
Effect of Glucose and Methylene Blue in Microbial Fuel Cells Using E. coli

Abstract
:1. Introduction
2. Materials and Methods
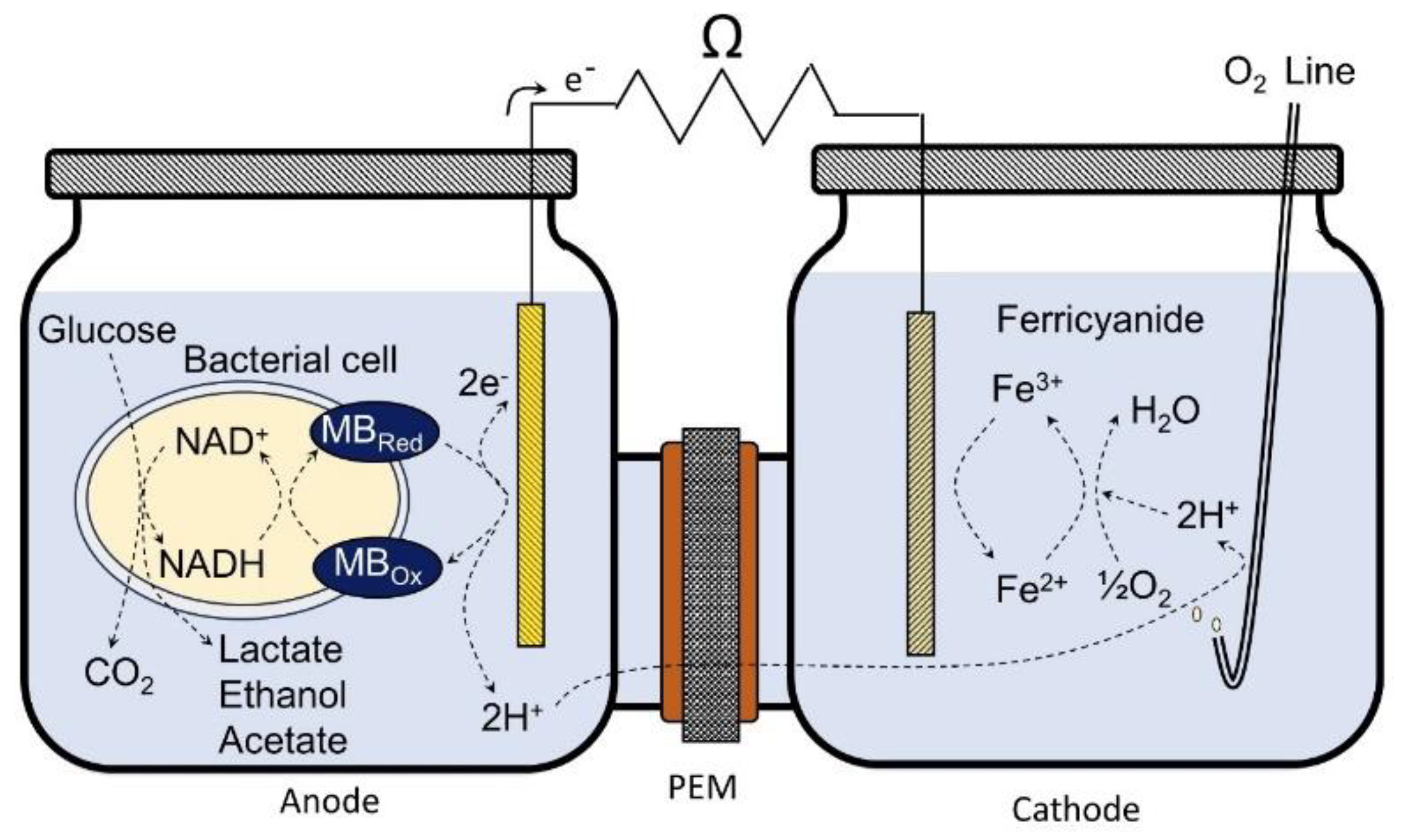
2.1. Reactor Design and Operation
2.2. Analytical Methods
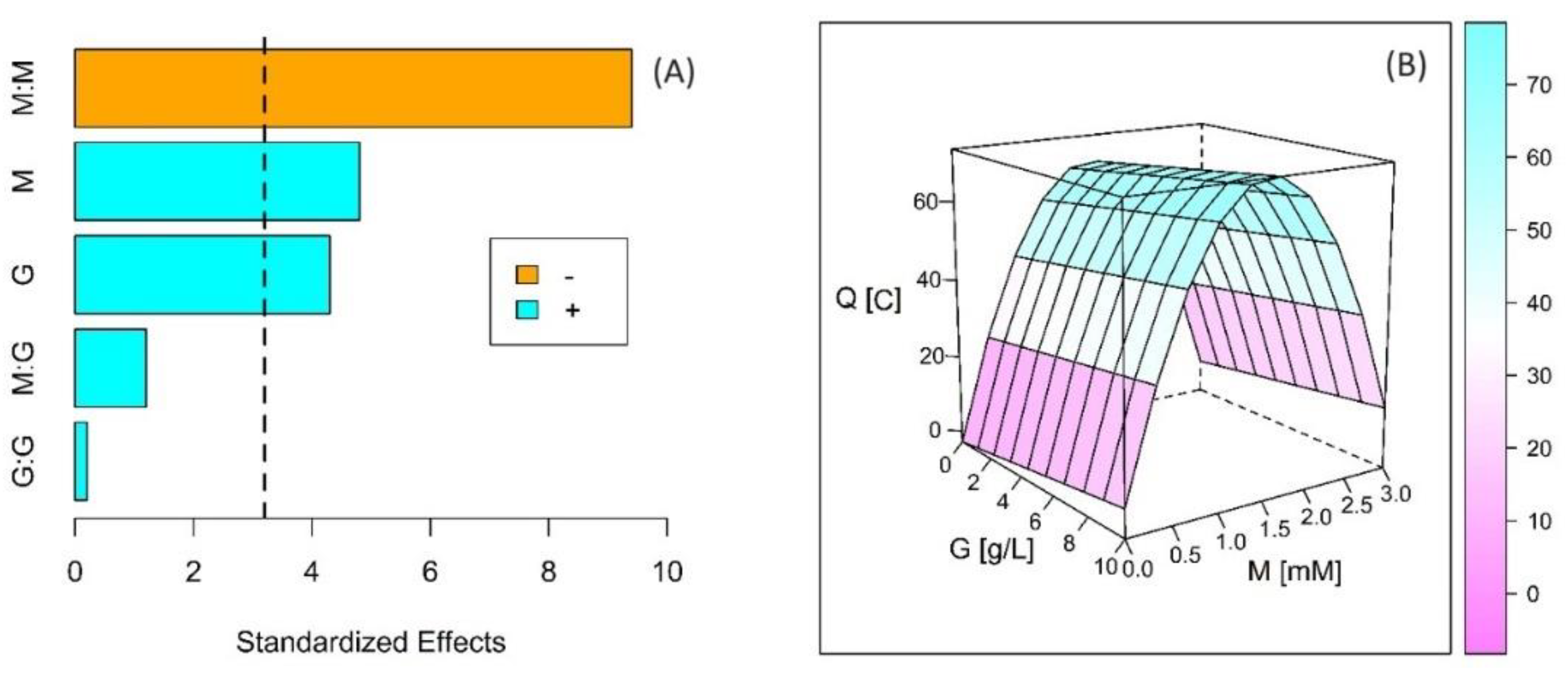
2.3. Experimental Design and Statistical Analysis
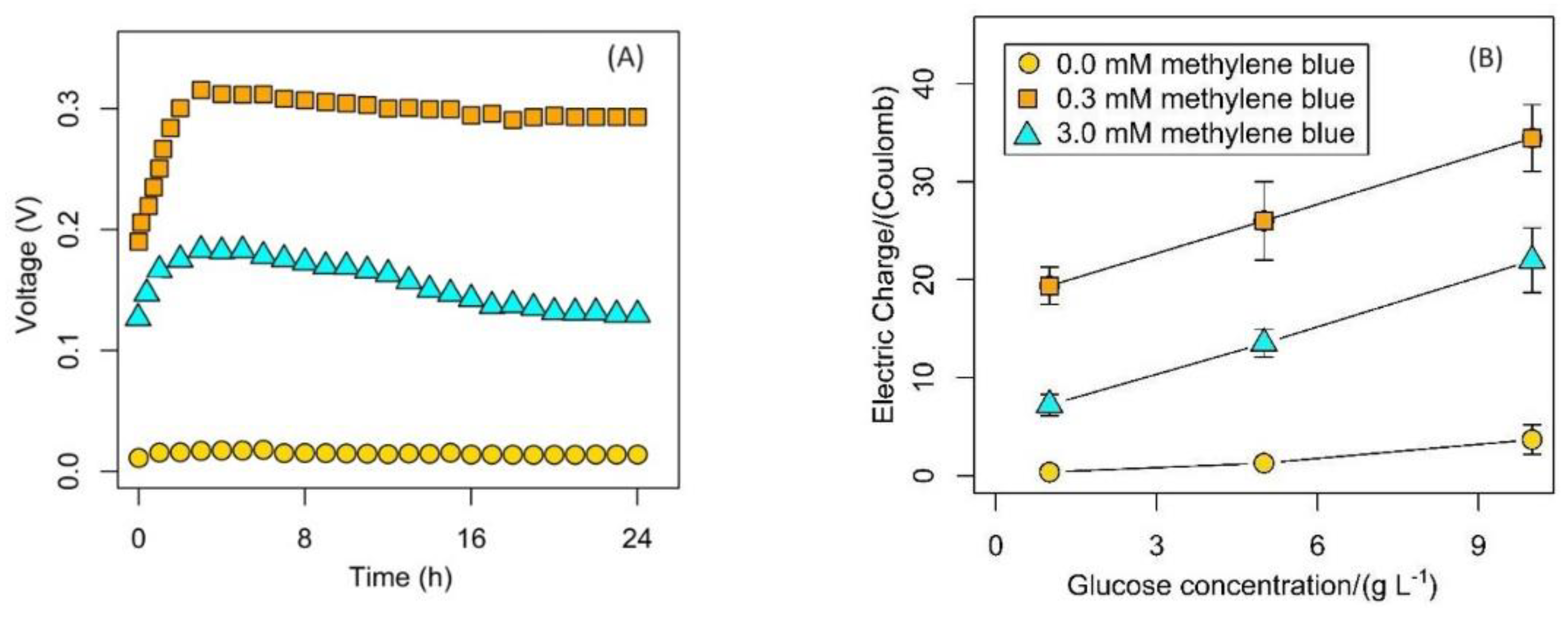
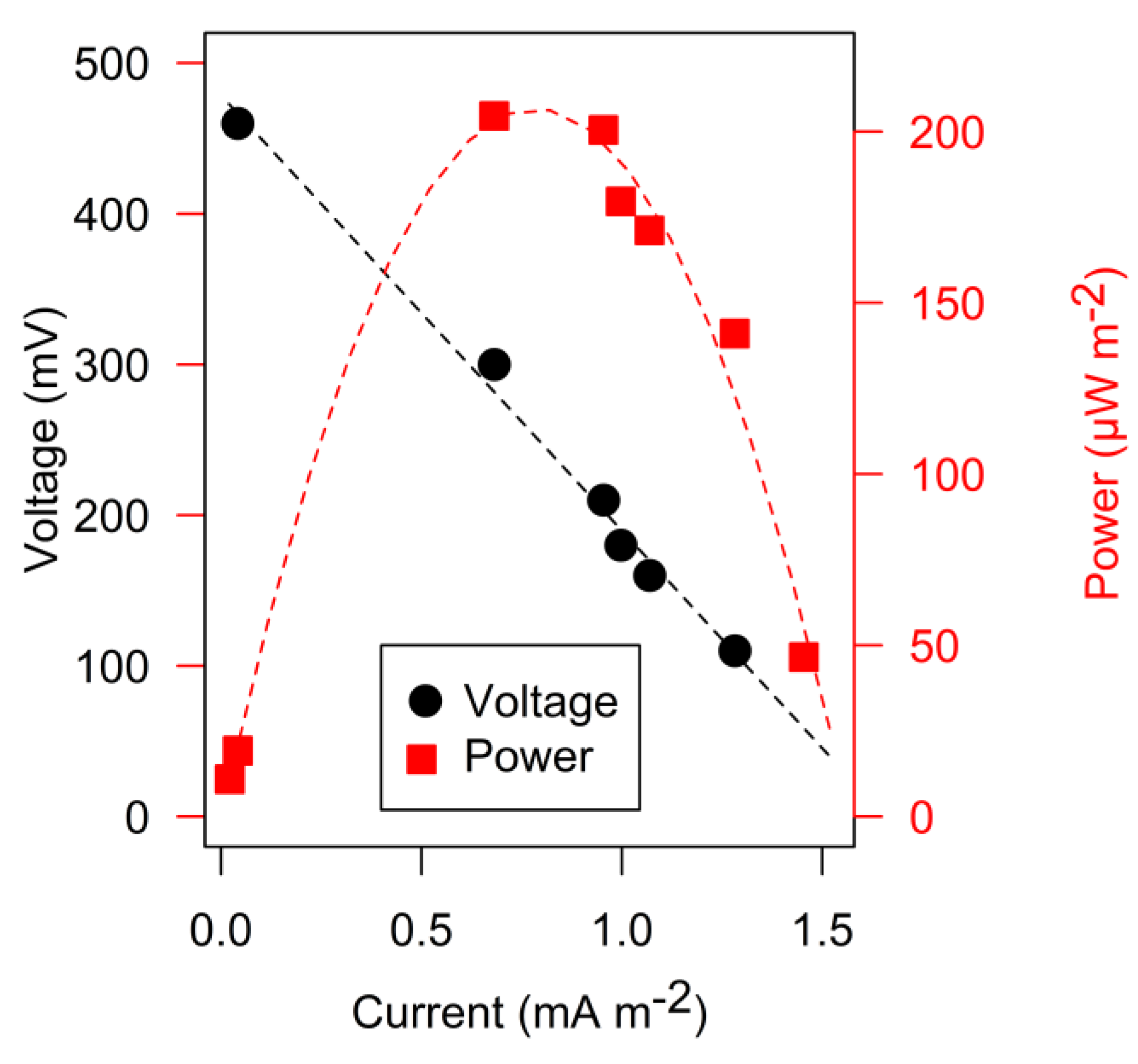
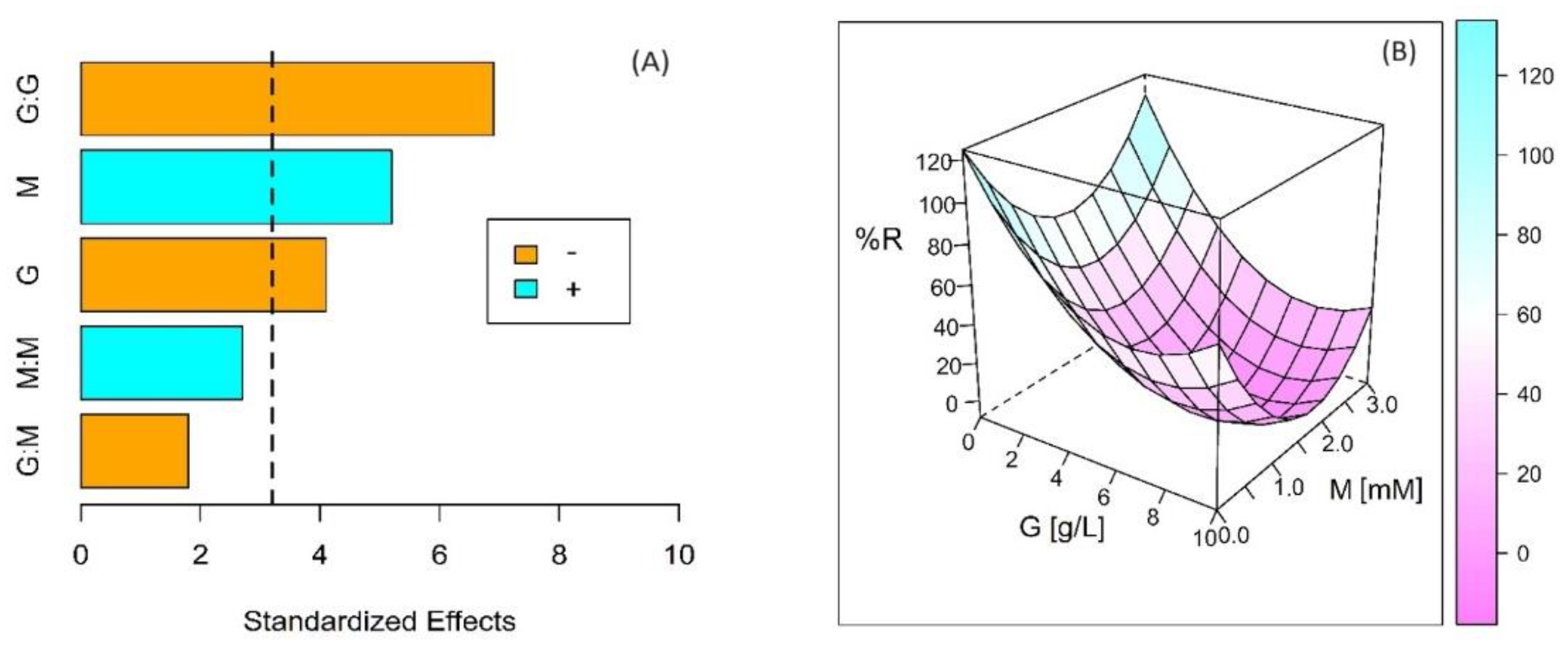
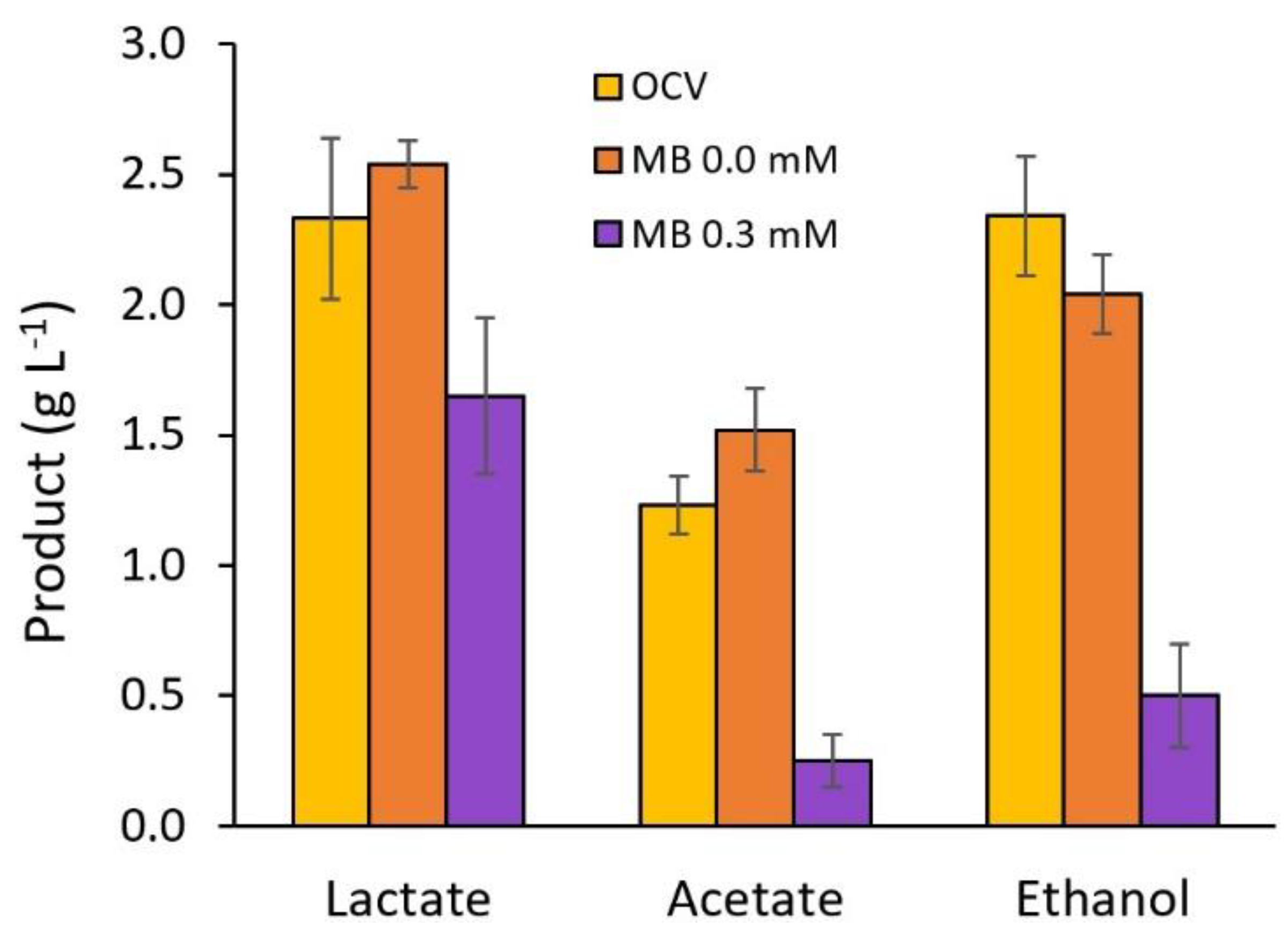
3. Results
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yolcan, O.O. World Energy Outlook and State of Renewable Energy: 10-Year Evaluation. Innov. Green Dev. 2023, 2, 100070. [Google Scholar] [CrossRef]
- Han, J.-H.; Bae, J.; Lim, J.; Jwa, E.; Nam, J.-Y.; Hwang, K.S.; Jeong, N.; Choi, J.; Kim, H.; Jeung, Y.-C. Acidification-Based Direct Electrolysis of Treated Wastewater for Hydrogen Production and Water Reuse. Heliyon 2023, 9, e20629. [Google Scholar] [CrossRef] [PubMed]
- Paraschiv, S.; Paraschiv, L.S.; Serban, A. An Overview of Energy Intensity of Drinking Water Production and Wastewater Treatment. Energy Rep. 2023, 9, 118–123. [Google Scholar] [CrossRef]
- Rocha-Meneses, L.; Luna-delRisco, M.; González, C.A.; Moncada, S.V.; Moreno, A.; Sierra-Del Rio, J.; Castillo-Meza, L.E. An Overview of the Socio-Economic, Technological, and Environmental Opportunities and Challenges for Renewable Energy Generation from Residual Biomass: A Case Study of Biogas Production in Colombia. Energies 2023, 16, 5901. [Google Scholar] [CrossRef]
- Tow, E.W.; Letcher, A.; Jaworowski, A.; Zucker, I.; Kum, S.; Azadiaghdam, M.; Blatchley, E.R.; Achilli, A.; Gu, H.; Melike, G.; et al. Modeling the Energy Consumption of Potable Water Reuse Schemes. Water Res. X 2021, 13, 100126. [Google Scholar] [CrossRef]
- Malik, S.; Kishore, S.; Dhasmana, A.; Kumari, P.; Mitra, T.; Chaudhary, V.; Kumari, R.; Bora, J.; Ranjan, A.; Minkina, T.; et al. A Perspective Review on Microbial Fuel Cells in Treatment and Product Recovery from Wastewater. Water 2023, 15, 316. [Google Scholar] [CrossRef]
- Li, X.; Abu-reesh, I.M.; He, Z. Development of Bioelectrochemical Systems to Promote Sustainable Agriculture. Agriculture 2015, 5, 367–388. [Google Scholar] [CrossRef]
- Ghangrekar, M.M.; Chatterjee, P. A Systematic Review on Bioelectrochemical Systems Research. Curr. Pollut. Rep. 2017, 3, 281–288. [Google Scholar] [CrossRef]
- Santoro, C.; Arbizzani, C.; Erable, B.; Ieropoulos, I. Microbial Fuel Cells: From Fundamentals to Applications. A Review. J. Power Sources 2017, 356, 225–244. [Google Scholar] [CrossRef]
- Goenka, R.; Mukherji, S.; Ghosh, P.C. Bioresource Technology Reports Characterization of Electrochemical Behaviour of Escherichia coli MTCC 1610 in a Microbial Fuel Cell. Bioresour. Technol. Rep. 2018, 3, 67–74. [Google Scholar] [CrossRef]
- Sturm-Richter, K.; Golitsch, F.; Sturm, G.; Kipf, E.; Dittrich, A.; Beblawy, S.; Kerzenmacher, S.; Gescher, J. Unbalanced Fermentation of Glycerol in Escherichia coli via Heterologous Production of an Electron Transport Chain and Electrode Interaction in Microbial Electrochemical Cells. Bioresour. Technol. 2015, 186, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lu, Q.; Li, K.; Xu, S.; Wang, X.; Chen, K.; Ouyang, P. Construction of an Electron Transfer Mediator Pathway for Bioelectrosynthesis by Escherichia coli. Front. Bioeng. Biotechnol. 2020, 8, 590667. [Google Scholar] [CrossRef] [PubMed]
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial Electron Transport and Energy Conservation—The Foundation for Optimizing Bioelectrochemical Systems. Front. Microbiol. 2015, 6, 575. [Google Scholar] [CrossRef] [PubMed]
- Price, C.E.; Driessen, A.J.M. Biogenesis of Membrane Bound Respiratory Complexes in Escherichia coli. Biochim. Biophys. Acta BBA Mol. Cell Res. 2010, 1803, 748–766. [Google Scholar] [CrossRef]
- Sugnaux, M.; Mermoud, S.; Ferreira, A.; Happe, M.; Fischer, F. Bioresource Technology Probing Electron Transfer with Escherichia coli: A Method to Examine Exoelectronics in Microbial Fuel Cell Type Systems. Bioresour. Technol. 2013, 148, 567–573. [Google Scholar] [CrossRef]
- Hoang, U.; Nguyen, P.; Grekov, D.; Geiselmann, J.; Stambouli, V.; Weidenhaupt, M.; Delabouglise, D. Electrochimica Acta Anodic Deposit from Respiration Metabolic Pathway of Escherichia coli. Electrochimica Acta 2014, 130, 200–205. [Google Scholar] [CrossRef]
- Zhang, T.; Cui, C.; Chen, S.; Yang, H.; Shen, P. The Direct Electrocatalysis of Escherichia coli through Electroactivated Excretion in Microbial Fuel Cell. Electrochem. Commun. 2008, 10, 293–297. [Google Scholar] [CrossRef]
- Kostylev, M.; Otwell, A.E.; Richardson, R.E.; Suzuki, Y. Cloning Should Be Simple: Escherichia coli DH5α-Mediated Assembly of Multiple DNA Fragments with Short End Homologies. PLoS ONE 2015, 10, e0137466. [Google Scholar] [CrossRef]
- Jahnke, J.P.; Sarkes, D.A.; Liba, J.L.; Sumner, J.J.; Stratis-Cullum, D.N. Improved Microbial Fuel Cell Performance by Engineering E. coli for Enhanced Affinity to Gold. Energies 2021, 14, 5389. [Google Scholar] [CrossRef]
- Nguyen, D.-T.; Tamura, T.; Tobe, R.; Mihara, H.; Taguchi, K. Microbial Fuel Cell Performance Improvement Based on FliC-Deficient E. coli Strain. Energy Rep. 2020, 6, 763–767. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Chen, Q.; Song, H.; He, Z.; Yang, Y. Enhanced Performance of Microbial Fuel Cells with Electron Mediators from Anthraquinone/Polyphenol-Abundant Herbal Plants. ACS Sustain. Chem. Eng. 2020, 8, 11263–11275. [Google Scholar] [CrossRef]
- Nawaz, A.; Hafeez, A.; Abbas, S.Z.; Haq, I.; Rafatullah, M. A State of the Art Review on Electron Transfer Mechanisms, Characteristics, Applications and Recent Advancements in Microbial Fuel Cells Technology. Green Chem. Lett. Rev. 2020, 13, 365–381. [Google Scholar] [CrossRef]
- Ng, I.S.; Hsueh, C.C.; Chen, B.Y. Electron Transport Phenomena of Electroactive Bacteria in Microbial Fuel Cells: A Review of Proteus Hauseri. Bioresour. Bioprocess. 2017, 4, 53. [Google Scholar] [CrossRef]
- Gemünde, A.; Lai, B.; Pause, L.; Krömer, J.; Holtmann, D. Redox Mediators in Microbial Electrochemical Systems. ChemElectroChem 2022, 9, e202200216. [Google Scholar] [CrossRef]
- Martinez, C.M.; Alvarez, L.H. Application of Redox Mediators in Bioelectrochemical Systems. Biotechnol. Adv. 2018, 36, 1412–1423. [Google Scholar] [CrossRef]
- Rahimnejad, M.; Najafpour, G.D.; Ghoreyshi, A.A.; Shakeri, M.; Zare, H. Methylene Blue as Electron Promoters in Microbial Fuel Cell. Int. J. Hydrogen Energy 2011, 36, 13335–13341. [Google Scholar] [CrossRef]
- Reiche, A.; Kirkwood, K.M. Bioresource Technology Comparison of Escherichia coli and Anaerobic Consortia Derived from Compost as Anodic Biocatalysts in a Glycerol-Oxidizing Microbial Fuel Cell. Bioresour. Technol. 2012, 123, 318–323. [Google Scholar] [CrossRef]
- Sharma, T.; Mohana, A.L.; Chandra, T.S.; Ramaprabhu, S. Development of Carbon Nanotubes and Nanofluids Based Microbial Fuel Cell. Int. J. Hydrogen Energy 2008, 33, 6749–6754. [Google Scholar] [CrossRef]
- Ojima, Y.; Kawaguchi, T.; Fukui, S.; Kikuchi, R.; Terao, K.; Koma, D. Promoted Performance of Microbial Fuel Cells Using Escherichia coli Cells with Multiple—Knockout of Central Metabolism Genes. Bioprocess Biosyst. Eng. 2019, 43, 323–332. [Google Scholar] [CrossRef]
- Popov, A.L.; Kim, J.R.; Dinsdale, R.M.; Esteves, S.R.; Guwy, A.J.; Premier, G.C. The Effect of Physico-Chemically Immobilized Methylene Blue and Neutral Red on the Anode of Microbial Fuel Cell. Biotechnol. Bioprocess Eng. 2012, 370, 361–370. [Google Scholar] [CrossRef]
- Aiyer, K.S. Synergistic Effects in a Microbial Fuel Cell between Co-Cultures and a Photosynthetic Alga Chlorella Vulgaris Improve Performance. Heliyon 2021, 7, e05935. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Xu, H.; Xiao, Y.; Calhoun, M.C.; Crisostomo, D.; Cliffel, D.E. The Utilization of Eschericia Coli and Shewanella Oneidensis for Microbial Fuel Cell. In IOP Conference Series: Materials Science and Engineering, Proceedings of the 3rd International Conference on Chemical Engineering Sciences and Applications 2017 (3rd ICChESA 2017), Banda Aceh, Indonesia, 20–21 September 2017; IOP Publishing Ltd.: Bristol, UK, 2018; Volume 334, pp. 1–6. [Google Scholar] [CrossRef]
- del Campo, A.G.; Canizares, P.; Lobato, J.; Rodrigo, M.; Fernandez Morales, F.J. Effects of External Resistance on Microbial Fuel Cell’s Performance. In Environment, Energy and Climate Change II. The Handbook of Environmental Chemistry; Lefebvre, G., Jiménez, E., Cabañas, B., Eds.; Springer: Cham, Switzerland, 2014; pp. 41–53. ISBN 1433-6863r978-3-642-03970-6. [Google Scholar]
- Moscoviz, R.; Toledo-Alarcón, J.; Trably, E.; Bernet, N. Electro-Fermentation: How to Drive Fermentation Using Electrochemical Systems. Trends Biotechnol. 2016, 34, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Zaitoun, M.A. A Kinetic-Spectrophotometric Method for the Determination of Glucose in Solutions. J. Anal. Chem. 2006, 61, 1010–1014. [Google Scholar] [CrossRef]
- Leon-Fernandez, L.F.; Rodrigo, M.A.; Villaseñor, J.; Fernandez-Morales, F.J. Electrocatalytic Dechlorination of 2,4-Dichlorophenol in Bioelectrochemical Systems. J. Electroanal. Chem. 2020, 876, 114731. [Google Scholar] [CrossRef]
- Taskan, E.; Ozkaya, B.; Hasar, H. Effect of Different Mediator Concentrations on Power Generation in MFC Using Ti-TiO2 Electrode. Int. J. Energy Sci. 2014, 4, 9–11. [Google Scholar] [CrossRef]
- Chen, Y.-A.; Yang, H.; Ouyang, D.; Liu, T.; Liu, D.; Zhao, X. Construction of Electron Transfer Chains with Methylene Blue and Ferric Ions for Direct Conversion of Lignocellulosic Biomass to Electricity in a Wide pH Range. Appl. Catal. B Environ. 2020, 265, 118578. [Google Scholar] [CrossRef]
- Gomes, T.F.; Pedrosa, M.M.; de Toledo, A.C.L.; Arnoni, V.W.; dos Santos Monteiro, M.; Piai, D.C.; Sylvestre, S.H.Z.; Ferreira, B. Bactericide Effect of Methylene Blue Associated with Low-Level Laser Therapy in Escherichia coli Bacteria Isolated from Pressure Ulcers. Lasers Med. Sci. 2018, 33, 1723–1731. [Google Scholar] [CrossRef]
- Nicu, A.I.; Pirvu, L.; Vamanu, A.; Stoian, G. The European Beech Leaves Extract Has an Antibacterial Effect by Inducing Oxidative Stress. Romanian Biotechnol. Lett. 2016, 22, 12071–12080. [Google Scholar]
- Ash, S.R.; Steczko, J.; Brewer, L.B.; Winger, R.K. Microbial Inactivation Properties of Methylene Blue—Citrate Solution. ASAIO J. 2006, 52, 17A. [Google Scholar] [CrossRef]
- Park, D.H.; Zeikus, J.G. Electricity Generation in Microbial Fuel Cells Using Neutral Red as an Electronophore. Appl. Environ. Microbiol. 2000, 66, 1292–1297. [Google Scholar] [CrossRef]
- Schmitz, S.; Rosenbaum, M.A. Boosting Mediated Electron Transfer in Bioelectrochemical Systems with Tailored Defined Microbial Cocultures. Biotechnol. Bioeng. 2018, 115, 2183–2193. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.R.; Navidjouy, N.; Rahimnejad, M.; Alizadeh, S.; Samarghandi, M.R.; Nematollahi, D. Effect of Different Concentrations of Substrate in Microbial Fuel Cells toward Bioenergy Recovery and Simultaneous Wastewater Treatment. Environ. Technol. 2022, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Förster, A.H.; Gescher, J. Metabolic Engineering of Escherichia coli for Production of Mixed-Acid Fermentation End Products. Front. Bioeng. Biotechnol. 2014, 2, 1–12. [Google Scholar] [CrossRef]
- Pinhal, S.; Ropers, D.; Geiselmann, J.; de Jong, H. Acetate Metabolism and the Inhibition of Bacterial Growth by Acetate. J. Bacteriol. 2019, 201, 1–19. [Google Scholar] [CrossRef]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial Mechanism of Lactic Acid on Physiological and Morphological Properties of Salmonella Enteritidis, Escherichia coli and Listeria Monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Roe, A.J.; O’Byrne, C.; McLaggan, D.; Booth, I.R. Inhibition of Escherichia coli Growth by Acetic Acid: A Problem with Methionine Biosynthesis and Homocysteine Toxicity. Microbiology 2002, 148, 2215–2222. [Google Scholar] [CrossRef]
- Xiang, K.; Qiao, Y.; Ching, C.B.; Li, C.M. GldA Overexpressing-Engineered E. coli as Superior Electrocatalyst for Microbial Fuel Cells. Electrochem. Commun. 2009, 11, 1593–1595. [Google Scholar] [CrossRef]
- Ren, J.; Li, N.; Du, M.; Zhang, Y.; Hao, C.; Hu, R. Study on the Effect of Synergy Effect between the Mixed Cultures on the Power Generation of Microbial Fuel Cells. Bioengineered 2021, 12, 844–854. [Google Scholar] [CrossRef]
- Vassilev, I.; Averesch, N.J.H.; Ledezma, P.K.M. Anodic Electro-Fermentation: Empowering Anaerobic Production Processes via Anodic Respiration. Biotechnol. Adv. 2021, 48, 107728. [Google Scholar]
- Rossi, R.; Fedrigucci, A.; Setti, L. Characterization of Electron Mediated Microbial Fuel Cell by Saccharomyces Cerevisiae. Chem. Eng. Trans. 2015, 43, 337–342. [Google Scholar] [CrossRef]
- Alberto García Mogollón, C.; Carlos Quintero Díaz, J.; Omar Gil Posada, J. Production of Acetone, Butanol, and Ethanol by Electro-Fermentation with Clostridium Saccharoperbutylacetonicum N1-4. Bioelectrochemistry 2023, 152, 108414. [Google Scholar] [CrossRef]
- Engel, M.; Holtmann, D.; Ulber, R.; Tippkötter, N. Increased Biobutanol Production by Mediator-Less Electro-Fermentation. Biotechnol. J. 2019, 14, 1800514. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Kim, T.; Woo, H.M.; Um, Y. Electricity-Driven Metabolic Shift through Direct Electron Uptake by Electroactive Heterotroph Clostridiumpasteurianum. Sci. Rep. 2014, 4, 6961. [Google Scholar] [CrossRef] [PubMed]
- Moscoviz, R.; Trably, E.; Bernet, N. Electro-Fermentation Triggering Population Selection in Mixed-Culture Glycerol Fermentation. Microb. Biotechnol. 2018, 11, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, J.; Chen, Q.; Ma, D.; Zhang, G.; Cui, C.; Xin, Y.; Zhao, Y.; Hu, C. The Inhibiting Effect of Oxygen Diffusion on the Electricity Generation of Three-Chamber Microbial Fuel Cells. J. Power Sources 2020, 453, 227889. [Google Scholar] [CrossRef]
- Simoska, O.; Cummings, D.A., Jr.; Gaffney, E.M.; Langue, C.; Primo, T.G.; Weber, C.J.; Witt, C.E.; Minteer, S.D. Enhancing the Performance of Microbial Fuel Cells via Metabolic Engineering of Escherichia coli for Phenazine Production. ACS Sustain. Chem. Eng. 2023, 11, 11855–11866. [Google Scholar] [CrossRef]
- Jensen, H.M.; TerAvest, M.A.; Kokish, M.G.; Ajo-Franklin, C.M. CymA and Exogenous Flavins Improve Extracellular Electron Transfer and Couple It to Cell Growth in Mtr-Expressing Escherichia coli. ACS Synth. Biol. 2016, 5, 679–688. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G (g L−1) | M (mM) | I (mA m−2) | Qe (C) | %R |
|---|---|---|---|---|
| 1.0 | 0.0 | 0.048 ± 0.03 | 0.36 | 98.2 ± 1.4 |
| 1.0 | 0.3 | 0.974 ± 0.04 | 19.37 | 93.3 ± 1.0 |
| 1.0 | 3.0 | 0.432 ± 0.02 | 7.22 | 92.8 ± 2.7 |
| 5.0 | 0.0 | 0.081 ± 0.02 | 1.28 | 61.8 ± 7.1 |
| 5.0 | 0.3 | 1.434 ± 0.18 | 25.57 | 40.6 ± 2.8 |
| 5.0 | 3.0 | 0.832 ± 0.02 | 13.49 | 32.1 ± 6.8 |
| 10.0 | 0.0 | 0.058 ± 0.02 | 3.68 | 75.4 ± 6.6 |
| 10.0 | 0.3 | 2.037 ± 0.13 | 34.44 | 44.2 ± 2.8 |
| 10.0 | 3.0 | 1.201 ± 0.08 | 21.96 | 34.2 ± 3.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoya-Vallejo, C.; Gil Posada, J.O.; Quintero-Díaz, J.C. Effect of Glucose and Methylene Blue in Microbial Fuel Cells Using E. coli. Energies 2023, 16, 7901. https://doi.org/10.3390/en16237901
Montoya-Vallejo C, Gil Posada JO, Quintero-Díaz JC. Effect of Glucose and Methylene Blue in Microbial Fuel Cells Using E. coli. Energies. 2023; 16(23):7901. https://doi.org/10.3390/en16237901
Chicago/Turabian StyleMontoya-Vallejo, Carolina, Jorge Omar Gil Posada, and Juan Carlos Quintero-Díaz. 2023. "Effect of Glucose and Methylene Blue in Microbial Fuel Cells Using E. coli" Energies 16, no. 23: 7901. https://doi.org/10.3390/en16237901