1. Introduction
Flight is the most demanding form of locomotion, requiring aerodynamics, weight minimisation, balance and control. The fossil record shows ancient species of flying insects with similar body shapes to modern insects, indicating a remarkably stable evolved aeronautical solution to a diverse series of circumstances and habitats. With nature providing existence proofs of enduring solutions, it is useful to examine the function of all aeromechanical aspects of these designs.
Due to the importance of the wings, less attention has been paid to the role the body shape plays in the dynamics, performance and control of insect flight. Change in body shape can potentially be used for flight control as it can change the positions of centre of mass and centre of pressure [
1]. Even land animals have been observed to use body shape changes for control, including lizards and cheetahs [
2,
3]. In insects, strong abdominal steering reflexes have been observed as response to mechanical or visual stimuli. For instance, a study presented by [
4,
5] show that desert locusts (
Schistocerca gregaria), in response to an angled wind stimuli during tethered flight respond with large leg and abdominal movements. Fruit flies (
Drosophila melanogaster) also demonstrated similar responses to visual rotations [
6,
7,
8]. Additionally, in response to the speed of a translating visual pattern, modulation of the vertical abdominal angle was observed in honeybees (
Apis mellifera) [
9]. Moths (
Manduca sexta) also show strong abdominal responses to mechanical and visual rotations about pitch axis [
10]. Several videos by Rüppell show several dragonfly abdominal movements during flight [
11]. However, there has been little effort to quantify these effects [
12,
13] and no attempt to consider the flight performance implications of the mechanism in the literature. Aircraft performance in steady flight is mostly concerned with power requirements and energy consumption [
14]. Energy savings in flight from an articulated abdomen/tail are likely to emerge from torques associated with cancelling an off-center mass distribution, replacing expensive aerodynamic generation of torques. Maintenance of a torque when a structure is fixed to the ground needs no energy once applied. In practice, biological muscles holding a static torque opposing a load do consume energy [
15] while producing no mechanical power. When airborne, aerodynamic generation of a torque relative to the free stream is energetically expensive, ultimately leading to increased drag. A properly designed aircraft producing a torque in addition to lift in level flight is subject to higher drag, less maximum lift and requires more propulsive power to compensate for the loss.
The aircraft mathematical models developed in this paper are based on biologically inspired characteristics that emulate a dragonfly (
Odonata anisoptera). With two pairs of independently controlled, high aspect ratio wings and a high ratio of muscle to weight, dragonflies demonstrate superior flight performance compared to most other insect species. Dragonfly mastery of the air has been shown in a number of studies based on high speed video analysis of aerial combat and predation instances. High performance turning flight has been shown, with loads in turns exceeding 40 m/s
[
16] and the ability to takeoff while carrying more than three times their own body weight [
17]. Dragonflies have also been shown to pursue prey animals using the chasing strategies found in missiles [
18] and to have exceptionally high success rates in capturing aerial prey [
19,
20]. It has been demonstrated that they have surprisingly efficient glide performance comparable to model aircraft with higher Reynolds numbers (Re). Observed lift-to-drag ratios range from 3.5 to over 10 [
21,
22,
23]. Some species of dragonfly have been observed to migrate across hundreds of kilometres [
24], a difficult energetic achievement for such a small organism. Energy scavenging by dragonflies has also been observed, in which they soar on thermal updrafts and on rising air currents caused by slopes [
25].
The dragonfly body form is dominated by a long abdomen (weighing 31–35% of the total body mass), two pairs of comparatively high aspect ratio wings (weighing less than 2% of the total body mass), a dense thorax and a large head dominated by large eyes [
26]. The dragonfly abdomen is by no means a “tail” in the normal sense for vertebrates, neither is it analogous to traditional aircraft empennage. Aircraft tails are generally rigid structures on which there are aerodynamic surfaces, both horizontal and vertical. In many species of dragonfly, there are no aerodynamic structures at the distal end of the abdomen.
Figure 1a–d shows the typical abdomen shape of dragonfly families. Most dragonfly and damselfly abdomens are slender, as in
Figure 1a showing the silhouette of a dragonfly from the family Aeshnidae. Variations exist, for example, some “chasers” of the family Libellulidae have a shorter thicker abdomen as shown in
Figure 1b. Some “clubtails” of the family Gomphidae as shown in
Figure 1c, have a pronounced club shaped structure terminating the abdomen from which the family draws its name, strengthening a hypothesis of a predominantly inertial role in this case at least. Some “petaltails” of the family Petaluridae appear to have an aerodynamic structure terminating their abdomens (
Figure 1d). It might be relevant that the largest dragonfly species by some measures,
Petalura ingentissima, carries such a structure.
The abdomen is certainly required to exist regardless of flight control since it contains the digestive tract and anatomical features related to reproduction. It is also required to be actuated to some extent for mating and oviposition [
28], as well as the likely evolutionary requirement to be neither long and rigid or long and flexible. Thus, the cost of articulating the abdomen is only the additional muscle mass required to achieve rapid movements. Therefore, we focus on two observed abdominal motions and their utilisation for correction of imbalance in steady level flight and for a longitudinal pull up maneuver, which are:
Tail contraction and extension, illustrated in
Figure 2a and will be referred to as “Model 1” and,
Tail up down wag movement, illustrated in
Figure 2b and will be referred to as “Model 2”.
In the remainder of this paper we introduce the performance and dynamics model used to demonstrate and quantify the energetics of these techniques.
2. System Model
We have used the most accessible and analytical means possible for the analysis to avoid resorting to poorly understood, case specific or possibly methodological artefacts from fluid simulation. We have also focused on the case of longitudinal flight with fixed wings, which results in acceptable fidelity with a focus on wings with attached air flow and clear applicable outcomes. In demonstrating how features of dragonfly anatomy could save energy in flight, an aircraft in the well understood scale of small fixed wing models with Reynolds numbers in thousands, rather than dragonflies with Reynolds numbers in hundreds, was used. The discipline of aircraft performance contains well established methods and tools for performance estimation of fixed wing aircraft, which also yield algebraic expressions that are amenable to new derivations, optimisation and predictions. Wherever possible, algebraic expressions and conventional aerodynamic techniques were used, with the nomenclature commonly used in the aerospace literature [
29,
30].
For the purposes of isolating inertial from aerodynamic forces on the tail, the aerodynamic effects of the tail were ignored as these effects were expected to have little influence on the aerodynamics of the aircraft, compared to the wings, legs and thorax. A particular advantage of an inertial actuator is that it will continue to produce torques at low speeds, even when the aircraft is not flying or in an aerodynamic stall. For this reason, we might expect to see large abdominal motions at low speeds. To simplify this problem, the articulated tail was placed in the analytical framework developed for small fixed wing aircraft. Dragonflies have efficient wings, credited by Wakeling with glide slope of 6.3:1 [
21]. Many documented observations of dragonflies engaging in extended periods of soaring and gliding exist [
21,
25,
31,
32], so, fixed wing cruising and gliding flight is an appropriate and accessible aspect of their flight for our analysis. Although the analysis in this study focuses on fixed wing modes of flight, the qualities of the outcomes should apply to flapping wing flight. The assumption of the analysis is that the abdomen/tail is a mass that is already present and required. We focus on the question of the potential benefits to maneuverability and energetics of the inertial tail.
2.1. Reference Frames
The aircraft is modelled as a collection of rigid bodies. The dragonfly head to thorax region, including the wings are abstracted as a rigid body
with a mass
and will be referred to as the “body”. The abdomen, interchangeably referred to as the “tail” in this paper, is modelled as an added mass,
, concentrated at the tip of the tail
. Reference frames used for the development of the equations of motion are defined as shown in
Figure 3. The North-East-Down (NED) coordinate system which is commonly used in aeronautics is adapted for each of the Earth, body and tail [
33]. The coordinates in each frame are
,
and
respectively. The origin of the inertial frame (I) is fixed at an arbitrary point relative to the earth’s surface. The origin of the body-fixed frame (B) can be arbitrarily selected to be any point on the body of the aircraft. The tail reference frame (T), originates from the centre of gravity (
) of the tail point mass with its orientation the same as that of the body frame coordinate system when the tail is undeflected. The whole aircraft (
) will be referred to as the rigid body
. In addition, four reference points,
and
j are introduced. These points represent the locations of the center of mass of the body only, the whole aircraft, the tail and the tail joint location respectively.
2.2. Equations of Motion
Whilst only longitudinal flight was examined in this study, for completedness, the full non-linear dynamic model of the aircraft is presented. It is common practice to derive equations of motion referenced to the combined of an aircraft, however, to properly reflect the effects associated with dynamic changes in combined position, the dynamic equations presented in this study are referenced to point b which is the centre of mass of the rigid body in the body-fixed reference frame B. When the reference frame of a vector or tensor is not specified, it automatically means it is written in the body-fixed reference frame B.
2.2.1. Kinematics
The translational and rotational kinematic equations for an aircraft are available in the flight dynamics literature. In matrix form, the translational kinematic equations are [
29]
where
represents the position
of the aircraft relative to the inertial frame,
is the rotation matrix from body-fixed frame to inertial frame and
is the aircraft body velocity components
.
Since the intention is not to model very large attitude angles, the standard aerospace rotational kinematic equations using Euler angles
are chosen to represent the attitude of the aircraft and are given by [
29]
where
are the body frame angular velocities of the aircraft.
2.2.2. Dynamics
Two sets of dynamic equations are developed for the two modes of tail actuation considered in this study. The first set of equations represent Model 1, which was used for modelling the contraction and extension of the dragonfly tail during flight (see
Figure 2a). The second set of equations representing Model 2, was used to model the up and down wag movement of the dragonfly tail during flight, shown in
Figure 2b. The equations are derived based on the following assumptions:
Earth is flat and non-rotating and is the inertial frame.
The aircraft is electrically powered, has a constant total mass but varying mass distribution.
The mass and mass distribution of the rigid body remains the same throughout the flight.
Model 1 Aircraft Dynamics
The translational and rotational dynamic equations are given by Equations (
3) and (
4) respectively [
34,
35]
where
and
are the position, velocity and acceleration of the tail mass with respect to point
b in the body frame.
is the body frame angular velocity components
and
m is the total mass of the aircraft.
F and
M represent the total forces and moments acting on the aircraft respectively.
The change in
position with respect to point
b, written in the body frame is expressed as
In addition, the inertia matrix of the whole system about point
b is given by
where
is expressed as
and
is a
identity matrix.
The elements of the inertia tensor
J are
Model 2 Aircraft Dynamics
The translational and rotational dynamic equations are represented by Equations (
9) and (
10) respectively [
34,
35,
36,
37]:
where
and
are the angular velocity and acceleration of the tail mass with respect to the body frame respectively. The change in
position and the total inertia matrix of the whole system are also estimated using Equations (
5)–(
7), however,
2.3. Forces and Moments
The forces and moments acting on an aircraft are mainly due to aerodynamics, propulsion and gravity. In this study the same aerodynamic and propulsive models were used for both dynamic models developed. These models are well established for fixed wing aircraft in the flight dynamics literature such as [
33,
38] and will not be discussed in this paper. It is important, however, to note that we assume the aircraft thrust aligns with the longitudinal axis and hence, does not produce any moments.
The gravitational force model for both dynamic models are also the same and are detailed in [
33], however, the gravitational moments for both tail actuation modes differ. For Model 1, the gravitational moment
is
where
g is the acceleration due to gravity. However, for the Model 2, the gravitational moment produced by the tail is given by
2.4. Control Inputs
For propulsion, the control input used in this study is the thrust
. The aerodynamic control surfaces are the left and right elevons, denoted by
and
respectively and located on the trailing edges of the wings as shown in
Figure 3. Generally, elevons, depending on the desired flight regime, function as elevators or ailerons. The combined deflections of the elevons as elevators
or ailerons
are according to [
33]
With regards to the longitudinal motion of the aircraft model in this study, the two main functions of the elevators are for longitudinal trim and/or longitudinal control. When deflected, the camber of the airfoil of the wing is changed and the lift coefficient (
) changes consequently. Therefore, elevator deflection increases or decreases wing lift and pitching moment [
39]. The elevator deflection
, presented in this study is a function of the left and right elevons acting as left and right elevators respectively (
and
), such that
The tail mass linear displacement or angular deflection on the other hand, functions as an alternative moment generator, changing pitching moment [
34]. For Model 1, the control inputs are the tail mass positions
and for Model 2, the control inputs are the tail mass angular deflections
relative to the body frame. Where applicable, a downward deflection of a control surface or tail deflection is positive and an upward deflection is negative.
2.5. Energy Maneuverability
Maneuverability in flight involves the ability to perform a change, or a combination of changes in, altitude, direction and airspeed [
40]. As mentioned earlier, dragonflies demonstrate superior maneuverability as apex predators among insects. Energy maneuverability involves the analysis of maneuverability, expressed in terms of energy and energy rate. Although energy maneuverability is not directly concerned with electrical energy consumption, the use of appropriate energy maneuverability strategies can result in reduced electrical energy consumption.
Summing the forces acting on an aircraft, parallel to the flight path as shown in
Figure 4, the velocity change equation yields [
41,
42],
where
is the flight path angle.
The total energy or energy state of an aircraft, is expressed as the sum of kinetic and potential energy,
where
H is the height.
To enable comparison of maneuver performance of various aircraft, it is more convenient to use the specific energy
, which is the energy per unit mass. The specific energy is sometimes called the energy height,
The energy state of an aircraft can be changed by applying power. The rate of change of specific energy
, also known as specific excess power
is
Using the kinematic relation that
and the expression of
from Equation (
16), Equation (
19) becomes
5. Discussion
In this study, we have examined the effect of an inertial tail appendage on a relatively conventional fixed wing aircraft and considered performance, emphasising efficiency. For two longitudinal flight scenarios, we have shown how the abdominal movements, as observed in dragonflies, improve flight performance.
5.1. Correction of Imbalance in Steady Level Flight
It is safe to assume that passive stability for a four winged flapping craft, capable of nearly holonomic locomotion [
60], is not a necessary precondition in the same way that it is for a fixed wing aircraft. Yet, the stability of the surrogate fixed wing aircraft would be adversely affected by
variations, as a zero or negative stability margin would lead to a departure from controlled flight.
For Models 1 and 2 aircraft and for all cases relating to correction of imbalance in steady level flight, the plots for elevator deflection required to trim with respect to airspeed are consistent; the amount of elevator deflection required to trim decreased with increased speed because higher air speeds require less lift coefficient
and consequently less angle of attack [
39]. The power required plots as a function of airspeed also present consistent results as increased weight results in more lift required and consequently, an increase in induced drag (see
Figure 8 and
Figure 9) [
39].
Although the power required after both models were disturbed was obviously higher than the initial condition, performance improved in the form of lower power required compared to before the tail was moved to compensate for imbalance, however, the magnitude of power saved was different for each model due to the difference in initial configurations.
Specifically, for Model 1 with a linearly displaceable tail mass as in
Figure 2a, an average increase in elevator deflection of 161% was observed after the disturbance mass was introduced, compared to 55% after correction of imbalance was performed by linearly extending the tail. In addition, results showed an average of 12% increase in energy required after the disturbance mass was introduced, compared to 7% after correction of imbalance was performed by linearly extending the tail.
For Model 2 with a deflectable tail as in
Figure 2b, an average increase in elevator deflection of 116% was observed after the disturbance mass was introduced, compared to 46% after correction of imbalance was performed by moving the tail to its neutral position, which had zero deflection. The results showed an average of 13% increase in power required before, compared to 9% after correction of imbalance. Overall, for steady cruise, a 4–5% average in propulsive power savings is quite substantial for an electrically powered aircraft.
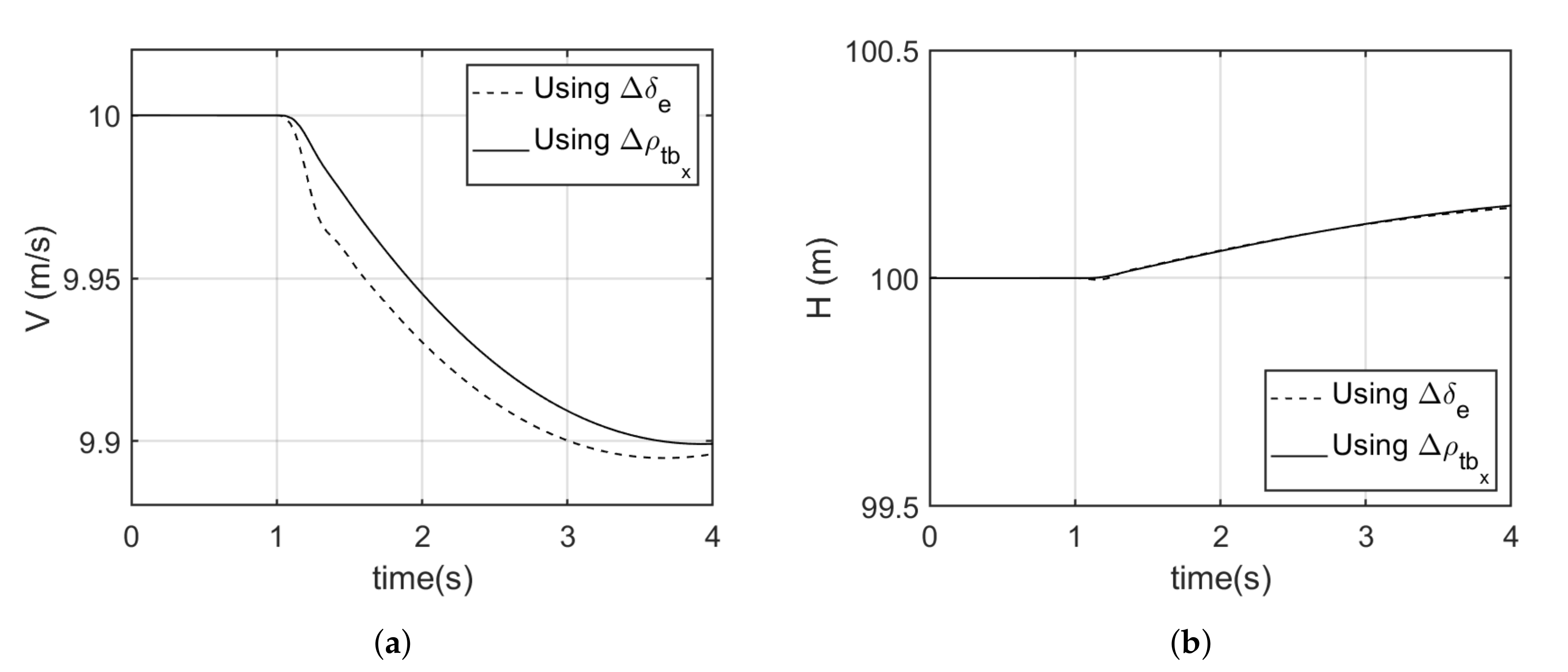
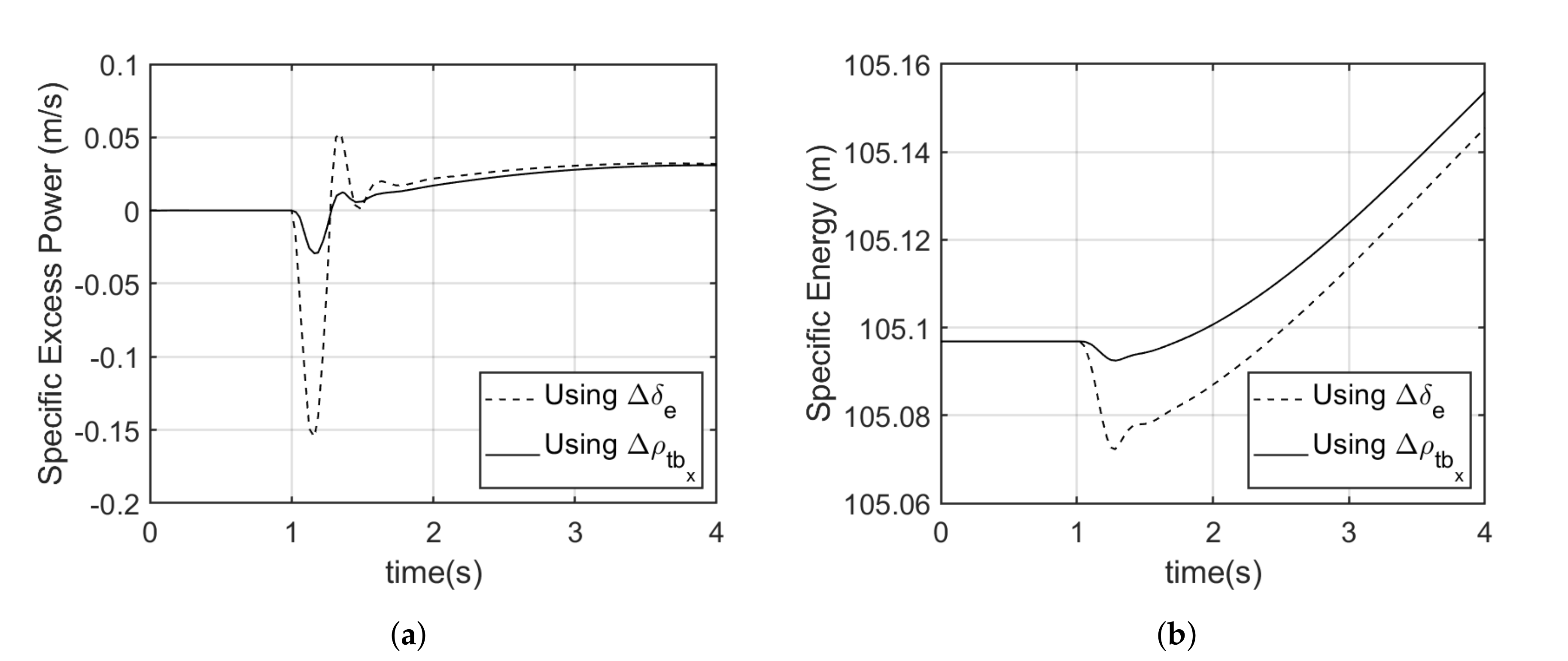
5.2. Pull up Maneuver
The two alternative moment generation mechanisms were able to achieve the same pull up maneuver as the elevator as shown in
Figure 10a and
Figure 12a. In the case of the Model 1, an average of 0.9% more specific energy was recorded when using tail displacement in comparison with using the elevator. Looked at across a short time period near the maneuver, the drop in specific excess power using tail movement was around −0.03 m/s, where it was −0.15 m/s using control surface deflection. Over a 4 s window, an average of 1.39% more excess power was recorded when using tail displacement in comparison to using the conventional elevator (see
Figure 9).
For the Model 2 aircraft, an average of 1% more specific energy was recorded when using tail angular deflection in comparison with using the elevator. Whereas, an average of 1.83% more excess power was recorded when using tail displacement in comparison with using the conventional elevator (see
Figure 11). Again, the transient specific excess power drop was negligible when using tail movement, but around 0.1m/s when using control surface deflection.
Generally, an aircraft that is able to maintain a higher specific energy and excess power has more maneuver advantage [
40]. The negative average
observed when using the elevator to initiate the maneuver results from the drag being greater than available thrust, which results in decreased energy. This is due to the additional drag induced as a result of elevator deflection [
33,
40].
Although the percentage savings in specific energy states and rates caused by using either of the tail actuation modes as opposed to using the conventional elevator may seem small in magnitude, they make a difference in combat. A dragonfly in pursuit of prey for instance, but particularly in territorial conflict, is engaged in pure aerial combat. In such bouts, dragonflies may be required to make the same or a combination of energy consuming maneuvers in sequence, in a life or death conflict to deplete the opponent’s energy. The transient advantage of using a more energy efficient control effector to initiate a maneuver is substantial by the standards of optimisation of flying systems.
Whilst the results obtained in this study illustrate the potential energy effectiveness demonstrated by insects in flight, fidelity was deliberately limited. The aircraft dynamics model could be made more comprehensive by including the actuator dynamics to quantify the energy required by the actuator to actuate the tail mass when correcting for the imbalance as well as for dynamic maneuvers. Given that the tail was modelled as a point mass and for reduced complexity, its aerodynamic properties were ignored. However, in analysing the stability of the hang-glider where control is achieved by the movement of the pilot relative to the wing, [
61] undertook experiments in the wind tunnel to evaluate the aerodynamics on a pilot. Results showed the lift and pitching moment were negligible, and although the drag was significant, its variation with AoA was small. Therefore, future studies should include the aerodynamic effects of actuated abdomens.
Abdominal postures should be explored in high speed footage of dragonflies when capturing prey and across the cycle of feeding and flying, possibly in existing high speed video footage such Georg Rüppell’s contributions that are publicly available at [
11].
6. Conclusions
We have mathematically expressed the basic means by which the abdomen of an efficient natural flyer like the dragonfly might be used to save energy and reduce peak power requirements in flight. The abdomen might save energy with reactive responses to achieve longitudinal balance and pull up maneuvers by manipulating moments. Active creation of torques is possible through movement of the abdomen. From analysis of a simulation model of a fixed wing aircraft with an articulated tail structure, we have shown that the amount of energy that might be saved by these techniques can be substantial. We have shown that flight power increases would be required to substitute for the transient effects that can be generated using the movement of the abdomen, while the possibility of trimming balance through tail posture might save energy for as long as the imbalance exists.
In translating these results to technological aircraft, there is a question of overall system integration since an abdomen may not have utility and would thus be simply excess weight. Aircraft may have similar problems to dragonflies in some applications, for example a variable payload weight, a need to fold or fit into a small space, combined with a benefit from high angular accelerations. The dragonfly also reveals that if there is a need to interact with the environment, in the dragonfly case this includes mating and oviposition, a long articulated abdomen is at least as useful as a long nose and intuitively less problematic.

 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



 represents the added mass.
represents the added mass.


 represents the added mass.
represents the added mass.