

Travelling through the Natural Hierarchies of Type I Collagen with X-rays: From Tendons of Cattle, Horses, Sheep and Pigs

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Wide- and Small-Angle X-ray Measurements
2.3. Mechanical Behavior
2.4. Thermal Properties
2.5. Statistical Analysis
3. Results
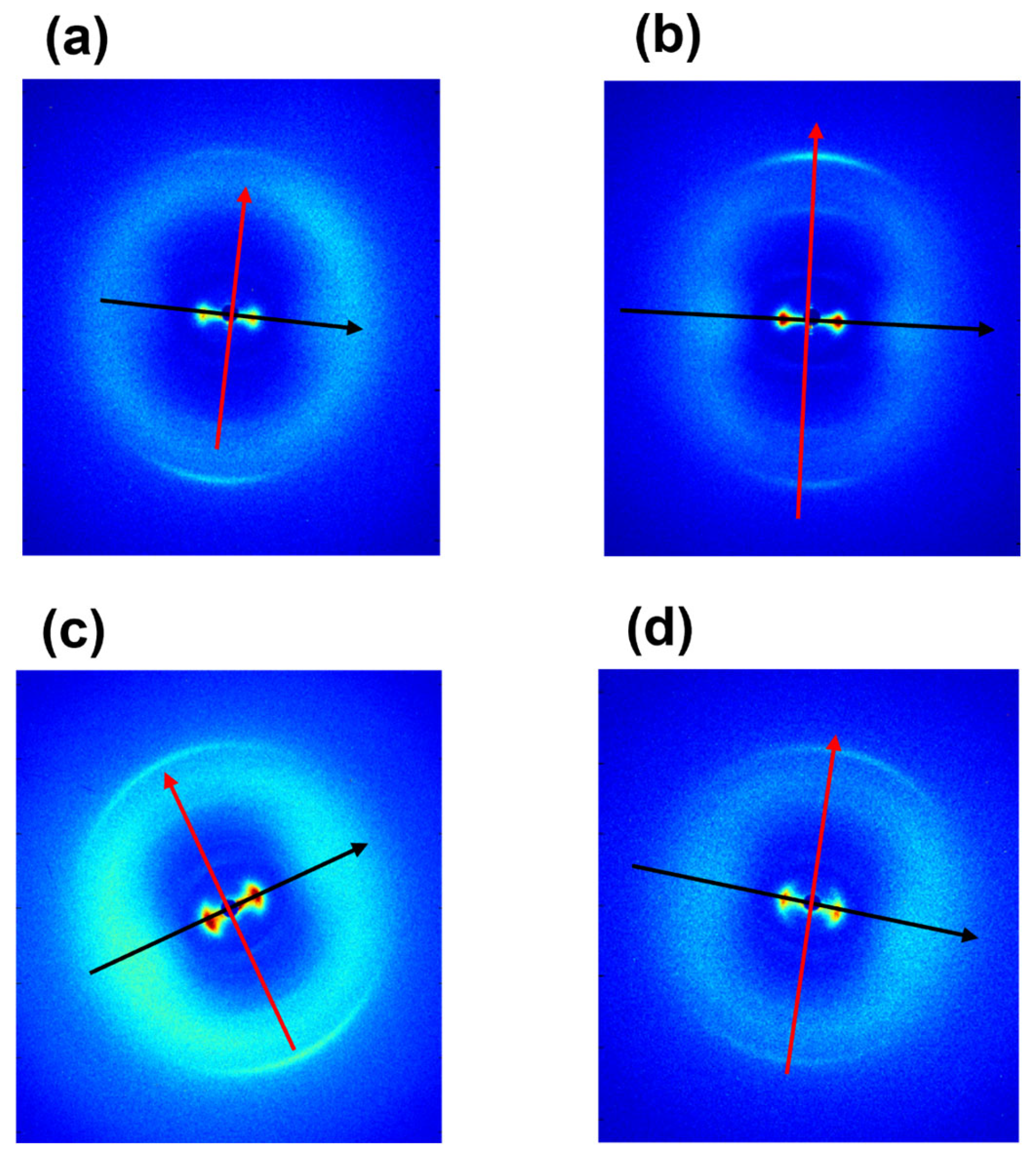
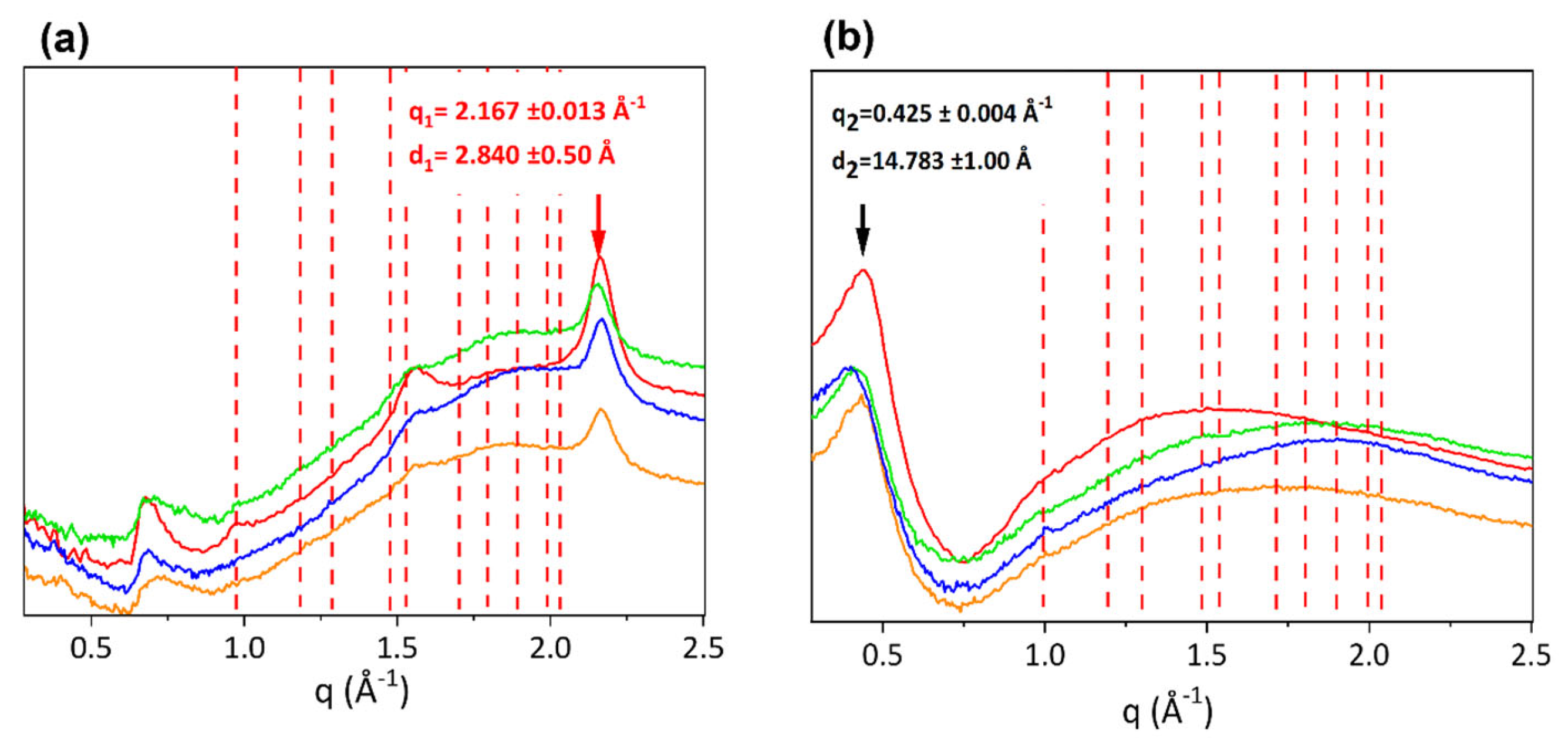
3.1. Ultrastructural Analyses (WAXS and SAXS)
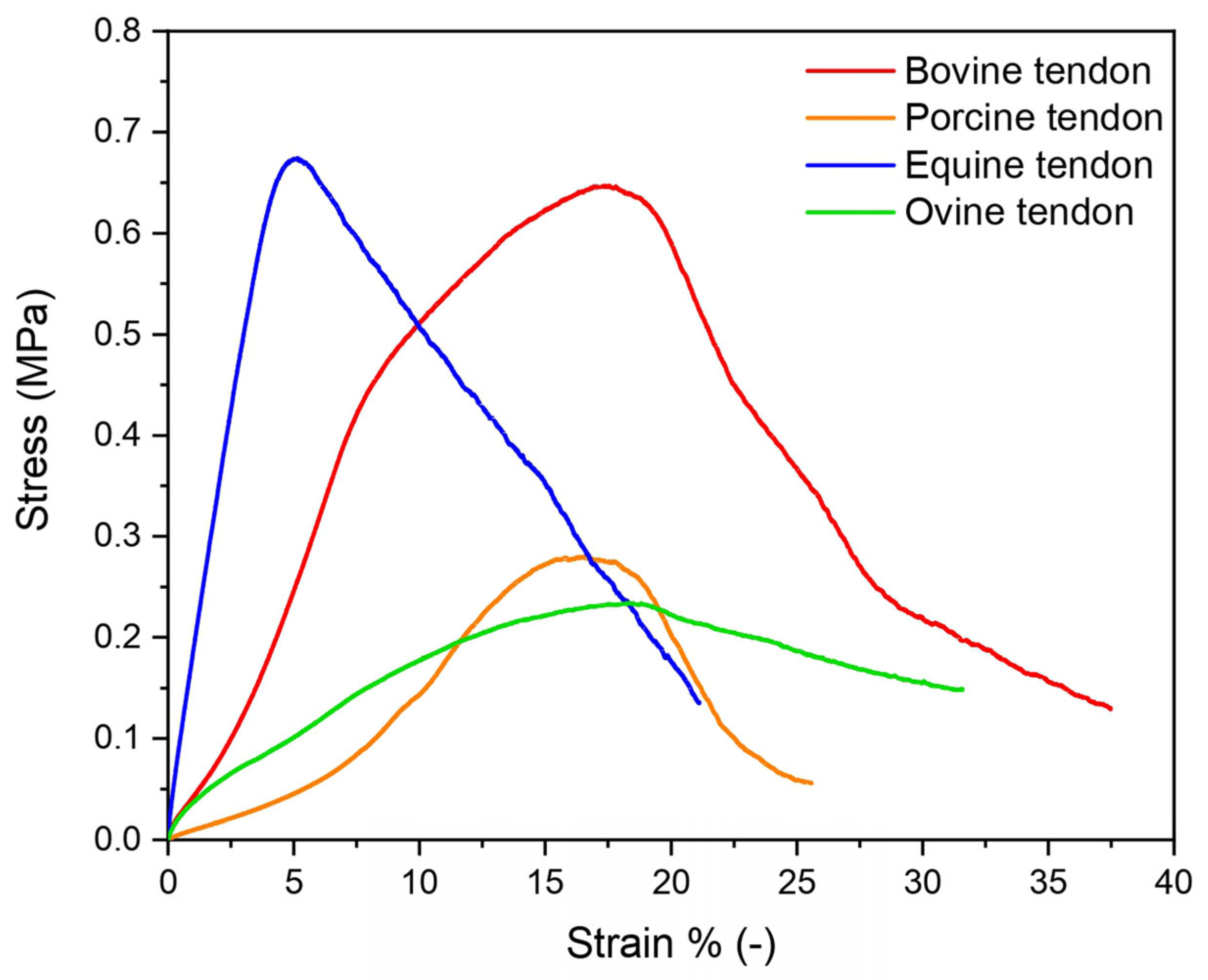
3.2. Mechanical Properties
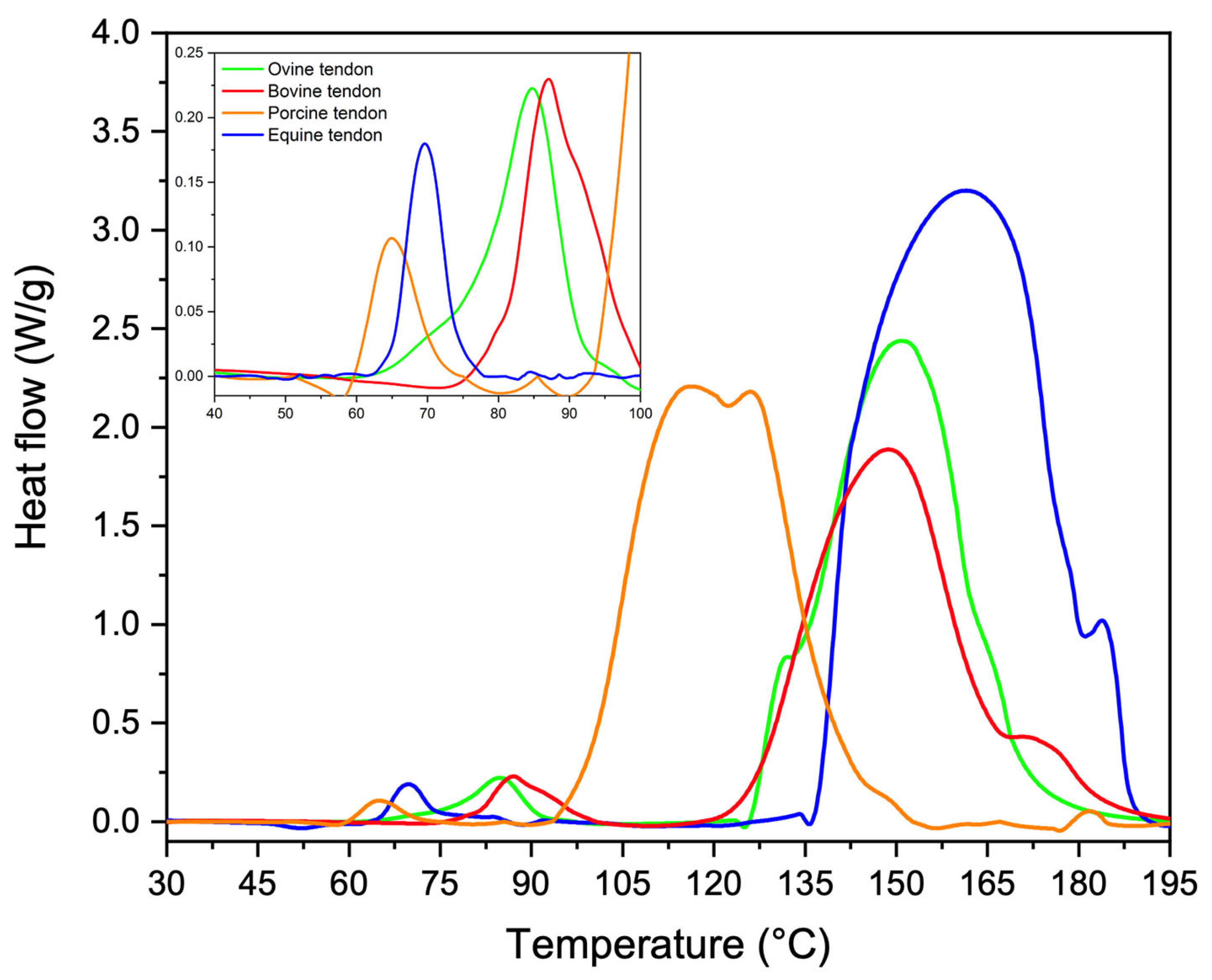
3.3. Thermal Properties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, Y.; Ko, H.; Kwon, I.K.; Shin, K. Extracellular Matrix Revisited: Roles in Tissue Engineering. Int. Neurourol. J. 2016, 20, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawelec, K.M.; Best, S.M.; Cameron, R.E. Collagen: A Network for Regenerative Medicine. J. Mater. Chem. B 2016, 4, 6484–6496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persikov, A.V.; Pillitteri, R.J.; Amin, P.; Schwarze, U.; Byers, P.H.; Brodsky, B. Stability Related Bias in Residues Replacing Glycines within the Collagen Triple Helix (Gly-Xaa-Yaa) in Inherited Connective Tissue Disorders. Hum. Mutat. 2004, 24, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [Green Version]
- Bella, J.; Brodsky, B.; Berman, H.M. Hydration Structure of a Collagen Peptide. Structure 1995, 3, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Rich, A.; Crick, F.H.C. The Molecular Structure of Collagen. J. Mol. Biol. 1961, 3, 483–506, IN1–IN4. [Google Scholar] [CrossRef]
- Okuyama, K.; Xu, X.; Iguchi, M.; Noguchi, K. Revision of Collagen Molecular Structure. Biopolymers 2006, 84, 181–191. [Google Scholar] [CrossRef]
- Terzi, A.; Gallo, N.; Bettini, S.; Sibillano, T.; Altamura, D.; Madaghiele, M.; De Caro, L.; Valli, L.; Salvatore, L.; Sannino, A.; et al. Sub- and Supramolecular X-ray Characterization of Engineered Tissues from Equine Tendon, Bovine Dermis, and Fish Skin Type-I Collagen. Macromol. Biosci. 2020, 20, 2000017. [Google Scholar] [CrossRef]
- Giannini, C.; De Caro, L.; Terzi, A.; Fusaro, L.; Altamura, D.; Diaz, A.; Lassandro, R.; Boccafoschi, F.; Bunk, O. Decellularized Pericardium Tissues at Increasing Glucose, Galactose and Ribose Concentrations and at Different Time Points Studied Using Scanning X-ray Microscopy. IUCrJ 2021, 8, 621–632. [Google Scholar] [CrossRef]
- Hadley, J.; Malik, N.; Meek, K. Collagen as a Model System to Investigate the Use of Aspirin as an Inhibitor of Protein Glycation and Crosslinking. Micron 2001, 32, 307–315. [Google Scholar] [CrossRef]
- Minary-Jolandan, M.; Yu, M.-F. Nanomechanical Heterogeneity in the Gap and Overlap Regions of Type I Collagen Fibrils with Implications for Bone Heterogeneity. Biomacromolecules 2009, 10, 2565–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzi, A.; Storelli, E.; Bettini, S.; Sibillano, T.; Altamura, D.; Salvatore, L.; Madaghiele, M.; Romano, A.; Siliqi, D.; Ladisa, M.; et al. Effects of Processing on Structural, Mechanical and Biological Properties of Collagen-Based Substrates for Regenerative Medicine. Sci. Rep. 2018, 8, 1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzi, A.; Gallo, N.; Bettini, S.; Sibillano, T.; Altamura, D.; Campa, L.; Natali, M.L.; Salvatore, L.; Madaghiele, M.; De Caro, L.; et al. Investigations of Processing–Induced Structural Changes in Horse Type-I Collagen at Sub and Supramolecular Levels. Front. Bioeng. Biotechnol. 2019, 7, 203. [Google Scholar] [CrossRef] [PubMed]
- Terzi, A.; Sibillano, T.; De Caro, L.; Altamura, D.; Gallo, N.; Natali, M.L.; Sannino, A.; Salvatore, L.; Blasi, F.S.; Corallo, A.; et al. WAXS and SAXS Investigation of Collagen-Rich Diet Effect on Multiscale Arrangement of Type I Collagen in Tilapia Skin Fed in Aquaponics Plant. Crystals 2022, 12, 700. [Google Scholar] [CrossRef]
- Fratzl, P.; Misof, K.; Zizak, I.; Rapp, G.; Amenitsch, H.; Bernstorff, S. Fibrillar Structure and Mechanical Properties of Collagen. J. Struct. Biol. 1998, 122, 119–122. [Google Scholar] [CrossRef]
- Screen, H.R.C.; Bader, D.L.; Lee, D.A.; Shelton, J.C. Local Strain Measurement within Tendon. Strain 2004, 40, 157–163. [Google Scholar] [CrossRef]
- Squire, J.M.; Freundlich, A. Direct Observation of a Transverse Periodicity in Collagen Fibrils. Nature 1980, 288, 410–413. [Google Scholar] [CrossRef]
- Silver, F.H.; Kato, Y.P.; Ohno, M.; Wasserman, A.J. Analysis of Mammalian Connective Tissue: Relationship between Hierarchical Structures and Mechanical Properties. J. Long Term Eff. Med. Implant. 1992, 2, 165–198. [Google Scholar]
- Rosager, S.; Aagaard, P.; Dyhre-Poulsen, P.; Neergaard, K.; Kjaer, M.; Magnusson, S.P. Load-Displacement Properties of the Human Triceps Surae Aponeurosis and Tendon in Runners and Non-Runners: Physical Activity and Human Tendon. Scand. J. Med. Sci. Sport. 2002, 12, 90–98. [Google Scholar] [CrossRef]
- Buchanan, C.I.; Marsh, R.L. Effects of Long-Term Exercise on the Biomechanical Properties of the Achilles Tendon of Guinea Fowl. J. Appl. Physiol. 2001, 90, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Altamura, D.; Lassandro, R.; Vittoria, F.A.; De Caro, L.; Siliqi, D.; Ladisa, M.; Giannini, C. X-ray Microimaging Laboratory (XMI-LAB). J. Appl. Cryst. 2012, 45, 869–873. [Google Scholar] [CrossRef]
- Siliqi, D.; De Caro, L.; Ladisa, M.; Scattarella, F.; Mazzone, A.; Altamura, D.; Sibillano, T.; Giannini, C. SUNBIM: A Package for X-ray Imaging of Nano- and Biomaterials Using SAXS, WAXS, GISAXS and GIWAXS Techniques. J. Appl. Cryst. 2016, 49, 1107–1114. [Google Scholar] [CrossRef]
- Bunk, O.; Bech, M.; Jensen, T.H.; Feidenhans’l, R.; Binderup, T.; Menzel, A.; Pfeiffer, F. Multimodal X-ray Scatter Imaging. New J. Phys. 2009, 11, 123016. [Google Scholar] [CrossRef] [Green Version]
- Henrich, B.; Bergamaschi, A.; Broennimann, C.; Dinapoli, R.; Eikenberry, E.F.; Johnson, I.; Kobas, M.; Kraft, P.; Mozzanica, A.; Schmitt, B. PILATUS: A Single Photon Counting Pixel Detector for X-ray Applications. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2009, 607, 247–249. [Google Scholar] [CrossRef]
- Lutz-Bueno, V.; Arboleda, C.; Leu, L.; Blunt, M.J.; Busch, A.; Georgiadis, A.; Bertier, P.; Schmatz, J.; Varga, Z.; Villanueva-Perez, P.; et al. Model-Free Classification of X-Ray Scattering Signals Applied to Image Segmentation. J. Appl. Cryst. 2018, 51, 1378–1386. [Google Scholar] [CrossRef] [Green Version]
- Gallo, N.; Terzi, A.; Sibillano, T.; Giannini, C.; Masi, A.; Sicuro, A.; Blasi, F.S.; Corallo, A.; Pennetta, A.; De Benedetto, G.E.; et al. Age-Related Properties of Aquaponics-Derived Tilapia Skin (Oreochromis Niloticus): A Structural and Compositional Study. Int. J. Mol. Sci. 2023, 24, 1938. [Google Scholar] [CrossRef]
- Gallo, N.; Natali, M.L.; Quarta, A.; Gaballo, A.; Terzi, A.; Sibillano, T.; Giannini, C.; De Benedetto, G.E.; Lunetti, P.; Capobianco, L.; et al. Aquaponics-Derived Tilapia Skin Collagen for Biomaterials Development. Polymers 2022, 14, 1865. [Google Scholar] [CrossRef] [PubMed]
- Gallo, N.; Natali, M.L.; Curci, C.; Picerno, A.; Gallone, A.; Vulpi, M.; Vitarelli, A.; Ditonno, P.; Cascione, M.; Sallustio, F.; et al. Analysis of the Physico-Chemical, Mechanical and Biological Properties of Crosslinked Type-I Collagen from Horse Tendon: Towards the Development of Ideal Scaffolding Material for Urethral Regeneration. Materials 2021, 14, 7648. [Google Scholar] [CrossRef]
- Salvatore, L.; Calò, E.; Bonfrate, V.; Pedone, D.; Gallo, N.; Natali, M.L.; Sannino, A.; Madaghiele, M. Exploring the Effects of the Crosslink Density on the Physicochemical Properties of Collagen-Based Scaffolds. Polym. Test. 2021, 93, 106966. [Google Scholar] [CrossRef]
- Sun, L.; Hou, H.; Li, B.; Zhang, Y. Characterization of Acid- and Pepsin-Soluble Collagen Extracted from the Skin of Nile Tilapia (Oreochromis Niloticus). Int. J. Biol. Macromol. 2017, 99, 8–14. [Google Scholar] [CrossRef]
- Burgio, V.; Civera, M.; Rodriguez Reinoso, M.; Pizzolante, E.; Prezioso, S.; Bertuglia, A.; Surace, C. Mechanical Properties of Animal Tendons: A Review and Comparative Study for the Identification of the Most Suitable Human Tendon Surrogates. Processes 2022, 10, 485. [Google Scholar] [CrossRef]
- Robinson, P.S.; Lin, T.W.; Reynolds, P.R.; Derwin, K.A.; Iozzo, R.V.; Soslowsky, L.J. Strain-Rate Sensitive Mechanical Properties of Tendon Fascicles from Mice with Genetically Engineered Alterations in Collagen and Decorin. J. Biomech. Eng. 2004, 126, 252–257. [Google Scholar] [CrossRef]
- Chambers, N.C.; Herod, T.W.; Veres, S.P. Ultrastructure of Tendon Rupture Depends on Strain Rate and Tendon Type. J. Orthop. Res. 2018, 36, 2842–2850. [Google Scholar] [CrossRef] [Green Version]
- Samouillan, V.; Delaunay, F.; Dandurand, J.; Merbahi, N.; Gardou, J.-P.; Yousfi, M.; Gandaglia, A.; Spina, M.; Lacabanne, C. The Use of Thermal Techniques for the Characterization and Selection of Natural Biomaterials. JFB 2011, 2, 230–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatore, L.; Gallo, N.; Aiello, D.; Lunetti, P.; Barca, A.; Blasi, L.; Madaghiele, M.; Bettini, S.; Giancane, G.; Hasan, M.; et al. An Insight on Type I Collagen from Horse Tendon for the Manufacture of Implantable Devices. Int. J. Biol. Macromol. 2020, 154, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, L.; Gallo, N.; Natali, M.L.; Terzi, A.; Sannino, A.; Madaghiele, M. Mimicking the Hierarchical Organization of Natural Collagen: Toward the Development of Ideal Scaffolding Material for Tissue Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 644595. [Google Scholar] [CrossRef]
- Sasaki, N.; Odajima, S. Stress-Strain Curve and Young’s Modulus of a Collagen Molecule as Determined by the X-ray Diffraction Technique. J. Biomech. 1996, 29, 655–658. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples |
|---|
| Bovine tendon |
| Porcine tendon |
| Extensor equine tendon |
| Ovine tendon |
| Equatorial Direction | Meridional Direction | ||||
|---|---|---|---|---|---|
| Tendon | Lateral q2-Spacing (Å−1) | Lateral d2-Spacing (Å) | FWHM (Å−1) | Axial Turn q1-Spacing (Å−1) | Axial Turn d1-Spacing (Å) |
| Extensor equine | 0.397 ± 0.001 | 15.826 ± 0.040 | 0.124 ± 0.003 | 2.169 ± 0.001 | 2.896 ± 0.001 |
| Bovine | 0.429 ± 0.001 | 14.646 ± 0.034 | 0.114 ± 0.004 | 2.162 ± 0.001 | 2.906 ± 0.001 |
| Ovine | 0.415 ± 0.001 | 15.140 ± 0.040 | 0.134 ± 0.002 | 2.156 ± 0.001 | 2.914 ± 0.001 |
| Porcine | 0.426 ± 0.001 | 14.749 ± 0.034 | 0.111 ± 0.002 | 2.168 ± 0.001 | 2.898 ± 0.001 |
| Tendon | E (MPa) | σ max (MPa) | εr % (−) |
|---|---|---|---|
| Equine extensor | 13.8 ± 3.0 | 0.6 ± 0.2 | 4.9 ± 1.5 |
| Bovine | 4.8 ± 1.2 | 0.7 ± 0.1 | 20.3 ± 4.7 |
| Ovine | 2.7 ± 1.5 | 0.3 ± 0.1 | 17.1 ± 3.2 |
| Porcine | 0.9 ± 0.2 | 0.3 ± 0.1 | 14.8 ± 3.1 |
| Tendon | TI (°C) | ΔHI (J/g) | TI (°C) | ΔHI (J/g) |
|---|---|---|---|---|
| Equine extensor | 70 ± 10 | 16 ± 4 | 171 ± 19 | 1422 ± 203 |
| Bovine | 86 ± 9 | 33 ± 7 | 151 ± 18 | 673 ± 84 |
| Ovine | 84 ± 8 | 23 ± 8 | 154 ± 22 | 789 ± 92 |
| Porcine | 65 ± 12 | 9 ± 3 | 112 ± 17 | 912 ± 63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terzi, A.; Gallo, N.; Sibillano, T.; Altamura, D.; Masi, A.; Lassandro, R.; Sannino, A.; Salvatore, L.; Bunk, O.; Giannini, C.; et al. Travelling through the Natural Hierarchies of Type I Collagen with X-rays: From Tendons of Cattle, Horses, Sheep and Pigs. Materials 2023, 16, 4753. https://doi.org/10.3390/ma16134753
Terzi A, Gallo N, Sibillano T, Altamura D, Masi A, Lassandro R, Sannino A, Salvatore L, Bunk O, Giannini C, et al. Travelling through the Natural Hierarchies of Type I Collagen with X-rays: From Tendons of Cattle, Horses, Sheep and Pigs. Materials. 2023; 16(13):4753. https://doi.org/10.3390/ma16134753
Chicago/Turabian StyleTerzi, Alberta, Nunzia Gallo, Teresa Sibillano, Davide Altamura, Annalia Masi, Rocco Lassandro, Alessandro Sannino, Luca Salvatore, Oliver Bunk, Cinzia Giannini, and et al. 2023. "Travelling through the Natural Hierarchies of Type I Collagen with X-rays: From Tendons of Cattle, Horses, Sheep and Pigs" Materials 16, no. 13: 4753. https://doi.org/10.3390/ma16134753