
Biochar and Hyperthermophiles as Additives Accelerate the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements during Composting

Abstract
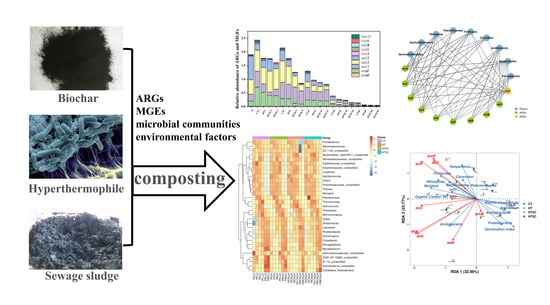
:
1. Introduction
2. Materials and Methods
2.1. Materials and Experiment Setup
2.2. Sample Collection
2.3. Real-Time Quantitative PCR (qPCR)
2.4. 16S rRNA Gene High-Throughput Sequencing
2.5. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Changes during Composting
3.2. Variation of ARGs and an MGE during Composting
3.3. Evolution of the Soil Microbial Community
3.4. Relationships between the Bacterial Community, ARGs and the MGE
3.5. Conditions Affecting the Change of ARGs in Composting
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Z.; Sun, X. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar. Bioresour. Technol. 2014, 171, 274–284. [Google Scholar]
- Kalashnikov, M.; Mueller, M.; Mcbeth, C.; Lee, J.C.; Campbell, J.; Sharon, A.; Sauer-Budge, A.F. Rapid phenotypic stress-based microfluidic antibiotic susceptibility testing of Gram-negative clinical isolates. Sci. Rep. 2017, 7, 8031. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Hu, X.; Zhang, Y.; Jiang, W.; Zhang, X. Co/La modified Ti/PbO 2 anodes for chloramphenicol degradation: Catalytic performance and reaction mechanism. Chemosphere 2021, 285, 131568. [Google Scholar] [CrossRef]
- Luo, Y.; Mao, D.; Rysz, M.; Zhou, Q.; Alvarez, P. Trends in Antibiotic Resistance Genes Occurrence in the Haihe River, China. Environ. Sci. Technol. 2010, 44, 7220–7225. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, H.; Ma, J.; Sun, W.; Yang, Y.; Zhang, X. Compost-bulking agents reduce the reservoir of antibiotics and antibiotic resistance genes in manures by modifying bacterial microbiota. Sci. Total Environ. 2019, 649, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cui, Y.; Zhang, W.; Wang, C.; Li, A. Fate of antibiotics and the related antibiotic resistance genes during sludge stabilization in sludge treatment wetlands. Chemosphere 2019, 224, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Burch, T.R.; Sadowsky, M.J.; LaPara, T.M. Fate of Antibiotic Resistance Genes and Class 1 Integrons in Soil Microcosms Following the Application of Treated Residual Municipal Wastewater Solids. Environ. Sci. Technol. 2014, 48, 5620–5627. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.Y.; Shi, P.; Hu, Q.; Li, B.; Zhang, T.; Zhang, X.X. Bacterial Community Shift Drives Antibiotic Resistance Promotion during Drinking Water Chlorination. Environ. Sci. Technol. 2015, 49, 12271–12279. [Google Scholar] [CrossRef]
- Cheng, D.M.; Feng, Y.; Liu, Y.W.; Xue, J.M.; Li, Z.J. Dynamics of oxytetracycline, sulfamerazine, and ciprofloxacin and related antibiotic resistance genes during swine manure composting. J. Environ. Manag. 2019, 230, 102–109. [Google Scholar] [CrossRef]
- Xie, W.Y.; Yang, X.P.; Li, Q.; Wu, L.H.; Shen, Q.R.; Zhao, F.J. Changes in antibiotic concentrations and antibiotic resistome during commercial composting of animal manures. Environ. Pollut. 2016, 219, 182–190. [Google Scholar] [CrossRef]
- Ray, P.; Chen, C.; Knowlton, K.F.; Pruden, A.; Xia, K. Fate and Effect of Antibiotics in Beef and Dairy Manure during Static and Turned Composting. J. Environ. Qual. 2017, 46, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Q.; Wang, Y.; Wen, Q.X. Effects of chlortetracycline on the fate of multi-antibiotic resistance genes and the microbial community during swine manure composting. Environ. Pollut. 2018, 237, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ben, W.W.; Zhang, Y.; Yang, M.; Qiang, Z.M. Effects of thermophilic composting on oxytetracycline, sulfamethazine, and their corresponding resistance genes in swine manure. Environ. Sci. Process. Impacts 2015, 17, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Chen, M.X.; Sui, Q.W.; Tong, J.; Jiang, C.; Lu, X.T.; Zhang, Y.X.; Wei, Y.S. Impacts of addition of natural zeolite or a nitrification inhibitor on antibiotic resistance genes during sludge composting. Water Res. 2016, 91, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Gu, J.; Wang, X.; Yu, J.; Ma, J. Responses of antibiotic and heavy metal resistance genes to bamboo charcoal and bamboo vinegar during aerobic composting. Environ. Pollut. 2019, 252, 1097–1105. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Duan, Y.; Liu, T.; Awasthi, S.K.; Zhang, Z. Relevance of biochar to influence the bacterial succession during pig manure composting. Bioresour. Technol. 2020, 304, 122962. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Ducey, T.F.; Cantrell, K.B.; Novak, J.M.; Lentz, R.D. Designer, acidic biochar influences calcareous soil characteristics. Chemosphere 2016, 142, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Su, J.Q.; Wei, B.; Ou-Yang, W.Y.; Huang, F.Y.; Zhao, Y.; Xu, H.J.; Zhu, Y.G. Antibiotic Resistome and Its Association with Bacterial Communities during Sewage Sludge Composting. Environ. Sci. Technol. 2015, 49, 7356–7363. [Google Scholar] [CrossRef]
- He, G.; Jiang, X.; Yao, L.; Liu, G.; Liu, W. Effects of tetracycline on nitrogen and carbon cycling rates and microbial abundance in sediments with and without biochar amendment. Chemosphere 2020, 270, 129509. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, J.A.; Stromberger, M.E.; Lentz, R.D.; Dungan, R.S. Hardwood biochar and manure co-application to a calcareous soil. Chemosphere 2016, 142, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Lu, X.; Rensing, C.; Friman, V.P.; Geisen, S.; Zhi, C.; Zhen, Y.; Zhong, W.; Zhou, S.; Zhu, Y. Hyperthermophilic Composting Accelerates the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements in Sewage Sludge. Environ. Sci. Technol. 2018, 52, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Mao, Q.; Su, Y.A.; Zhang, H.; Wen, D. Thermophilic rather than mesophilic sludge anaerobic digesters possess lower antibiotic resistant genes abundance. Bioresour. Technol. 2021, 329, 124924. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Jia, T.; Liao, H.; Liu, X.; Zhou, P.; Zhi, C.; Christopher, R.; Zhou, S. The distinctive microbial community improves composting efficiency in a full-scale hyperthermophilic composting plant. Bioresour. Technol. 2018, 265, 146–154. [Google Scholar]
- Cheng, W.; Lu, H.; Dong, D.; Deng, H.; Strong, P.J.; Wang, H.; Wu, W. Insight into the Effects of Biochar on Manure Composting: Evidence Supporting the Relationship between N2O Emission and Denitrifying Community. Environ. Sci. Technol. 2013, 47, 7341–7349. [Google Scholar]
- Chen, Y.X.; Huang, X.D.; Han, Z.Y.; Huang, X.; Hu, B.; Shi, D.Z.; Wu, W.X. Effects of bamboo charcoal and bamboo vinegar on nitrogen conservation and heavy metals immobility during pig manure composting. Chemosphere 2010, 78, 1177–1181. [Google Scholar] [CrossRef]
- Steiner, C.; Das, K.C.; Melear, N.; Lakly, D. Reducing Nitrogen Loss during Poultry Litter Composting Using Biochar. J. Environ. Qual. 2010, 39, 1236–1242. [Google Scholar] [CrossRef] [Green Version]
- Rasapoor, M.; Nasrabadi, T.; Kamali, M.; Hoveidi, H. The effects of aeration rate on generated compost quality, using aerated static pile method. Waste Manag. 2009, 29, 570–573. [Google Scholar] [CrossRef]
- Gomez, D.K.V.; Castaneda, P.B.; Rosales, J.D.M.; Rodriguez, L.M.G. Anaerobic digestion of bean straw applying a fungal pre-treatment and using cow manure as co-substrate. Environ. Technol. 2020, 41, 2863–2874. [Google Scholar] [CrossRef]
- Zhao, J.C.; Sun, X.N.; Awasthi, M.K.; Wang, Q.; Ren, X.N.; Li, R.H.; Chen, H.Y.; Wang, M.J.; Liu, T.; Zhang, Z.Q. Performance evaluation of gaseous emissions and Zn speciation during Zn-rich antibiotic manufacturing wastes and pig manure composting. Bioresour. Technol. 2018, 267, 688–695. [Google Scholar] [CrossRef]
- Gillings, M.R. Class 1 integrons as invasive species. Curr. Opin. Microbiol. 2017, 38, 10–15. [Google Scholar] [CrossRef]
- Li, H.C.; Duan, M.L.; Gu, J.; Zhang, Y.J.; Qian, X.; Ma, J.; Zhang, R.R.; Wang, X.J. Effects of bamboo charcoal on antibiotic resistance genes during chicken manure composting. Ecotoxicol. Environ. Saf. 2017, 140, 1–6. [Google Scholar] [CrossRef]
- Guan, J.; Wasty, A.; Grenier, C.; Chan, M. Influence of temperature on survival and conjugative transfer of multiple antibiotic-resistant plasmids in chicken manure and compost microcosms. Poult. Sci. 2007, 86, 610–613. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Sun, W.; Gu, J.; Wang, X.-J.; Zhang, Y.-J.; Duan, M.-L.; Li, H.-C.; Zhang, R.-R. Reducing antibiotic resistance genes, integrons, and pathogens in dairy manure by continuous thermophilic composting. Bioresour. Technol. 2016, 220, 425–432. [Google Scholar] [CrossRef]
- Liao, H.; Zhao, Q.; Cui, P.; Chen, Z.; Yu, Z.; Geisen, S.; Friman, V.-P.; Zhou, S. Efficient reduction of antibiotic residues and associated resistance genes in tylosin antibiotic fermentation waste using hyperthermophilic composting. Environ. Int. 2019, 133, 105203. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.M.; Changxiong, Z.; Bing, G.; Raza, A.H.; Tingting, S.; Hongna, L. The fate of antibiotic resistance genes in cow manure composting: Shaped by temperature-controlled composting stages. Bioresour. Technol. 2021, 320, 124403. [Google Scholar]
- Honghong, G.; Shuhong, X.; Mubasher, N.; Jialong, L.; Jei, G. Role of Bentonite on the Mobility of Antibiotic Resistance Genes, and Microbial Community in Oxytetracycline and Cadmium Contaminated Soil. Front. Microbiol. 2018, 9, 2722. [Google Scholar]
- Qiu, X.; Zhou, G.; Zhang, J.; Wang, W. Microbial community responses to biochar addition when a green waste and manure mix are composted: A molecular ecological network analysis. Bioresour. Technol. 2019, 273, 666–671. [Google Scholar] [CrossRef]
- Huerta, B.; Marti, E.; Gros, M.; Lopez, P.; Pompeo, M.; Armengol, J.; Barcelo, D.; Balcazar, J.L.; Rodriguez-Mozaz, S.; Marce, R. Exploring the links between antibiotic occurrence, antibiotic resistance, and bacterial communities in water supply reservoirs. Sci. Total Environ. 2013, 456, 161–170. [Google Scholar] [CrossRef]
- Jzab, C.; Qsab, C.; Tl, D.; Hui, Z.; Ps, D.; Ywab, C. Sludge bio-drying followed by land application could control the spread of antibiotic resistance genes—ScienceDirect. Environ. Int. 2019, 130, 104906. [Google Scholar]
- Zhang, R.; Gu, J.; Wang, X.; Li, Y.; Zhang, K.; Yin, Y.; Zhang, X. Contributions of the microbial community and environmental variables to antibiotic resistance genes during co-composting with swine manure and cotton stalks. J. Hazard. Mater. 2018, 358, 82–91. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fierer, N.; Dantas, G. Bacterial phylogeny structures soil resistomes across habitats. Nature 2014, 509, 612. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.L.; An, X.L.; Li, H.; Su, J.Q.; Ma, Y.B.; Zhu, Y.G. Long-term field application of sewage sludge increases the abundance of antibiotic resistance genes in soil. Environ. Int. 2016, 92–93, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Huang, X.H.; Sun, J.Z.; Graham, D.W.; Xie, B. Antibiotic Resistance Genes and Associated Microbial Community Conditions in Aging Landfill Systems. Environ. Sci. Technol. 2017, 51, 12859–12867. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.-W.; Wang, J.-T.; Li, J.; Shi, X.-Z.; Ma, Y.-B.; Chen, D.; He, J.-Z. Long-Term Nickel Contamination Increases the Occurrence of Antibiotic Resistance Genes in Agricultural Soils. Environ. Sci. Technol. 2017, 51, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sangwan, N.; Li, H.Y.; Su, J.Q.; Oyang, W.Y.; Zhang, Z.J.; Gilbert, J.A.; Zhu, Y.G.; Ping, F.; Zhang, H.L. The antibiotic resistome of swine manure is significantly altered by association with the Musca domestica larvae gut microbiome. ISME J. 2017, 11, 100–111. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; Mcarthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | ermB | ermF | sul1 | sul2 | sul3 | tetA | tetG | tetM | tetO | IntI1 |
|---|---|---|---|---|---|---|---|---|---|---|
| CT | 13.8% a | 96.2% c | 87.5% c | 93.0% c | 96.0% b | 59.3% d | 88.7% c | -- | 23.1% d | 96.9% c |
| HT | 14.7% a | 96.6% b,c | 95.5% a | 97.5% b | 98.5% a | 77.2% c | 88.2% d | 42.6% | 60.0% c | 97.7% b |
| HT2C | 11.0% b | 97.9% a,b | 94.8% b | 99.3% a,b | 98.4% a | 86.6% b | 93.4% a | -- | 77.9% b | 99.3% a |
| HT5C | 7.9% c | 98.5% a | 95.3% a | 99.5% a | 98.8% a | 92.2% a | 91.6% b | -- | 80.1% a | 99.3% a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Zhang, A.; Guo, T.; Zhu, Y.; Shao, Y. Biochar and Hyperthermophiles as Additives Accelerate the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements during Composting. Materials 2021, 14, 5428. https://doi.org/10.3390/ma14185428
Fu Y, Zhang A, Guo T, Zhu Y, Shao Y. Biochar and Hyperthermophiles as Additives Accelerate the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements during Composting. Materials. 2021; 14(18):5428. https://doi.org/10.3390/ma14185428
Chicago/Turabian StyleFu, Yanli, Aisheng Zhang, Tengfei Guo, Ying Zhu, and Yanqiu Shao. 2021. "Biochar and Hyperthermophiles as Additives Accelerate the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements during Composting" Materials 14, no. 18: 5428. https://doi.org/10.3390/ma14185428