
The Structure and Function of Biomaterial Endolysin EFm1 from E. faecalis Phage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Phage and Strains
2.2. Plasmid Construction
2.3. Protein Expression and Purification
2.4. Crystallization and Structure Determination
2.5. Lytic Activity Experiment
2.6. CRISPR Interference Experiments
2.7. Statistical Analysis and Figure Preparation
3. Results
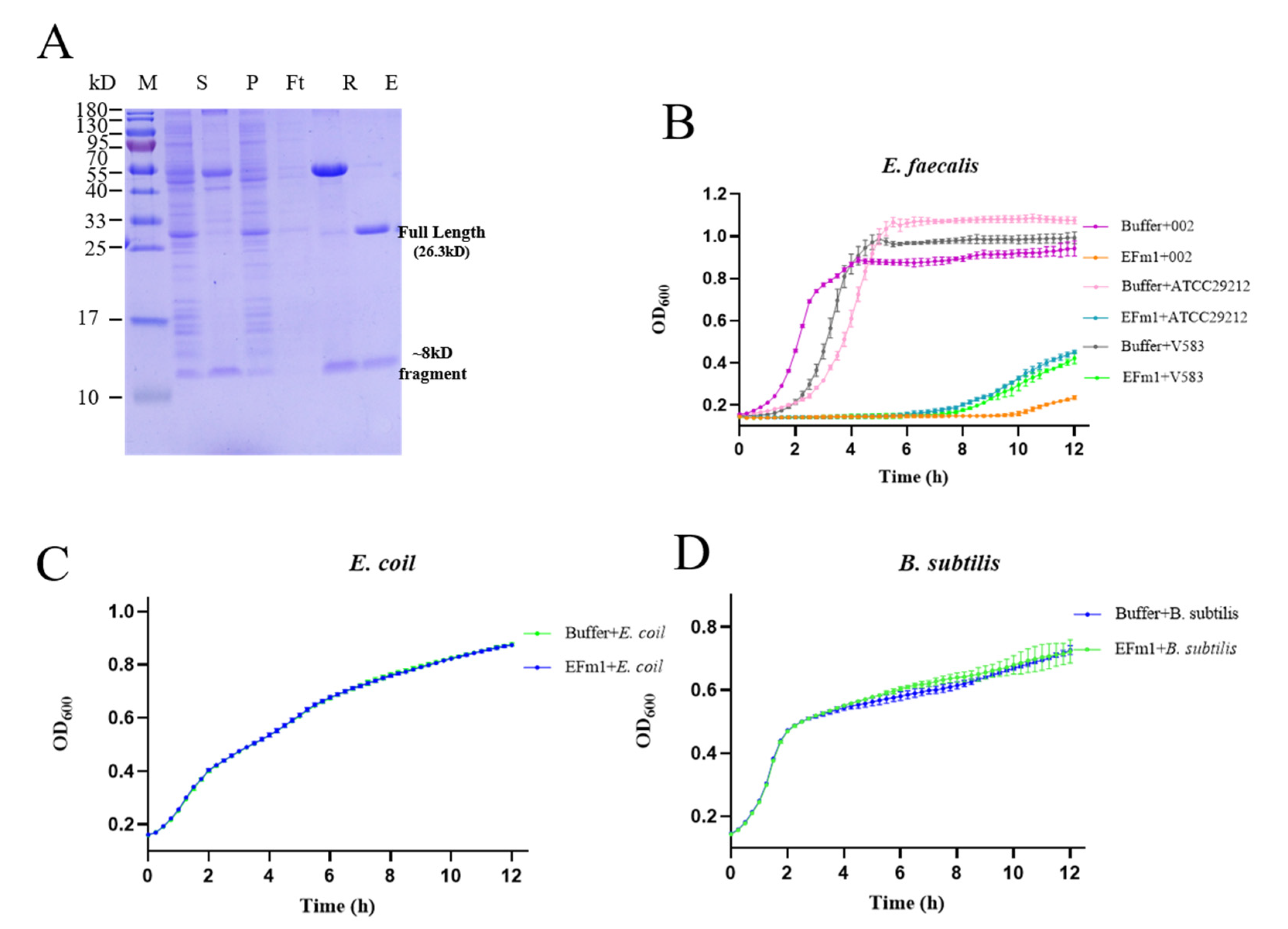
3.1. Lytic Activity of EFm1
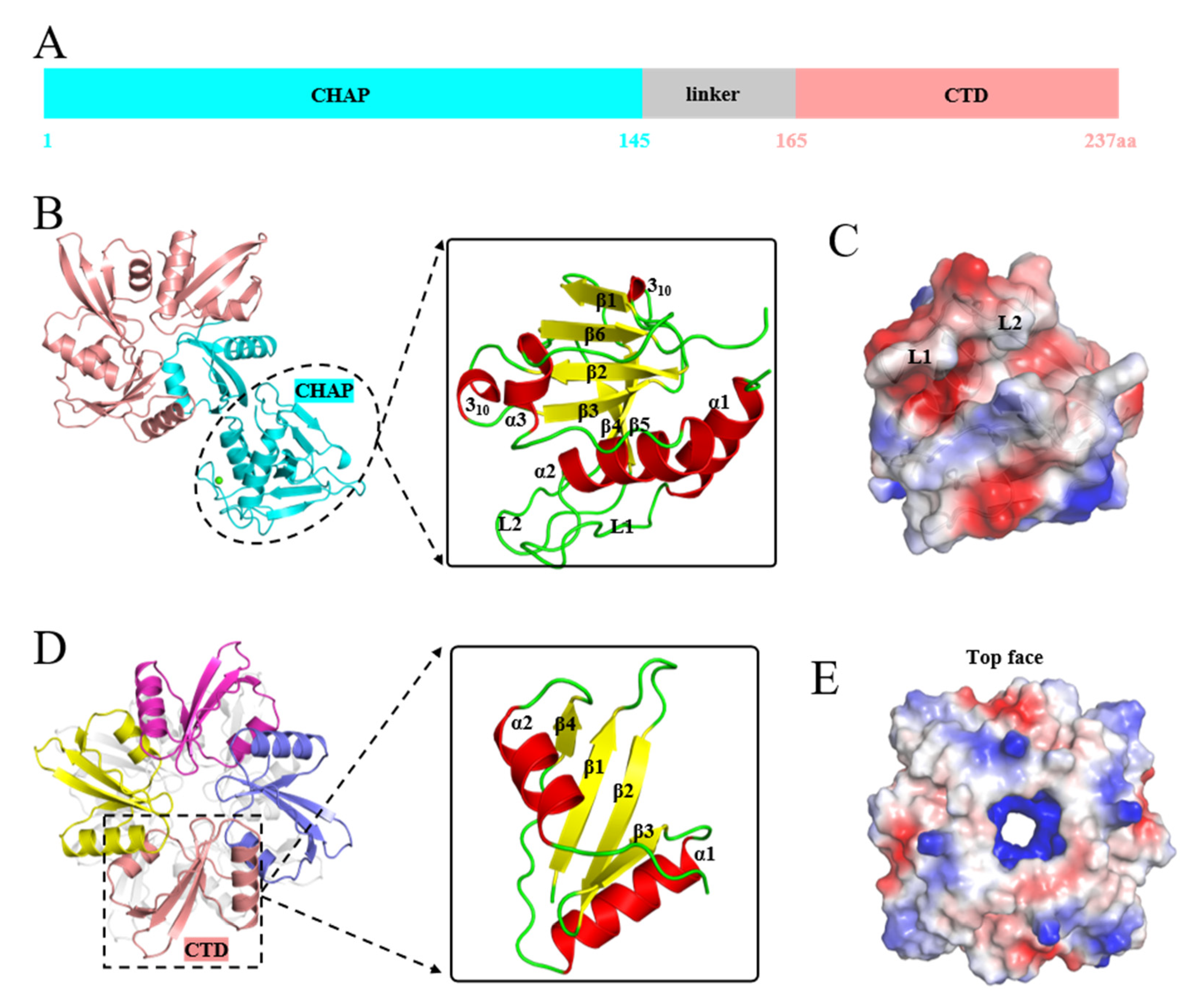
3.2. Structures of EFm1D166Q and EFm1166–237 aa
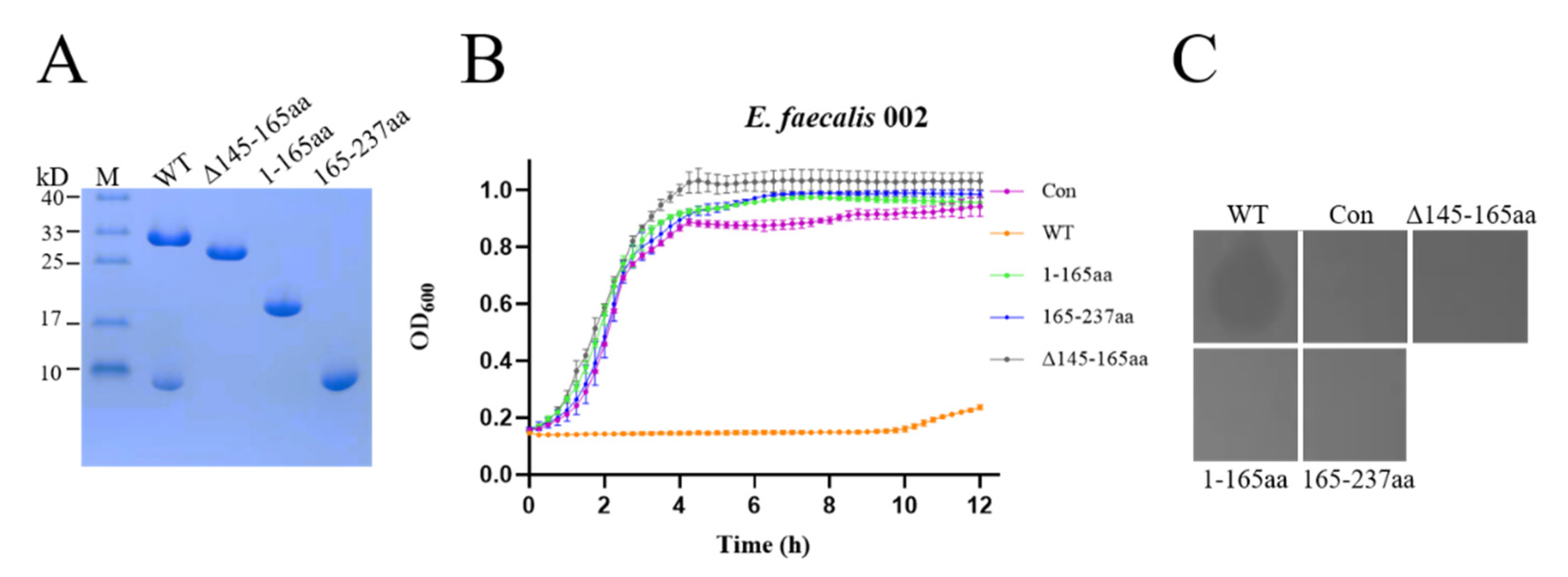
3.3. The Effect of the NTD, CTD and Linker on the Lytic Activity of EFm1
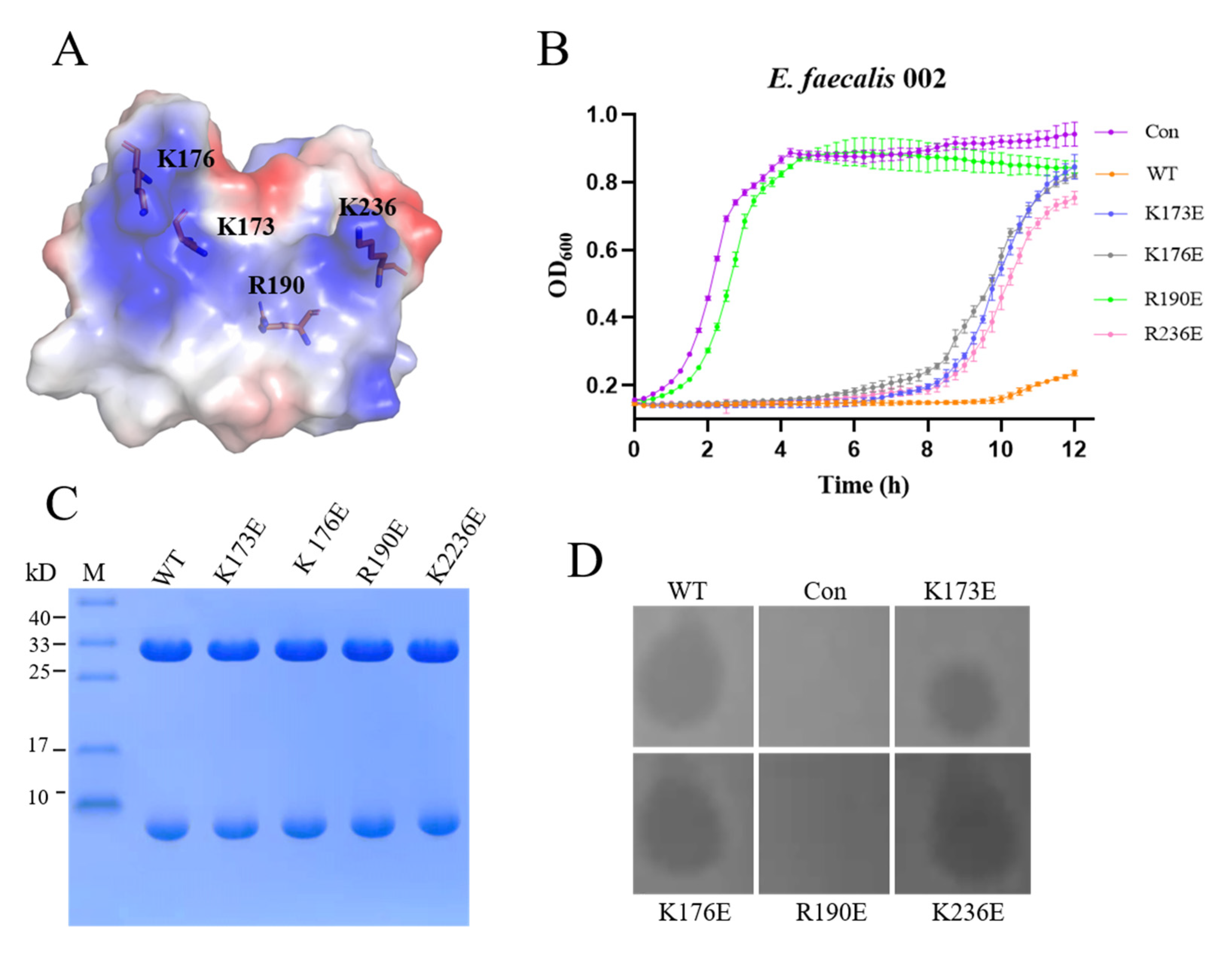
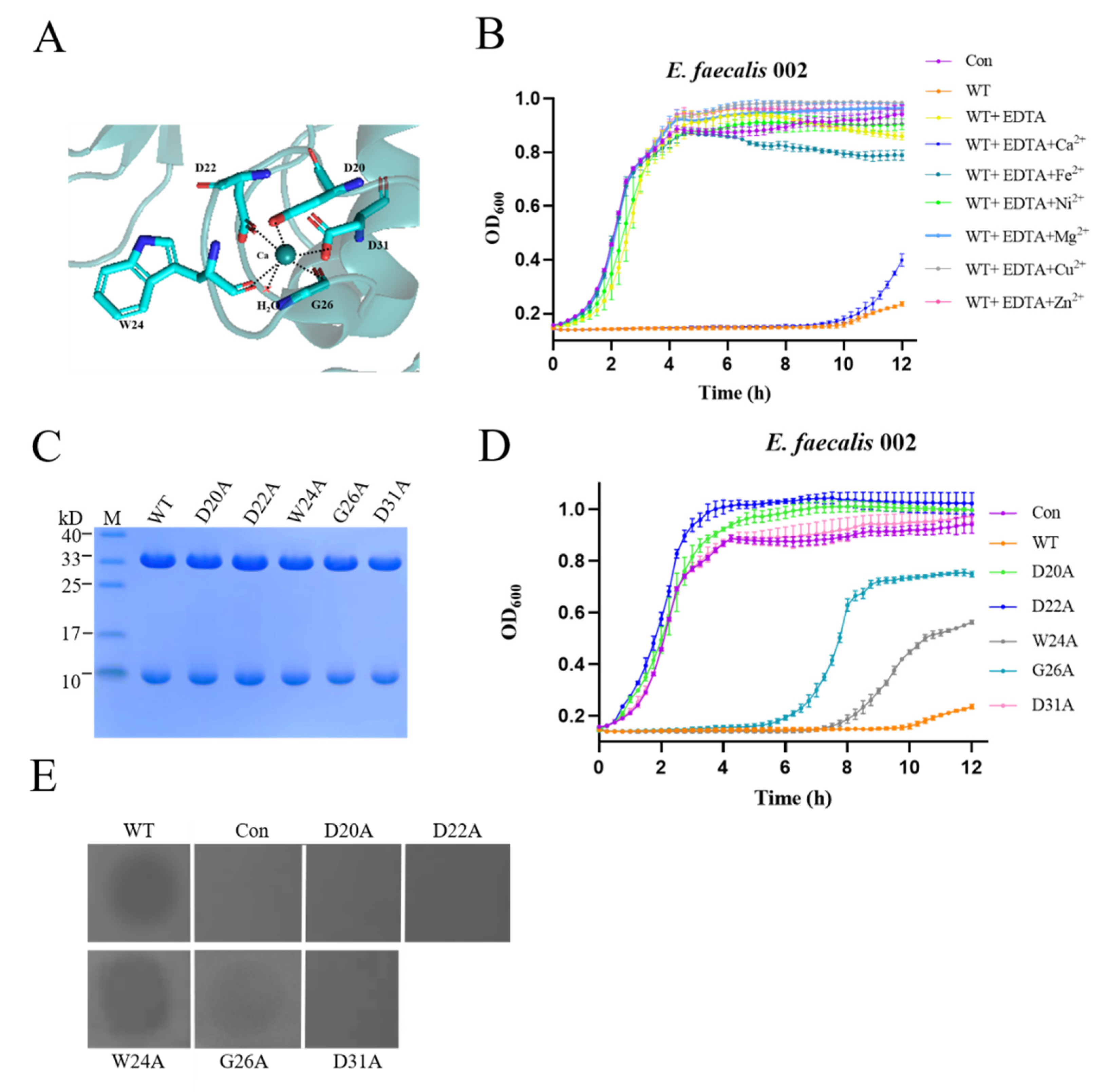
3.4. Binding Sites and Iron Channel of EFm1
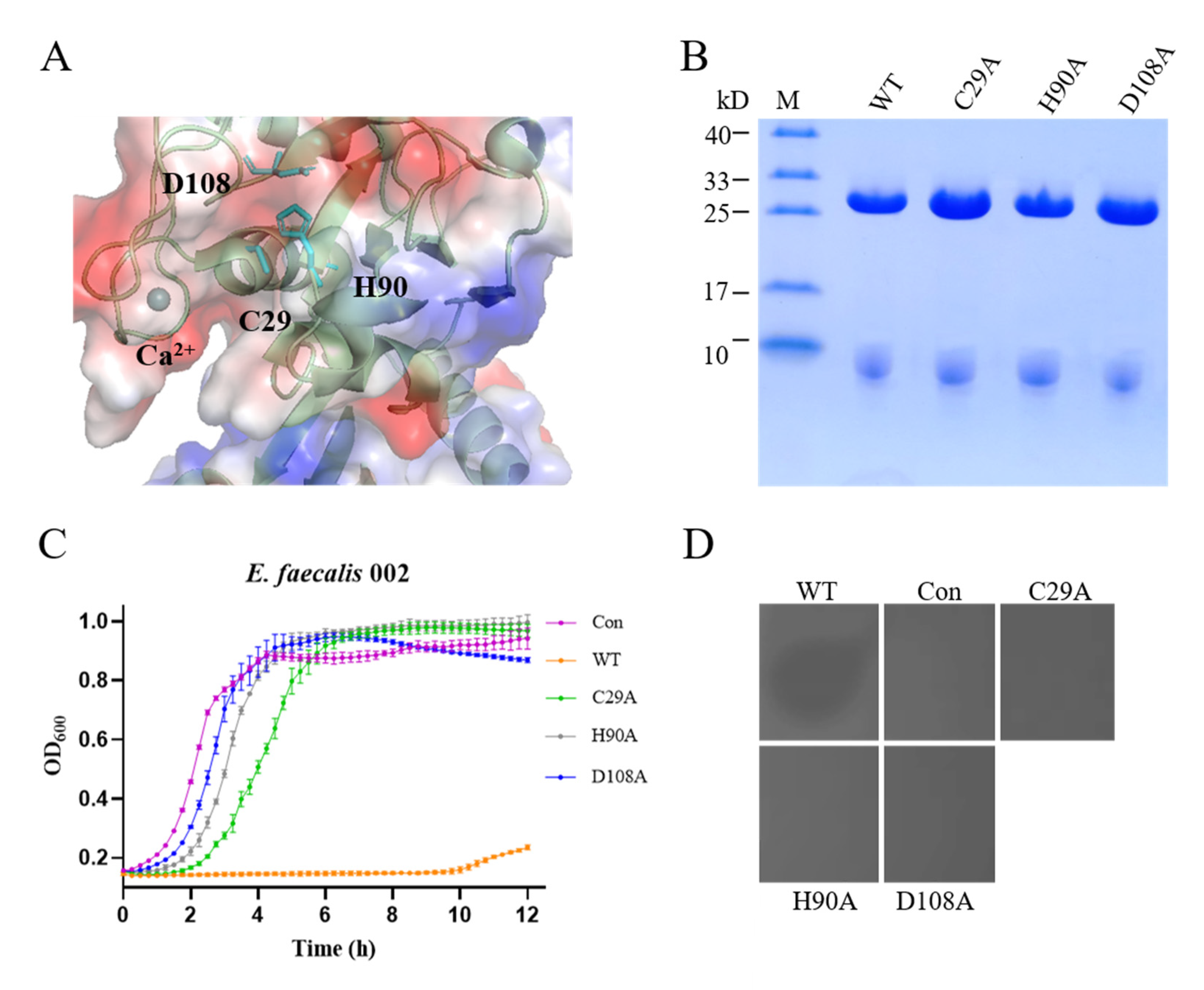
3.5. Lytic Activity of the CHAP Domain of EFm1
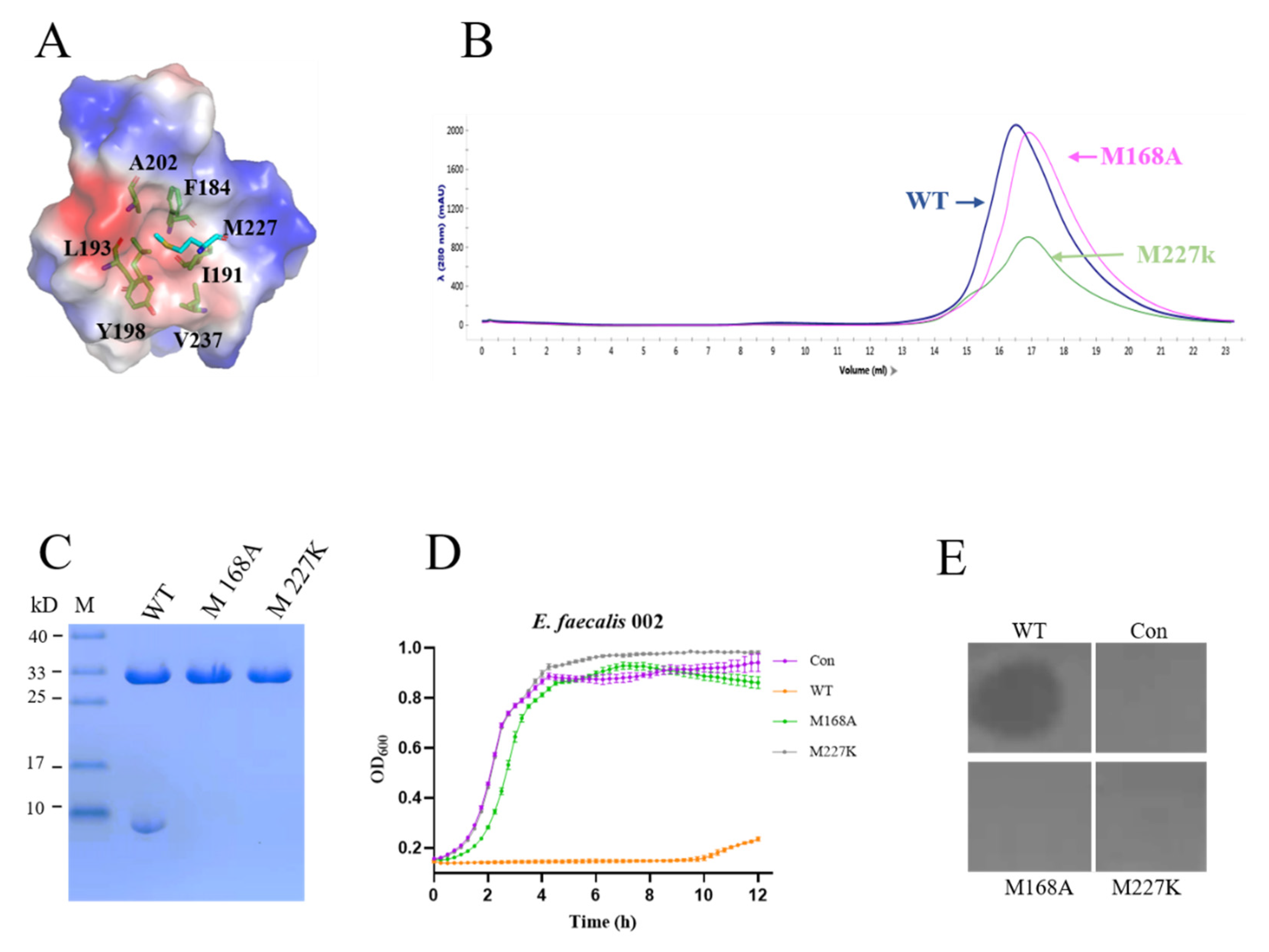
3.6. Lytic Activity of the Tetramer of EFm1
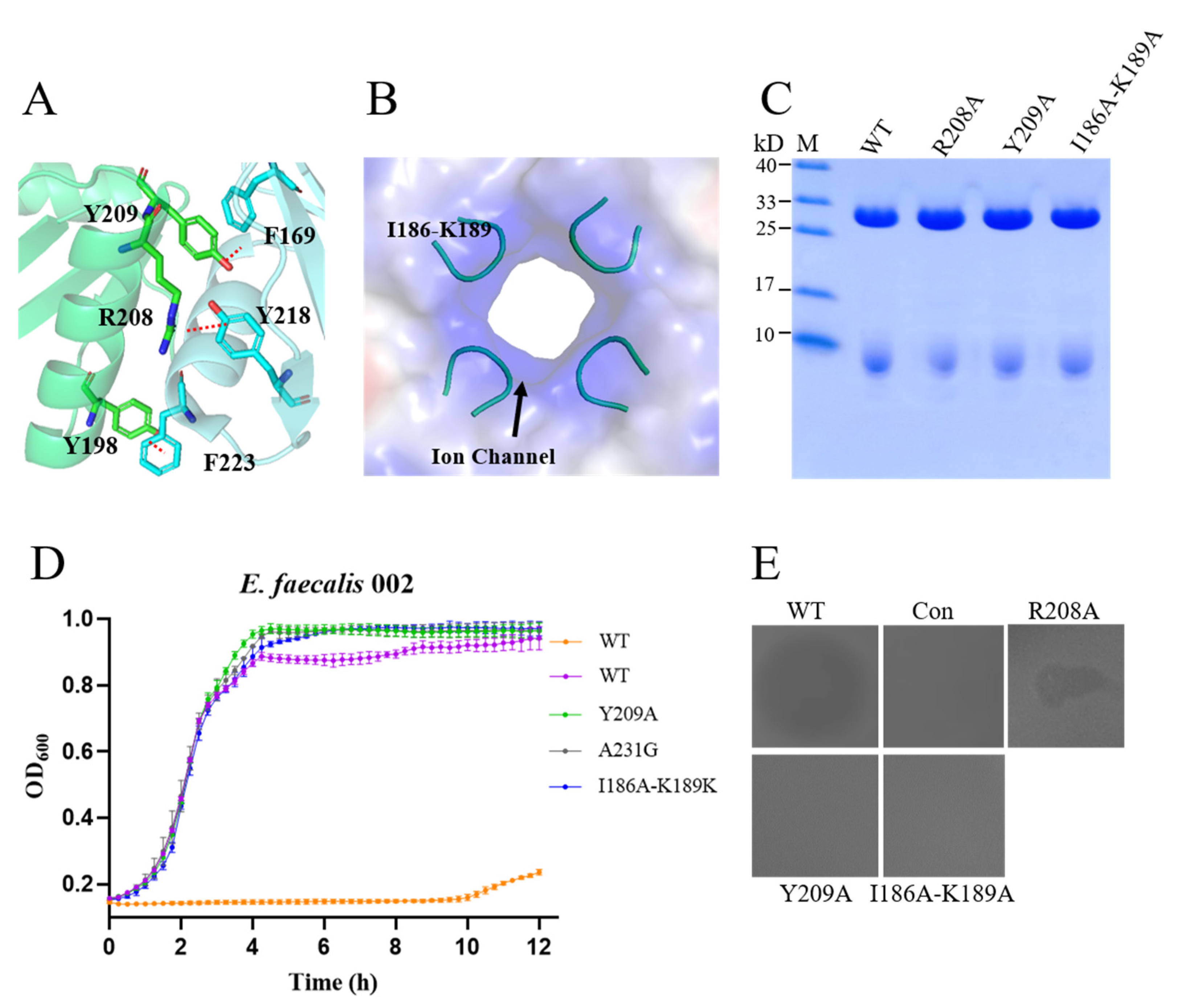
3.7. Interaction of the Monomer and Ion Channel in the CTD
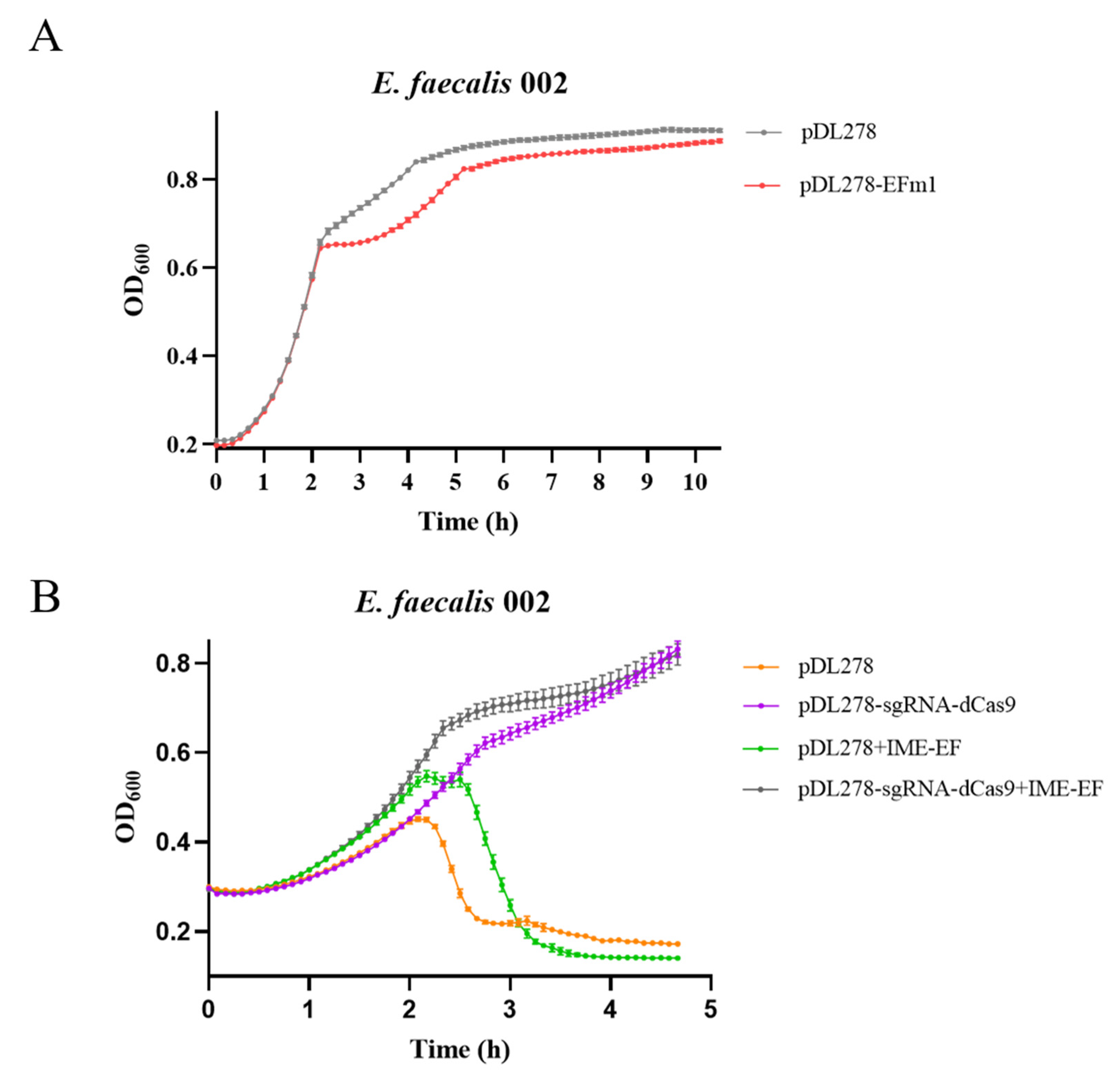
3.8. Expression In Vivo and Knockdown of EFm1 in 002
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Philippe, J.G.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Huang, X.; Ding, S.; Wang, Y.; Shen, J.; Zhu, K. A broad-spectrum antibiotic adjuvant reverses multidrug-resistant gram-negative pathogens. Nat. Microbiol. 2020, 5, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Pöntinen, A.K.; Top, J.; Arredondo-Alonso, S.; Tonkin-Hill, G.; Freitas, A.R.; Novais, C.; Gladstone, R.A.; Pesonen, M.; Meneses, R.; Pesonen, H.; et al. Apparent nosocomial adaptation of Enterococcus faecalis predates the modern hospital era. Nat. Commun. 2021, 12, 1523. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Singh, M.; Kim, S.J.; Schaefer, J. Characterization of the tertiary structure of the peptidoglycan of Enterococcus faecalis. Biochim. Biophys. Acta. 2017, 1859, 2171–2180. [Google Scholar] [CrossRef]
- Yoon, Y.K.; Sim, H.S.; Kim, J.Y.; Park, D.W.; Sohn, J.W.; Roh, K.H.; Lee, S.E.; Kimm, M.J. Epidemiology and control of an outbreak of vancomycin-resistant enterococci in the intensive care units. Yonsei Med. J. 2009, 50, 637–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbagh, P.; Rajabnia, M.; Maali, A.; Ferdosi-Shahandashti, E. Integron and its role in antimicrobial resistance: A literature review on some bacterial pathogens. Iran. J. Basic Med. Sci. 2021, 24, 136–142. [Google Scholar] [PubMed]
- Hegstad, K.; Mikalsen, T.; Coque, T.M.; Werner, G.; Sundsfjord, A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium. Clin. Microbiol. Infec. 2010, 16, 541–554. [Google Scholar] [CrossRef]
- Biswas, B.; Adhya, S.; Washart, P.; Paul, B.; Trostel, A.N.; Powell, B.; Carlton, R.; Merril1, C.R. Bacteriophage therapy rescues mice bacteremic from a clinical isolate of vancomycin-resistant Enterococcus faecium. Infect. Immun. 2002, 70, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Fischetti, V.A. Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens. Int. J. Med. Microbiol. 2010, 300, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Gondil, V.S.; Harjai, K.; Chhibber, S. Endolysins as emerging alternative therapeutic agents to counter drug-resistant infections. Int. J. Antimicrob. Agents 2020, 55, 105844. [Google Scholar] [CrossRef]
- Chang, Y. Bacteriophage-derived endolysins applied as potent biocontrol agents to enhance food safety. Microorganisms 2020, 8, 724. [Google Scholar] [CrossRef]
- Gu, J.; Xi, H.; Cheng, M.; Han, W. Phage-derived lysins as therapeutic agents against multidrug-resistant Enterococcus faecalis. Future Microbiol. 2018, 13, 275–278. [Google Scholar] [CrossRef] [Green Version]
- Hathaway, H.; Ajuebor, J.; Stephens, L.; Coffey, A.; Potter, U.; Sutton, J.M.; Jenkins, A.T.A. Thermally triggered release of the bacteriophage endolysin CHAP(K) and the bacteriocin lysostaphin for the control of methicillin resistant Staphylococcus aureus (MRSA). J. Control. Release 2017, 245, 108–115. [Google Scholar] [CrossRef]
- Zhang, W.; Mi, Z.; Yin, X.; Fan, H.; An, X.; Zhang, Z.; Chen, J.; Tong, Y. Characterization of enterococcus faecalis phage IME-EF1 and its endolysin. PLoS ONE 2013, 8, e80435. [Google Scholar] [CrossRef]
- Cheng, M.; Zhang, Y.; Li, X.; Liang, J.; Gu, J. Endolysin LysEF-P10 shows potential as an alternative treatment strategy for multi-drug-resistant Enterococcus faecalis infections. Sci. Rep. 2017, 7, 10164. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Zhen, X.; Zhou, H.; Zhao, F.; Fan, C.; Perčulija, V.; Tong, Y.; Mi, Z.; Ouyang, S. Structural and functional insights into a novel two-component endolysin encoded by a single gene in Enterococcus faecalis phage. PLoS Pathog. 2020, 16, e1008394. [Google Scholar] [CrossRef]
- Dou, C.; Xiong, J.; Gu, Y.; Yin, K.; Wang, J.; Hu, Y.; Zhou, D.; Fu, X.; Qi, S.; Zhu, X.; et al. Structural and functional insights into the regulation of the lysis-lysogeny decision in viral communities. Nat. Microbiol. 2018, 3, 1285–1294. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Cooper, D.R.; Derewenda, Z.S.; Eisenberg, D. Toward rational protein crystallization: A Web server for the design of crystallizable protein variants. Protein Sci. 2007, 16, 1569–1576. [Google Scholar] [CrossRef] [Green Version]
- Slabinski, L.; Jaroszewski, L.; Rychlewski, L.; Wilson, I.A.; Lesley, S.A.; Godzik, A. XtalPred: A web server for prediction of protein crystallizability. Bioinformatics 2007, 15, 3403–3405. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Hu, Y.; Gu, Y.; Huang, J.; He, J.; Luo, L.; Yang, Y.; Yin, S.; Dou, C.; Wang, T.; et al. Deciphering the regulatory and catalytic mechanisms of an unusual SAM-dependent enzyme. Signal Transduct. Target. Ther. 2019, 24, 17. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, F.; Huang, S.; Sun, B.; Zhang, K.; Liu, K.; Wang, Z.; Xu, C.; Wang, S.; Yang, L. The macromolecular crystallography beamline of SSRF. Nucl. Sci. Tech. 2015, 26, 8–13. [Google Scholar] [CrossRef]
- Zhang, J.; Mccabe, K.A.; Bell, C.E. Crystal structures of λ exonuclease in complex with DNA suggest an electrostatic ratchet mechanism for processivity. Proc. Natl. Acad. Sci. USA 2011, 108, 11872–11877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Grossekunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Mooers, B.H.M. Shortcuts for faster image creation in PyMOL. Protein Sci. 2020, 29, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motulsky, H.J. Common misconceptions about data analysis and. Pharmacol. Res. Perspect. 2015, 3, e00093. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.; Buckle, A.M.; Mitchell, M.S.; Hoopes, J.T.; Gallagher, D.T.; Heselpoth, R.D.; Shen, Y.; Reboul, C.F.; Law, R.H.P.; Fischetti, V.A.; et al. X-ray crystal structure of the streptococcal specific phage lysin PlyC. Proc. Natl. Acad. Sci. USA 2012, 109, 12752–12757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eijsink, V.G.H.; Matthews, B.W.; Vriend, G. The role of calcium ions in the stability and instability of a thermolysin-like protease. Protein Sci. 2011, 20, 1346–1355. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Feng, Y.; Feng, X.; Sun, C.; Lei, L.; Ding, W.; Niu, F.; Jiao, L.; Yang, M.; Li, Y.; et al. Structural and biochemical characterization reveals LysGH15 as an unprecedented "EF-Hand-Like" calcium-binding phage lysin. PloS Pathog. 2014, 10, e1004109. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Yun, Y.S.; Koo, H.M.; Kim, Y.S.; Choi, K.Y.; Oh, B.H. Characterization of a novel ser-cisser-lys catalytic triad in comparison with the classical ser-his-asp triad. J. Biol. Chem. 2003, 278, 24937. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.H.; Hsu, Y.H.; Wu, W.J.; Chang, K.C.; Yeh, C.S. Phage digestion of a bacterial capsule imparts resistance to two antibiotic agents. Microorganisms 2021, 9, 794. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Schuch, R.; Chahales, P.; Zhu, S.; Fischetti, V.A. PlyC: A multimeric bacteriophage lysin. Proc. Natl. Acad. Sci. USA 2006, 103, 10765–10770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, A.; Bott, R.; Vonrhein, C.; Bricogne, G.; Power, S.; Day, A.G. A novel combination of two classic catalytic schemes. J. Mol. Biol. 2002, 320, 303–309. [Google Scholar] [CrossRef]
- Ortega, M.A.; Hao, Y.; Zhang, Q.; Walker, M.C.; Donk, W.; Nair, S.K. Structure and mechanism of the tRNA-dependent lantibiotic dehydratase NisB. Nature 2015, 517, 509–512. [Google Scholar] [CrossRef]
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Lee, J.H.; Yoon, H.; Kang, D.H.; Ryu, S. Genomic investigation of lysogen formation and host lysis systems of the Salmonella temperate bacteriophage SPN9CC. Appl. Environ. Microbiol. 2014, 80, 374–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ to 3′) | Usage |
|---|---|---|
| EFm11–165 aa-F | AGACGATTAGCTCGAGCGGCCGCATCGT | pGEX-EFm11–165 aa |
| EFm11–165 aa-R | GCTCGAGCTAATCGTCTCCTTTAAATTGTCCTAGATT | |
| EFm1165–237 aa-F | CTGGGATCCGATGATATTATGTTCATCTATTACAAACGCAC | pGEX-EFm1165–237 aa |
| EFm1165–237 aa-R | ATATCATCGGATCCCAGGGGCCCCTG | |
| EFm1△145–165 aa-F | GCGTTTGTAATAGATGAACATAATATCTGAAGCAGCTTCGTAAGG | pGEX-EFm1△145–165 aa |
| EFm1△145–165 aa-R | GATATTATGTTCATCTATTACAAACGCACTAAGCAAGGA | |
| EFm1D166Q-F | GAGACGATCAAATTATGTTCATCTATTACAAACGCACTAA | pGEX-EFm1D166Q |
| EFm1D166Q-R | CATAATTTGATCGTCTCCTTTAAATTGTCCTAGATT | |
| EFm1C29A-F | TACGCAAGCGATGGACTTGACAGTAGACGTTATGCA | pGEX-EFm1C29A |
| EFm1C29A-R | AGTCCATCGCTTGCGTACCATACCATCCGTC | |
| EFm1H90A-F | ATATGGTGCGACAGGTATCGCAACAGAGGATGG | pGEX-EFm1H90A |
| EFm1H90A-R | TACCTGTCGCACCATATTGAGCATAGTATCCTAAGCC | |
| EFm1D108A-F | CAGTGTTGCGCAAAACTGGATTAACCCAAGCC | pGEX-EFm1D108A |
| EFm1D108A-R | AGTTTTGCGCAACACTGACAAAGGTTCCGTCAG | |
| EFm1D20A-F | AAAGGCGTGGCGGCTGACGGATGGTATGGTACG | pGEX-EFm1D20A |
| EFm1D20A-R | TCAGCCGCCACGCCTTTTCCGACAAGG | |
| EFm1D22A-F | ACGCTGCGGGATGGTATGGTACGCAATGTATG | pGEX-EFm1D22A |
| EFm1D22A-R | ATACCATCCCGCAGCGTCCACGCCTTTTCC | |
| EFm1W24A-F | TGACGGAGCGTATGGTACGCAATGTATGGACTTGA | pGEX-EFm1W24A |
| EFm1W24A-R | TACCATACGCTCCGTCAGCGTCCACGCC | |
| EFm1G26A-F | TGACGGAGCGACGCAATGTATGGACTTGACAGTAG | pGEX-EFm1G26A |
| EFm1G26A-R | ATTGCGTCGCTCCGTCAGCGTCCACGCC | |
| EFm1D31A-F | GTACGCAATGTATGGCCTTGACAGT | pGEX-EFm1D31A |
| EFm1D31A-R | ACGTCTACTGTCAAGGCCATACATT | |
| EFm1M168A-F | GAGACGATGATATTGCGTTCATCTATTACAAACGCACTAAGCA | pGEX-EFm1M168A |
| EFm1M168A-R | CGCAATATCATCGTCTCCTTTAAATTGTCC | |
| EFm1M227A-F | TTGAAGGCGATGGAAGCAGCTTTACCACAAGT | pGEX-EFm1M227A |
| EFm1M227A-R | GCTTCCATCGCCTTCAATCCAAAGTTGTCGTGGT | |
| EFm1M227K-F | GAAGAAGATGGAAGCAGCTTTACCACAAGT | pGEX-EFm1M227K |
| EFm1M227K-R | CTGCTTCCATCTTCTTCAATCCAAAGTTGTCGTGGT | |
| EFm1K173E-F | CTATTACGCGCGCACTAAGCAAGGAAGCACT | pGEX-EFm1K173E |
| EFm1K173E-R | TAGTGCGCTCGTAATAGATGAACATAATATCATCGTCTCC | |
| EFm1K176E-F | ACGCACTGAGCAAGGAAGCACTGAGCAATGG | pGEX-EFm1K176E |
| EFm1K176E-R | TTCCTTGVTVAGTGCGTTTGTAATAGATGAACATAATATC | |
| EFm1R190E-F | GAGGTAAAGAGATCTACTTACCAACAATGACTTACGTAAAC | pGEX-EFm1R190E |
| EFm1R190E-R | GTAGATCTCTTTACCTCCAATAACGAACCATTG | |
| EFm1K236E-F | AGTTGAGGTATAGCTCGAGCGGCCGC | pGEX-EFm1K236E |
| EFm1K236E-R | CGAGCTATACCTCAACTTGTGGTAAAGCTGCTTCCA | |
| EFm1I186A-K189A-F | ACTGAGCAATGGTTCGTTCGTATCTACTTA | pGEX-EFm1I186A–K189A |
| EFm1I186A-K189A-R | TGTTGGTAAGTAGATACGAACGAACCATTG | |
| EFm1Y209A-F | GACCTTATCAAACGAGCTGGTGGAA | pGEX-EFm1Y209A |
| EFm1Y209A-R | GTTAGTGTTTCCACCAGCTCGTTTG | |
| EFm1R208A-F | GCATATGGTGGAAACACTAACGTAACGA | pGEX-EFm1R208A |
| EFm1R208A-R | GTGTTTCCACCATATGCTTTGATAAGGTCGTTAGCTTCGTT | |
| pDL278-EFm1-F | GAGCTCGGTACCCGGGGATCCCTATACTTTAACTTGTGGTAAA | pDL278-EFm1 |
| pDL278-EFm1-R | ACCATGATTACGCCAAGCTTAATGGTTAAATTAAACGATGTA | |
| dCas9-F | GACCATGATTACGCCAAGCTTAATGGACAAGAAGTACAGCATCGGC | pDL278-dCas9 |
| dCas9-R | GAGCTCGGTACCCGGGGATCCTTAGTCGCCGCCCAGCTG | |
| pDL278-F | ATGACCATGATTACGCCAAGCTTAATGGACAAGAA | Linearized pDL278-dCas9 |
| pDL278-R | TTGTTATCCGCTCACAATTCCACACAACATACGAG | |
| sgRNA-F | GAATTGTGAGCGGATAACAAGACGTTATGCAACGCTTCTTGTTTTAGAGCTAGAAATAGCAAG | pDL278-dCas9-sgRNA |
| sgRNA-R | CTTGGCGTAATCATGGTCATCAAAAAAAGCACCGACTCGG |
| Property | EFm1D166Q | CTD |
|---|---|---|
| Space group | I 41 2 2 | I 41 2 2 |
| Cell constants | 46.87 Å 57.26 Å 91.27 Å | 115.59 Å 115.59 Å 179.72 Å |
| a, b, c, α, β, γ | 90.00° 96.57° 90.00° | 90.00° 96.57° 90.00° |
| Resolution (Å) | 48.46–2.20 | 49.73–2.00 |
| 48.42–2.20 | 49.68–2.00 | |
| % Data completeness | 99.9 (48.46–2.20) | 99.0 (49.73–2.00) |
| (in resolution range) | 98.2 (48.42–2.20) | 98.2 (49.68–2.00) |
| Rmerge | 0.00 | 0.00 |
| < I/σ (I) > | 2.75 (at 2.20 Å) | 1.39 (at 2.00 Å) |
| Refinement program | REFMAC 5.8.0267, REFMAC 5.8.0267 | REFMAC 5.8.0267, REFMAC 5.8.0267 |
| R, Rfree | 0.179, 0.215 | 0.183, 0.216 |
| 0.197, 0.223 | 0.177, 0.214 | |
| Rfree test set | 1295 reflections (5.27%) | 1998 reflections (4.91%) |
| Wilson B-factor (Å) | 27.1 | 21.8 |
| Anisotropy | 0.681 | 0.121 |
| Bulk solvent ksol (e/Å), Bsol (Å) | 0.39, 33.6 | 0.42, 58.0 |
| L-test for twinning | < |L| > = 0.49, < L2 > = 0.32 | < |L| > = 0.49, < L2 > = 0.32 |
| Estimated twinning fraction | No twinning to report. | No twinning to report. |
| FO, FC correlation | 0.94 | 0.96 |
| Total number of atoms | 6953 | 9609 |
| Average B, all atoms (Å) | 24.0 | 23.0 |
| Structural Characteristics | Mutants | Stability | Lytic Activity |
|---|---|---|---|
| Critical domains | NTD (1–165 aa) | − | − |
| CTD (165–237 aa) | − | − | |
| EFm1 lacking the linker (Δ145–165 aa) | − | − | |
| Putative residues binding to the host cell | K173E | + | + |
| K176E | + | + | |
| R190E | + | − | |
| K236E | + | + | |
| Residues binding calcium | D20A | + | − |
| D22A | + | − | |
| W24A | + | + | |
| G26A | + | + | |
| D31A | + | − | |
| Residues forming a putative catalytic triad | C29A | + | − |
| H90A | + | − | |
| D108A | + | − | |
| Residues producing and assembling the 8 kDa fragment | M168A | − | − |
| M227K | − | − | |
| Residues forming the interaction surface | R208A | + | − |
| Y209A | + | − | |
| Residues forming the putative ion channel | I186A-K189A | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Zeng, X.; Wang, L.; Zheng, Y.; Zhang, G.; Cheng, W. The Structure and Function of Biomaterial Endolysin EFm1 from E. faecalis Phage. Materials 2022, 15, 4879. https://doi.org/10.3390/ma15144879
Zhou X, Zeng X, Wang L, Zheng Y, Zhang G, Cheng W. The Structure and Function of Biomaterial Endolysin EFm1 from E. faecalis Phage. Materials. 2022; 15(14):4879. https://doi.org/10.3390/ma15144879
Chicago/Turabian StyleZhou, Xuerong, Xiaotao Zeng, Li Wang, Yanhui Zheng, Guixiang Zhang, and Wei Cheng. 2022. "The Structure and Function of Biomaterial Endolysin EFm1 from E. faecalis Phage" Materials 15, no. 14: 4879. https://doi.org/10.3390/ma15144879