ± 0.002b | 5.03 ± 0.063a | |
| A1B3C3 | 2.37 ± 0.022b | 1.57 ± 0.045ab | 4.87 ± 0.220a | |
| A2B1C2 | 2.74 ± 0.050b | 1.61 ± 0.025a | 4.96 ± 0.198a | |
| A2B2C3 | 2.77 ± 0.145b | 0.75 ± 0.025de | 4.66 ± 0.116a | |
| A2B3C1 | 2.62 ± 0.081b | 0.72 ± 0.024e | 4.25 ± 0.283a | |
| A3B1C3 | 2.82 ± 0.100b | 0.82 ± 0.012cde | 4.96 ± 0.707a | |
| A3B2C1 | 2.56 ± 0.043b | 0.84 ± 0.020cd | 4.40 ± 0.178a | |
| A3B3C2 | 2.84 ± 0.119b | 0.92 ± 0.043c | 4.70 ± 0.143a | |
| CK | 2.62 ± 0.051b | 0.89 ± 0.074c | 4.95 ± 0.112a | |
| Treatments | 4 Applications | 6 Applications | 8 Applications | |||
|---|---|---|---|---|---|---|
| QI | Ranking | QI | Ranking | QI | Ranking | |
| A1B1C1 | 2.12 | 8 | 1.84 | 9 | 2.12 | 8 |
| A1B2C2 | 2.35 | 7 | 2.41 | 5 | 2.15 | 7 |
| A1B3C3 | 2.69 | 5 | 2.37 | 6 | 2.23 | 6 |
| A2B1C2 | 1.93 | 9 | 2.14 | 7 | 2.04 | 9 |
| A2B2C3 | 2.35 | 6 | 2.11 | 8 | 3.43 | 1 |
| A2B3C1 | 2.78 | 4 | 3.42 | 1 | 2.80 | 2 |
| A3B1C3 | 3.01 | 1 | 2.45 | 4 | 2.51 | 4 |
| A3B2C1 | 2.79 | 3 | 2.92 | 3 | 2.60 | 3 |
| A3B3C2 | 2.84 | 2 | 3.19 | 2 | 2.37 | 5 |
| CK | 1.59 | 10 | 1.59 | 10 | 1.59 | 10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, S.; Shi, K.; Ma, J.; Zhang, L.; Ma, L.; Jia, Z. Effects of Fertilization Ratios and Frequencies on the Growth and Nutrient Uptake of Magnolia wufengensis (Magnoliaceae). Forests 2019, 10, 65. https://doi.org/10.3390/f10010065
Deng S, Shi K, Ma J, Zhang L, Ma L, Jia Z. Effects of Fertilization Ratios and Frequencies on the Growth and Nutrient Uptake of Magnolia wufengensis (Magnoliaceae). Forests. 2019; 10(1):65. https://doi.org/10.3390/f10010065
Chicago/Turabian StyleDeng, Shixin, Kankan Shi, Jiang Ma, Lili Zhang, Luyi Ma, and Zhongkui Jia. 2019. "Effects of Fertilization Ratios and Frequencies on the Growth and Nutrient Uptake of Magnolia wufengensis (Magnoliaceae)" Forests 10, no. 1: 65. https://doi.org/10.3390/f10010065