
Characterization of the Ensemble of Lignin-Remodeling DyP-Type Peroxidases from Streptomyces coelicolor A3(2)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Cloning
2.2. Overexpression
2.3. Purification
2.4. Determination of Enzyme Concentration
2.5. Enzyme Assays
2.6. Organosolv Extraction of Lignin
2.7. Enzymatic Treatment of Organosolv Lignins
3. Results
3.1. The Ensemble of DyP-Peroxidases of Streptomyces coelicolor
3.2. Production and Purification of Recombinant ScDyPs
3.3. Biochemical Characterization
3.3.1. Optimal pH for ScDyPs
3.3.2. Temperature Dependency
3.3.3. Thermostability
3.3.4. Steady-State Kinetics
3.4. Lignin Remodeling Activity of ScDyPs
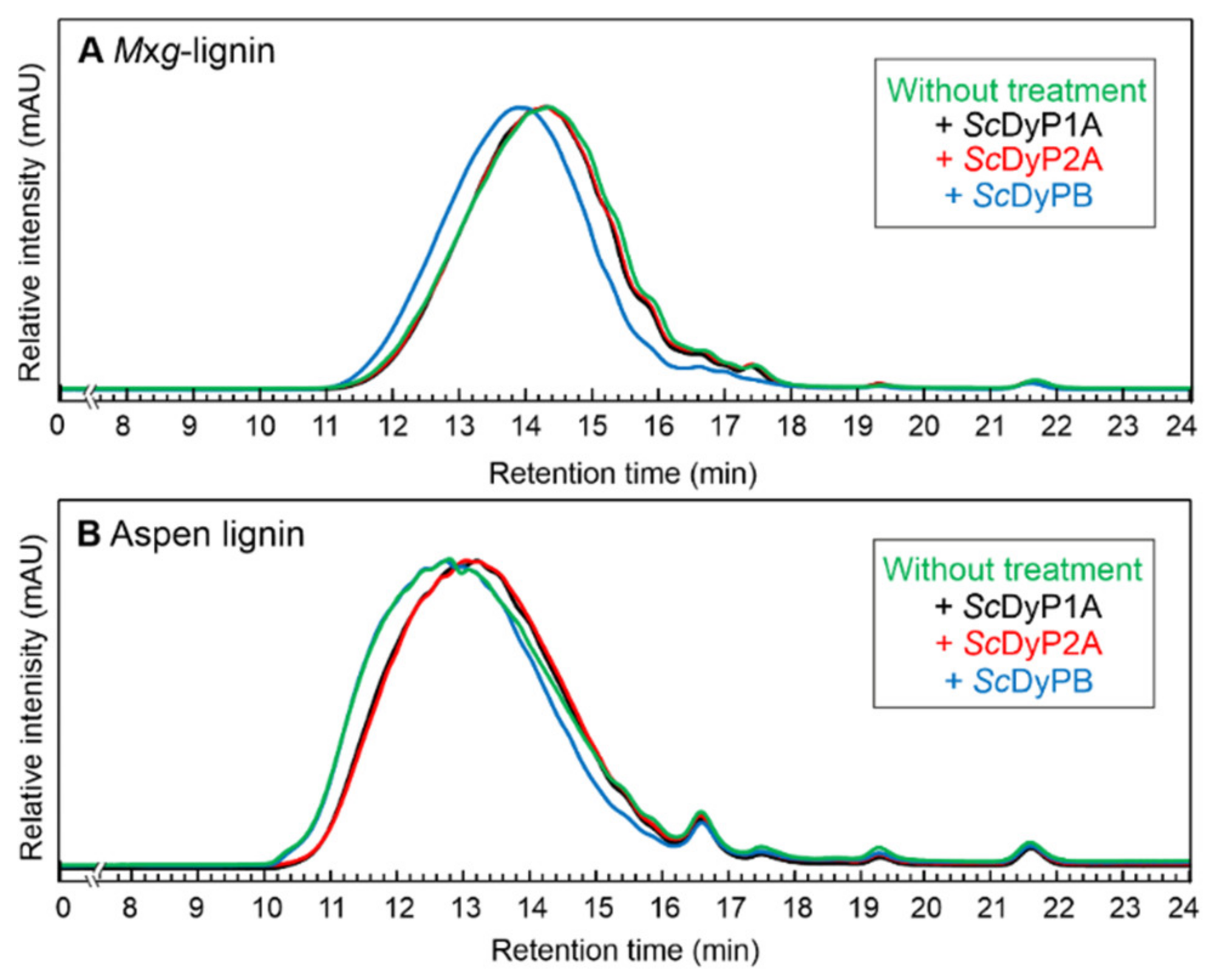
3.4.1. Identification of Molecular Weight Distribution of Lignin Samples by GPC Analysis
3.4.2. Identification of Low-Molecular-Weight Compounds in Lignin Samples by GC-MS Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dahmen, N.; Lewandowski, I.; Zibek, S.; Weidtmann, A. Integrated Lignocellulosic Value Chains in a Growing Bioeconomy: Status Quo and Perspectives. GCB Bioenergy 2019, 11, 107–117. [Google Scholar] [CrossRef]
- Watkins, D.; Nuruddin, M.; Hosur, M.; Tcherbi-Narteh, A.; Jeelani, S. Extraction and Characterization of Lignin from Different Biomass Resources. J. Mater. Res. Technol. 2015, 4, 26–32. [Google Scholar] [CrossRef]
- Chandel, A.K.; Garlapati, V.K.; Singh, A.K.; Antunes, F.A.F.; da Silva, S.S. The Path Forward for Lignocellulose Biorefineries: Bottlenecks, Solutions, and Perspective on Commercialization. Bioresour. Technol. 2018, 264, 370–381. [Google Scholar] [CrossRef]
- Cao, L.; Yu, I.K.M.; Liu, Y.; Ruan, X.; Tsang, D.C.W.; Hunt, A.J. Lignin Valorization for the Production of Renewable Chemicals : State-of-the- Art Review and Future Prospects. Bioresour. Technol. 2018, 269, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Anushree; Kumar, J.; Bhaskar, T. Utilization of Lignin: A Sustainable and Eco-Friendly Approach. J. Energy Inst. 2020, 93, 235–271. [Google Scholar] [CrossRef]
- Ragauskas, A.J.; Beckham, G.T.; Biddy, M.J.; Chandra, R.; Chen, F.; Davis, M.F.; Davison, B.H.; Dixon, R.A.; Gilna, P.; Keller, M.; et al. Lignin Valorization : Improving Lignin Processing in the Biorefinery. Science 2014, 344, 1246843. [Google Scholar] [CrossRef]
- Li, C.; Zhao, X.; Wang, A.; Huber, G.W.; Zhang, T. Catalytic Transformation of Lignin for the Production of Chemicals and Fuels. Chem. Rev. 2015, 115, 11559–11624. [Google Scholar] [CrossRef]
- Schutyser, W.; Renders, T.; Van Den Bosch, S.; Koelewijn, S.F.; Beckham, G.T.; Sels, B.F. Chemicals from Lignin: An Interplay of Lignocellulose Fractionation, Depolymerisation, and Upgrading. Chem. Soc. Rev. 2018, 47, 852–908. [Google Scholar]
- Becker, J.; Wittmann, C. A Field of Dreams: Lignin Valorization into Chemicals, Materials, Fuels, and Health-Care Products. Biotechnol. Adv. 2019, 37, 107360. [Google Scholar]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Chio, C.; Sain, M.; Qin, W. Lignin Utilization : A Review of Lignin Depolymerization from Various Aspects. Renew. Sustain. Energy Rev. 2019, 107, 232–249. [Google Scholar] [CrossRef]
- Koranyi, T.I.; Fridrich, B.; Pineda, A.; Barta, K. Development of “Lignin-First” Approaches for the Valorization of Lignocellulosic Biomass. Molecules 2020, 25, 2815. [Google Scholar] [CrossRef] [PubMed]
- Dessbesell, L.; Paleologou, M.; Leitch, M.; Pulkki, R.; Xu, C. Global Lignin Supply Overview and Kraft Lignin Potential as an Alternative for Petroleum-Based Polymers. Renew. Sustain. Energy Rev. 2020, 123, 109768. [Google Scholar] [CrossRef]
- Bajwa, D.S.; Pourhashem, G.; Ullah, A.H.; Bajwa, S.G. A Concise Review of Current Lignin Production, Applications, Products and Their Environment Impact. Ind. Crops Prod. 2019, 139, 111526. [Google Scholar] [CrossRef]
- Jõul, P.; Ho, T.T.; Kallavus, U.; Konist, A.; Leiman, K.; Salm, O.S.; Kulp, M.; Koel, M.; Lukk, T. Characterization of Organosolv Lignins and Their Application in the Preparation of Aerogels. Materials 2022, 15, 2861. [Google Scholar] [CrossRef]
- Bauer, S.; Sorek, H.; Mitchell, V.D.; Ibáñez, A.B.; Wemmer, D.E. Characterization of Miscanthus Giganteus Lignin Isolated by Ethanol Organosolv Process under Reflux Condition. J. Agric. Food Chem. 2012, 60, 8203–8212. [Google Scholar] [CrossRef]
- Tribot, A.; Amer, G.; Abdou Alio, M.; de Baynast, H.; Delattre, C.; Pons, A.; Mathias, J.D.; Callois, J.M.; Vial, C.; Michaud, P.; et al. Wood-Lignin: Supply, Extraction Processes and Use as Bio-Based Material. Eur. Polym. J. 2019, 112, 228–240. [Google Scholar] [CrossRef]
- Chan, J.C.; Paice, M.; Zhang, X. Enzymatic Oxidation of Lignin: Challenges and Barriers Toward Practical Applications. ChemCatChem 2020, 12, 401–425. [Google Scholar] [CrossRef]
- Fache, M.; Boutevin, B.; Caillol, S. Vanillin Production from Lignin and Its Use as a Renewable Chemical. ACS Sustain. Chem. Eng. 2016, 4, 35–46. [Google Scholar] [CrossRef]
- Van Den Bosch, S.; Renders, T.; Kennis, S.; Koelewijn, S.F.; Van Den Bossche, G.; Vangeel, T.; Deneyer, A.; Depuydt, D.; Courtin, C.M.; Thevelein, J.M.; et al. Integrating Lignin Valorization and Bio-Ethanol Production: On the Role of Ni-Al2O3 Catalyst Pellets during Lignin-First Fractionation. Green Chem. 2017, 19, 3313–3326. [Google Scholar] [CrossRef]
- Shuai, L.; Talebi Amiri, M.; Questell-Santiago, Y.M.; Héroguel, F.; Li, Y.; Kim, H.; Meilan, R.; Chapple, C.; Ralph, J.; Luterbacher, J.S. Formaldehyde Stabilization Facilitates Lignin Monomer Production during Biomass Depolymerization. Science 2016, 354, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Rahmanpour, R. Pathways for Degradation of Lignin in Bacteria and Fungi. Nat. Prod. Rep. 2011, 28, 1883–1896. [Google Scholar] [CrossRef]
- Tian, J.; Pourcher, A.; Bouchez, T.; Gelhaye, E.; Peu, P. Occurrence of Lignin Degradation Genotypes and Phenotypes among Prokaryotes. Appl. Microbiol. Biotechnol. 2014, 98, 9527–9544. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Lukk, T.; Solbiati, J.O.; Bauer, S.; Nair, S.K.; Cronan, J.E.; Gerlt, J.A. Roles of Small Laccases from Streptomyces in Lignin Degradation. Biochemistry 2014, 53, 4047–4058. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkolazka, A.; Paszczyński, A. Lignin Degradation: Microorganisms, Enzymes Involved, Genomes Analysis and Evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, C.; Wu, X.; Tsang, C.; Mou, J.; Yan, J.; Liu, Y.; Sze, C.; Lin, K. Recent Advancement in Lignin Biorefinery: With Special Focus on Enzymatic Degradation and Valorization. Bioresour. Technol. 2019, 291, 121898. [Google Scholar] [CrossRef] [PubMed]
- Zámocky, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent Evolution of Four Heme Peroxidase Superfamilies. Arch. Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.U.N.; Shoda, M. Purification and Characterization of a Novel Peroxidase from Geotrichum Candidum Dec 1 Involved in Decolorization of Dyes. Appl. Environ. Microbiol. 1999, 65, 1029–1035. [Google Scholar] [CrossRef]
- Sugano, Y.; Muramatsu, R.; Ichiyanagi, A.; Sato, T.; Shoda, M. DyP, a Unique Dye-Decolorizing Peroxidase, Represents a Novel Heme Peroxidase Family: ASP171 Replaces the Distal Histidine of Classical Peroxidases. J. Biol. Chem. 2007, 282, 36652–36658. [Google Scholar] [CrossRef]
- Kim, S.J.; Ishikawa, K.; Hirai, M.; Shoda, M. Characteristics of a Newly Isolated Fungus, Geotrichum Candidum Dec 1, Which Decolorizes Various Dyes. J. Ferment. Bioeng. 1995, 79, 601–607. [Google Scholar] [CrossRef]
- Singh, R.; Eltis, L.D. The Multihued Palette of Dye-Decolorizing Peroxidases. Arch. Biochem. Biophys. 2015, 574, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Nishihashi, Y.; Narioka, T.; Yoshida, T.; Morita, M.; Sugano, Y. Characterization of a Novel DyP-Type Peroxidase from Streptomyces Avermitilis. J. Biosci. Bioeng. 2017, 123, 425–430. [Google Scholar] [CrossRef]
- Ahmad, M.; Roberts, J.N.; Hardiman, E.M.; Singh, R.; Eltis, L.D.; Bugg, T.D.H. Identification of DypB from Rhodococcus Jostii RHA1 as a Lignin Peroxidase. Biochemistry 2011, 50, 5096–5107. [Google Scholar] [CrossRef] [PubMed]
- Liers, C.; Bobeth, C.; Pecyna, M.; Ullrich, R.; Hofrichter, M. DyP-like Peroxidases of the Jelly Fungus Auricularia Auricula-Judae Oxidize Nonphenolic Lignin Model Compounds and High-Redox Potential Dyes. Appl. Microbiol. Biotechnol. 2010, 85, 1869–1879. [Google Scholar] [CrossRef]
- Min, K.; Gong, G.; Woo, H.M.; Kim, Y.; Um, Y. A Dye-Decolorizing Peroxidase from Bacillus Subtilis Exhibiting Substrate-Dependent Optimum Temperature for Dyes and β-Ether Lignin Dimer. Sci. Rep. 2015, 5, 8245. [Google Scholar] [CrossRef] [PubMed]
- Fawal, N.; Li, Q.; Savelli, B.; Brette, M.; Passaia, G.; Fabre, M.; Mathé, C.; Dunand, C. PeroxiBase: A Database for Large-Scale Evolutionary Analysis of Peroxidases. Nucleic Acids Res. 2013, 41, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Sugano, Y. A Structural and Functional Perspective of DyP-Type Peroxidase Family. Arch. Biochem. Biophys. 2015, 574, 49–55. [Google Scholar] [CrossRef]
- Colpa, D.I.; Fraaije, M.W.; Bloois, E. Van. DyP-Type Peroxidases: A Promising and Versatile Class of Enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1–7. [Google Scholar] [CrossRef]
- Salvachúa, D.; Prieto, A.; Martínez, Á.T.; Martínez, M.J. Enzyme from Irpex Lacteus and Its Application in Enzymatic Hydrolysis of Wheat Straw. Appl. Environ. Microbiol. 2013, 79, 4316–4324. [Google Scholar] [CrossRef]
- Roberts, J.N.; Singh, R.; Grigg, J.C.; Murphy, M.E.P.; Bugg, T.D.H.; Eltis, L.D. Characterization of Dye-Decolorizing Peroxidases from Rhodococcus Jostii RHA1. Biochemistry 2011, 50, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Van Bloois, E.; Pazmiño, D.E.T.; Winter, R.T.; Fraaije, M.W. A Robust and Extracellular Heme-Containing Peroxidase from Thermobifida Fusca as Prototype of a Bacterial Peroxidase Superfamily. Appl. Microbiol. Biotechnol. 2010, 86, 1419–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmanpour, R.; Bugg, T.D.H. Characterisation of Dyp-Type Peroxidases from Pseudomonas Fluorescens Pf-5: Oxidation of Mn(II) and Polymeric Lignin by Dyp1B. Arch. Biochem. Biophys. 2015, 574, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete Genome Sequence of the Model Actinomycete Streptomyces Coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Teder, T.; Lõhelaid, H.; Samel, N. Structural and Functional Insights into the Reaction Specificity of Catalase-Related Hydroperoxide Lyase: A Shift from Lyase Activity to Allene Oxide Synthase by Site-Directed Mutagenesis. PLoS ONE 2017, 12, e0185291. [Google Scholar] [CrossRef]
- Breslmayr, E.; Hanžek, M.; Hanrahan, A.; Leitner, C.; Kittl, R.; Šantek, B.; Oostenbrink, C.; Ludwig, R. A Fast and Sensitive Activity Assay for Lytic Polysaccharide Monooxygenase. Biotechnol. Biofuels 2018, 11, 79. [Google Scholar] [CrossRef]
- Ollinger, J.; Song, K.B.; Antelmann, H.; Hecker, M.; Helmann, J.D. Role of the Fur Regulon in Iron Transport in Bacillus Subtilis. J. Bacteriol. 2006, 188, 3664–3673. [Google Scholar] [CrossRef] [PubMed]
- Biswas, L.; Biswas, R.; Nerz, C.; Ohlsen, K.; Schlag, M.; Schäfer, T.; Lamkemeyer, T.; Ziebandt, A.K.; Hantke, K.; Rosenstein, R.; et al. Role of the Twin-Arginine Translocation Pathway in Staphylococcus. J. Bacteriol. 2009, 191, 5921–5929. [Google Scholar] [CrossRef]
- Létoffé, S.; Heuck, G.; Delepelaire, P.; Lange, N.; Wandersman, C. Bacteria Capture Iron from Heme by Keeping Tetrapyrrol Skeleton Intact. Proc. Natl. Acad. Sci. USA 2009, 106, 11719–11724. [Google Scholar] [CrossRef]
- Zitare, U.A.; Habib, M.H.; Rozeboom, H.; Mascotti, M.L.; Todorovic, S.; Fraaije, M.W. Mutational and Structural Analysis of an Ancestral Fungal Dye-Decolorizing Peroxidase. FEBS J. 2021, 288, 3602–3618. [Google Scholar] [CrossRef]
- Jeong, H.; Barbe, V.; Lee, C.H.; Vallenet, D.; Yu, D.S.; Choi, S.H.; Couloux, A.; Lee, S.W.; Yoon, S.H.; Cattolico, L.; et al. Genome Sequences of Escherichia Coli B Strains REL606 and BL21(DE3). J. Mol. Biol. 2009, 394, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant Protein Expression in Escherichia Coli: Advances and Challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosano, G.L.; Morales, E.S.; Ceccarelli, E.A. New Tools for Recombinant Protein Production in Escherichia Coli: A 5-Year Update. Protein Sci. 2019, 28, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Lithwick, G.; Margalit, H. Hierarchy of Sequence-Dependent Features Associated with Prokaryotic Translation. Genome Res. 2003, 13, 2665–2673. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, C.; Govindarajan, S.; Minshull, J. Codon Bias and Heterologous Protein Expression. Trends Biotechnol. 2004, 22, 346–353. [Google Scholar] [CrossRef]
- Al-Hawash, A.B.; Zhang, X.; Ma, F. Strategies of Codon Optimization for High-Level Heterologous Protein Expression in Microbial Expression Systems. Gene Rep. 2017, 9, 46–53. [Google Scholar] [CrossRef]
- Johansson, A.S.; Bolton-Grob, R.; Mannervik, B. Use of Silent Mutations in CDNA Encoding Human Glutathione Transferase M2-2 for Optimized Expression in Escherichia Coli. Protein Expr. Purif. 1999, 17, 105–112. [Google Scholar] [CrossRef]
- Jung, Y.; Kwak, J.; Lee, Y. High-Level Production of Heme-Containing Holoproteins in Escherichia Coli. Appl. Microbiol. Biotechnol. 2001, 55, 187–191. [Google Scholar] [CrossRef]
- Chouchane, S.; Lippai, I.; Magliozzo, R.S. Catalase-Peroxidase (Mycobacterium Tuberculosis KatG) Catalysis and Isoniazid Activation. Biochemistry 2000, 39, 9975–9983. [Google Scholar] [CrossRef]
- Sudhamsu, J.; Kabir, M.; Airola, M.V.; Patel, B.A.; Yeh, S.R.; Rousseau, D.L.; Crane, B.R. Co-Expression of Ferrochelatase Allows for Complete Heme Incorporation into Recombinant Proteins Produced in E. Coli. Protein Expr. Purif. 2010, 73, 78–82. [Google Scholar] [CrossRef]
- Varnado, C.L.; Goodwin, D.C. System for the Expression of Recombinant Hemoproteins in Escherichia Coli. Protein Expr. Purif. 2004, 35, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Sugano, Y.; Nakano, R.; Sasaki, K.; Shoda, M. Efficient Heterologous Expression in Aspergillus Oryzae of a Unique Dye- Decolorizing Peroxidase, DyP, of Geotrichum Candidum Dec 1. Appl. Environ. Microbiol. 2000, 66, 1754–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, K.; Yoshida, T.; Hirashima, R.; Toriumi, R.; Akiyama, H.; Kakuta, Y.; Ishige, Y.; Sugano, Y. Characterization of Class V DyP-Type Peroxidase SaDyP1 from Streptomyces Avermitilis and Evaluation of SaDyPs Expression in Mycelium. Int. J. Mol. Sci. 2021, 22, 8683. [Google Scholar] [CrossRef] [PubMed]
- Linde, D.; Ayuso-Fernández, I.; Laloux, M.; Aguiar-Cervera, J.E.; de Lacey, A.L.; Ruiz-Dueñas, F.J.; Martínez, A.T. Comparing Ligninolytic Capabilities of Bacterial and Fungal Dye-Decolorizing Peroxidases and Class-Ii Peroxidase-Catalases. Int. J. Mol. Sci. 2021, 22, 2629. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Borges, P.T.; Scocozza, M.F.; Silva, D.; Taborda, A.; Brissos, V.; Frazão, C.; Martins, L.O. Loops around the Heme Pocket Have a Critical Role in the Function and Stability of Bsdyp from Bacillus Subtilis. Int. J. Mol. Sci. 2021, 22, 10862. [Google Scholar] [CrossRef]
- Chaplin, A.K.; Wilson, M.T.; Worrall, J.A.R. Kinetic Characterisation of a Dye Decolourising Peroxidase from Streptomyces Lividans: New Insight into the Mechanism of Anthraquinone Dye Decolourisation. Dalt. Trans. 2017, 46, 9420–9429. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, L.; Guo, W.; Jia, L.; Fu, Y.; Gui, S.; Lu, F. Cloning, Expression, and Characterization of a Thermostable and PH-Stable Laccase from Klebsiella Pneumoniae and Its Application to Dye Decolorization. Process Biochem. 2017, 53, 125–134. [Google Scholar] [CrossRef]
- Arregui, L.; Ayala, M.; Gómez-Gil, X.; Gutiérrez-Soto, G.; Hernández-Luna, C.E.; Herrera De Los Santos, M.; Levin, L.; Rojo-Domínguez, A.; Romero-Martínez, D.; Saparrat, M.C.N.; et al. Laccases: Structure, Function, and Potential Application in Water Bioremediation. Microb. Cell Fact. 2019, 18, 200. [Google Scholar] [CrossRef]
- Chen, C.; Shrestha, R.; Jia, K.; Gao, P.F.; Geisbrecht, B.V.; Bossmann, S.H.; Shi, J.; Li, P. Characterization of Dye-Decolorizing Peroxidase (DyP) from Thermomonospora Curvata Reveals Unique Catalytic Properties of A-Type DyPs. J. Biol. Chem. 2015, 290, 23447–23463. [Google Scholar] [CrossRef]
- Kokkonen, P.; Beier, A.; Mazurenko, S.; Damborsky, J.; Bednar, D.; Prokop, Z. Substrate Inhibition by the Blockage of Product Release and Its Control by Tunnel Engineering. RSC Chem. Biol. 2021, 2, 645–655. [Google Scholar] [CrossRef]
- Benslama, O.; Mansouri, N.; Arhab, R. In Silico Investigation of the Lignin Polymer Biodegradation by Two Actinomycetal Peroxidase Enzymes. Mater. Today Proc. 2022, 53, 1–5. [Google Scholar] [CrossRef]
- Gilbert, R.G.; Hess, M.; Jenkins, A.D.; Jones, R.G.; Kratochvíl, P.; Stepto, R.F.T. Dispersity in Polymer Science (IUPAC Recommendations 2009). Pure Appl. Chem. 2009, 81, 351–353. [Google Scholar]
- Kim, K.H.; Kim, C.S. Recent Efforts to Prevent Undesirable Reactions from Fractionation to Depolymerization of Lignin: Toward Maximizing the Value from Lignin. Front. Energy Res. 2018, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Vignali, E.; Gigli, M.; Cailotto, S.; Pollegioni, L.; Rosini, E.; Crestini, C. The Laccase-Lig Multienzymatic Multistep System in Lignin Valorization. ChemSusChem 2022, 15, e202201147. [Google Scholar] [CrossRef]
- Rahmanpour, R.; King, L.D.W.; Bugg, T.D.H. Identification of an Extracellular Bacterial Flavoenzyme That Can Prevent Re-Polymerisation of Lignin Fragments. Biochem. Biophys. Res. Commun. 2017, 482, 57–61. [Google Scholar] [CrossRef]
- Musengi, A.; Durrell, K.; Prins, A.; Khan, N.; Agunbiade, M.; Kudanga, T.; Kirby-McCullough, B.; Pletschke, B.I.; Burton, S.G.; Le Roes-Hill, M. Production and Characterisation of a Novel Actinobacterial DyP-Type Peroxidase and Its Application in Coupling of Phenolic Monomers. Enzyme Microb. Technol. 2020, 141, 109654. [Google Scholar] [CrossRef]






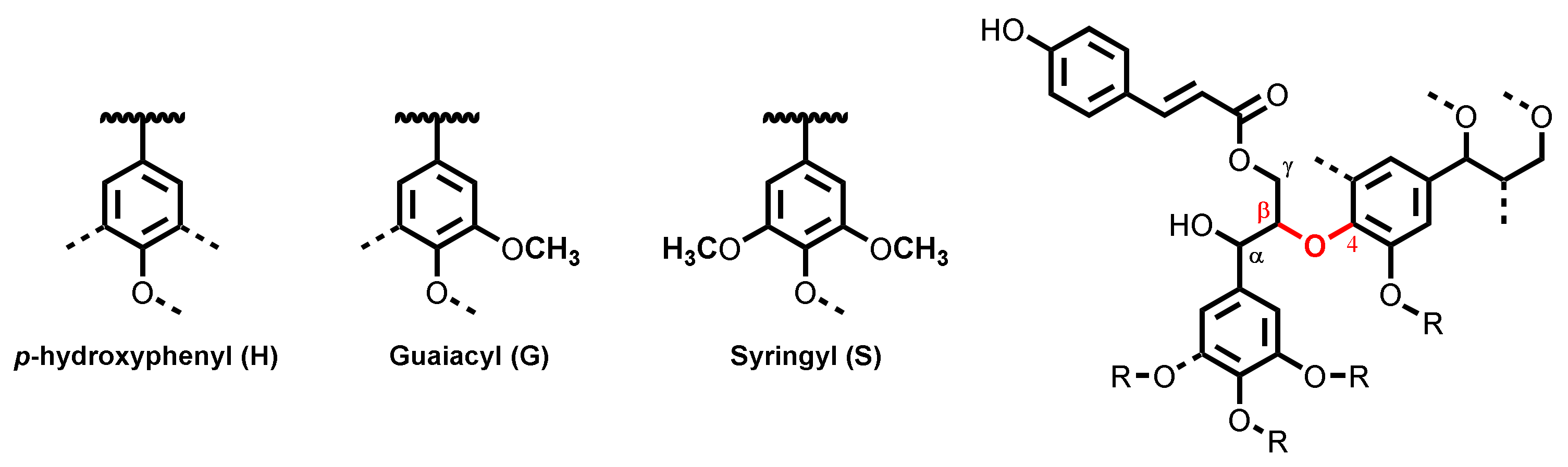{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ScDyP | Locus Tag | Annotation 1 |
|---|---|---|
| ScDyPB | SCO7193 | putative iron-dependent peroxidase |
| SCO7192 | sigma factor | |
| SCO7194 | putative polysaccharide biosynthesis protein | |
| ScDyP1A | SCO2276 | ferrous iron transport peroxidase |
| SCO2272, SCO2274 | heme ABC transporter | |
| SCO2273 | hemin transport system permease | |
| SCO2275 | ferrous iron transport periplasmic protein | |
| SCO2277 | ferrous iron transport permease | |
| SCO2278 | hypothetical protein | |
| ScDyP2A 2 | SCO3963 | DyP-type peroxidase family protein |
| SCO3971 | conserved hypothetical protein | |
| SCO3970 | Xaa-Pro aminopeptidase | |
| SCO3969 | possible ATP/GTP-binding protein | |
| SCO3968 | putative integral membrane protein | |
| SCO3967 | putative membrane protein | |
| SCO3966, SCO3965 | putative copper metallochaperone | |
| SCO3964 | copper transport protein | |
| SCO3962 | prephenate dehydratase | |
| SCO3961 | seryl-tRNA synthetase | |
| SCO3960 | hydrolase (HAD superfamily) |
| Rz Value | ScDyP1A | ScDyP2A | ScDyPB | ScDyP1A CO | ScDyP2A CO |
|---|---|---|---|---|---|
| Before reconstitution with hemin | 0.78 ± 0.02 | 0.51 ± 0.02 | 2.36 ± 0.04 | 2.01 ± 0.11 | 2.02 ± 0.01 |
| After reconstitution with hemin | 1.33 ± 0.08 | 1.14 ± 0.02 | 2.32 ± 0.07 | 1.92 ± 0.03 | 2.17 ± 0.03 |
| ScDyP1A | ScDyP2A | ScDyPB | |
|---|---|---|---|
| KM (mM) | 1.4 ± 0.15 | 2.3 ± 0.9 | 0.08 ± 0.02 |
| kcat (s−1) | 7.6 ± 1.1 | 31.7 ± 14.6 | 48.6 ± 5.7 |
| kcat/KM (mM−1·s−1) | 5.4 ± 0.2 | 13.8 ± 1.3 | 607 ± 95.2 |
| Ki (mM) | 4.3 ± 1.4 | 1.6 ± 1.0 | 0.4 ± 0.1 |
| Mxg-Lignin | Aspen Lignin | |||||
|---|---|---|---|---|---|---|
| Mn | Mw | ĐM | Mn | Mw | ĐM | |
| Without enzyme | 1509 ± 10 | 2435 ± 35 | 1.61 | 2772 ± 51 | 5533 ± 53 | 2.0 |
| +ScDyP1A | 1502 ± 8 | 2357 ± 26 | 1.57 | 2589 ± 12 | 4705 ± 72 | 1.82 |
| +ScDyP2A | 1489 ± 11 | 2332 ± 45 | 1.57 | 2429 ± 106 | 4371 ± 258 | 1.8 |
| +ScDyPB | 1856 ± 21 | 2921 ± 28 | 1.57 | 2994 ± 33 | 5607 ± 79 | 1.87 |
| (A) Unit | Compound Name | Without Treatment | ScDyP1A | ScDyP2A | ScDyPB |
|---|---|---|---|---|---|
| H | p-Hydroxycinnamic acid, ethyl ester | 100 | 78 | 78 | 2 |
| G | Ethyl (E)-ferulate | 100 | 67 | 75 | 2 |
| G | (E)-4-(3-Hydroxyprop-1-en-1-yl)-2-methoxyphenol | 100 | 77 | 69 | 9 |
| G | Glyceryl ferulate | 100 | 87 | 95 | nd * |
| S | trans-Sinapyl alcohol | 100 | 63 | 69 | 5 |
| (B) Unit | Compound Name | Without Treatment | ScDyP1A | ScDyP2A | ScDyPB |
| S | trans-Sinapyl alcohol | 100 | 53 | 55 | 37 |
| G | (E)-4-(3-Hydroxyprop-1-en-1-yl)-2-methoxyphenol | 100 | 44 | 52 | 16 |
| S | trans-Sinapaldehyde | 100 | 64 | 68 | 53 |
| G | Butyrovanillone | 100 | 92 | 73 | nd * |
| G | 2-Propanone, 1-hydroxy-3-(4-hydroxy-3-methoxyphenyl)- | 100 | nd * | nd * | nd * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pupart, H.; Jõul, P.; Bramanis, M.I.; Lukk, T. Characterization of the Ensemble of Lignin-Remodeling DyP-Type Peroxidases from Streptomyces coelicolor A3(2). Energies 2023, 16, 1557. https://doi.org/10.3390/en16031557
Pupart H, Jõul P, Bramanis MI, Lukk T. Characterization of the Ensemble of Lignin-Remodeling DyP-Type Peroxidases from Streptomyces coelicolor A3(2). Energies. 2023; 16(3):1557. https://doi.org/10.3390/en16031557
Chicago/Turabian StylePupart, Hegne, Piia Jõul, Melissa Ingela Bramanis, and Tiit Lukk. 2023. "Characterization of the Ensemble of Lignin-Remodeling DyP-Type Peroxidases from Streptomyces coelicolor A3(2)" Energies 16, no. 3: 1557. https://doi.org/10.3390/en16031557