Introduction
Heavy metal contaminants in the environment are eventually deposited in soils in some form of a low solubility compound, such as pyrite [
28] or sorbed on surface-reactive phases, such as Fe and Mn oxides [
9,
22].While this phenomenon immobilizes the contaminants, thus limiting their effects upon biota and human health, it also places metal ions in an intimate contact with soil microbial community.
A number of studies were completed on the role of soil/sediment microbial community in heavy metal scavenging and release in recent decades [
2,
3,
7,
32].Those studies highlight the importance of metal-microbe interactions and microbial processes from sorption and desorption of metals on the cell surfaces to reduction/oxidation phenomena in determining the fate of heavy metals in the environment. The opposite side of the interaction, namely, how heavy metals at lower concentrations impact composition of a microbial community and functioning of specific pathways within that community is substantially less understood, especially in the circumneutral environments where metal levels are low and metals are not exceedingly mobile. Our knowledge of metal effects at lower, more environmentally relevant conditions, circumneutral pH, and response of metagenome to heavy metal challenge under these conditions is still limited to a handful of studies [46]. In contrast, highly acidic, high-metal conditions, such as those occurring in mine tailings have been thoroughly investigated [
7,
21, 48].
Although some heavy metals are required for life’s physiological processes (e.g., components of metalloenzymes), their excessive accumulation in living organisms is always detrimental. Generally, toxic metals cause enzyme inactivation, damage cells by acting as antimetabolites or form precipitates or chelates with essential metabolites. The impact of some toxic metals on human health has been reported by Forstner [
17]. Human diseases have resulted from consumption of cadmium-contaminated foods [
31,
37]. Likewise, Pb exposure can cause seizures, mental retardation, and behavioral disorders. The threat that heavy metals pose to human and animal health is aggravated by their low environmental mobility, even under high precipitations, and their long-term persistence in the environment. For instance, Pb, one of the more persistent metals, was estimated to have a soil retention time of 150 to 5000 years [44]. Also, the average biological half-life of Cd, another accumulation poison similar to lead, has been estimated to be about 18 years [
17]. As a result of low environmental mobility of those metals, a single contamination episode could set a stage for a long-term exposure of microbial community to metal, necessitating a long-term monitoring effort to assess metal effects.
Studies have shown that long-term heavy metal contamination of soils has harmful effects on soil microbial activity, especially microbial respiration [
16, 40]. Aside from long-term metal-mediated changes in soil enzyme activities, many reports have shown large reductions in microbial activity due to short-term exposure to toxic metals [
15,
23]. Bacterial activity, measured by thymidine incorporation technique, had been shown to be very sensitive to metal pollution both under laboratory [
12,
13] and field [38] conditions. Moreover, habitats that have high levels of metal contamination show lower numbers of microbes than uncontaminated habitats [
30, 41].
Enzyme activity is a soil property that is chemical in nature but has a direct biological origin. This activity arises from the presence of many types of enzymes that are present in the soil, and within soil microorganisms. From an assortment of enzymes present and active in soil, phosphatases are interesting groups of enzymes that catalyze the hydrolysis of phosphate from organic monoester linkages [47]. Phosphates released from such phosphatase action are very important to the plants and microorganisms that depend on soil for their phosphorus requirements.
It is well established that toxic effects of heavy metals are highly selective in the higher organisms. Specific organ targeting was shown for mercury [
29] and silver [
4] in invertebrates. Indications of specific inhibitory action of heavy metals have been produced in microbes as well [
18,
19]. Such selective targeting of specific enzymatic systems and pathways suggests that certain members of the microbial community would be more sensitive to heavy metal exposure than others, depending on the sensitivity of their critical metabolic pathways. Thus, while toxicity of heavy metals to microbes is a well established phenomenon, the effects of those metals upon specific enzymatic systems at lower (“sub-acute”) concentrations are not well known.
Denitrification is a natural microbial process converting nitrate to dinitrogen gas during anaerobic respiration. Such reduction occurs sequentially, with nitrate converted to nitrite, nitric oxide, nitrous oxide and, finally, nitrogen gas. A number of enzyme classes, mostly located in the periplasmic space, are involved in denitrification [39], with a number of corresponding genes that can be used as genetic markers for presence and expression of such enzymes in the soil metagenome. Based on their function, those enzymes can be broadly classified into nitrate reductases (converting nitrate to nitrite,
narG and
napA genetic markers correspond to those enzymes), nitrite reductases (nitrite to nitric oxide,
nirK and
nirS markers), nitric oxide reductases (nitric oxide to nitrous oxide,
norB marker) and nitrous oxide reductases (nitrous oxide to dinitrogen gas,
nosZ marker) [50]. The group of organisms capable of denitrification is not monophyletic; extensive horizontal gene transfer occurred early in the evolution of the pathway, resulting in nearly identical gene and enzyme forms in very distantly related organisms [
6, 39]. As a result, 16S rRNA, which serves as a standard marker to assess microbial community diversity, is essentially useless for studying denitrifying bacteria.
Effects of heavy metals upon the community of microbes responsible for nitrogen cycle are still largely unknown. Stephen et al. [46] analyzed the diversity changes of ammonia-oxidizing
Betaproteobacteria exposed to metals at substantial (over 50 ppm) concentrations by studying
amoA ammonium monooxygenase marker. Cloning and sequencing approach taken by those authors led to identification of large phylogenetic clusters of organisms responsible for ammonia oxidation in the presence of metal. However, no information was gained on the denitrifying community in this or other studies. Likewise, analyses of phospholipid-linked fatty acids commonly used in microbial community analysis (see, e.g., [
33,
34]) provides no insight into effects upon phylogenetically incoherent physiological groups of microbes.
There are two basic strategies for a microbe to function in metal-contaminated environment. One, a system of transmembrane metal pumps has evolved in a number of bacteria, for example, system encoded by sil and mer operons, conferring resistance to silver and mercury, respectively. Those pumps scavenge metals on the inside of the cell membrane and remove them from the cell, thus protecting the internal cell structures from toxic metal effects.
Heavy metal resistance by means of metal ion scavenging and removal, such as provided by the classic
mer[
27,
36] or
sil[
11] operons may not necessarily provide a sufficient degree of protection in the denitrifying organisms. As denitrification-related enzymes are generally located within the cell membrane or periplasmic space, expelling heavy metal ions out of the cell would place them in the immediate contact with denitrification-related enzymes, thus limiting utility of such a resistance strategy.
The fact that denitrification enzymes are located on or near the outer cell surfaces further increases the vulnerability of the entire denitrification pathway to chemical disruption [42]. Recent work has suggested a direct effect of heavy metals upon extracellular enzyme activities [
1,
25]. Combined with the fact that scavenging/pumping systems are unlikely to protect the denitrification pathway from heavy metal effects (and may, in fact exacerbate the situation), it is expected that denitrification pathway would be uniquely sensitive to heavy metals. The notion of selective inhibition of denitrification steps by heavy metals has been supported by work of Holtan-Harwig et al. [
26], suggesting the potential for production of undesirable byproducts, such as nitrous oxide.
The second mechanism of microbial resistance to metals is evolution of enzyme forms resistant to metals. This resistance pathway is expected to be the predominant in the denitrifying bacteria, due to inability to use metal pumps for the reasons described above. The metal-resistant forms of enzymes present in metal-stressed denitrifying community are expected to be readily identifiable by their gene sequence and therefore their genetic signature.
Disruption of denitrification by heavy metals could lead to a number of undesirable consequences, influencing the human health at both global and local levels. Suppressed denitrification in the soil could lead to enhanced nitrogen retention and flushing, resulting in nonpoint nutrient pollution in waterways receiving overland or subsurface flow from impacted locations. Nutrient pollution, in turn, leads to eutrophication and massive algal blooms, including those of toxic algae and cyanobacteria (e.g.,
Microcystis), affecting human populations relying on surface waters for municipal, recreational or agricultural needs. Specific inhibition of nitrous oxide reductase by metal has been observed recently [
26], resulting in incomplete denitrification leading to emission of nitrous (and possibly nitric) oxides. As nitrous oxide is a potent greenhouse gas that also damages ozone layer [
10,
14, 49], denitrification disruption via metal contamination could act as a link between local metal contamination and global climate change phenomena.
In the present study, we used molecular tools to assess the effects of low (1 ppm) and high (500 to 2000 ppm) of lead upon the denitrifying microbial community, as well as to assess applicability of the classical intermediate disturbance concept to microbial communities.
Materials and Methods
Soil samples for low-level experiments have been collected well in advance and stored in the lab for approx. 24 months. The soil has not been extensively characterized; however, visually it resembled Mississippi silty clay used in the other set of experiments. Pb was added as aqueous solution of Pb nitrate in 10 mL of water; amount of Pb was calculated to supply 1 ppm of Pb metal to the soil sample before water addition. The control sample received 10 mL of distilled water with no Pb. Samples were sealed and incubated at ambient temperature (18–24°C) for 18 months.
High-Pb experiments were conducted in a mixture of pulverized Delta topsoil (Yazoo silty clay) and peat humus (2:1 volumetric proportions). Predetermined amounts of soils were artificially contaminated with appropriate amounts of lead nitrate to yield the desired concentrations (0, 500, 1,000 and 2,000 mg Pb/kg dry soil) and incubated in the greenhouse.
At the end of the experiments, soil subsamples of ca. 0.5 gram were collected and DNA recovered using bead beating followed by modified phenol-chloroform procedure [
20]. Crude DNA extracts were purified using GeneClean kit from Stratagene (La Jolla, CA) according to manufacturer’s instructions.
Two markers were selected for amplification in this study. Short (∼200 bp) 16S rRNA gene fragment was recovered with a P2-P3 primer set [
35]. A ca. 200 bp fragment of
nirK, a marker for NirK copper-containing nitrous oxide reductase was amplified by use of nirK1fnirK3r primer pair [
5]. Forward primer nirK1f was modified by attaching a GC-tail at the 5′ end in the manner similar to the P2 primer above. Standard PCR mixture [43] was used to amplify the fragments, with 5 minutes intial melting time at 95°C, followed by 35 cycles of 30 seconds melt at 95°C, 30 seconds annealing at 45°C and 60 seconds extension at 72°C. Cycling was followed by 5 minutes of final extension at 72°C. Resulting fragments were checked for proper and uniform size by 1.2% agarose gel electrophoresis in 0.5x TAE buffer against 100 bp DNA ladder (New England Biolabs, Ipswich, MA).
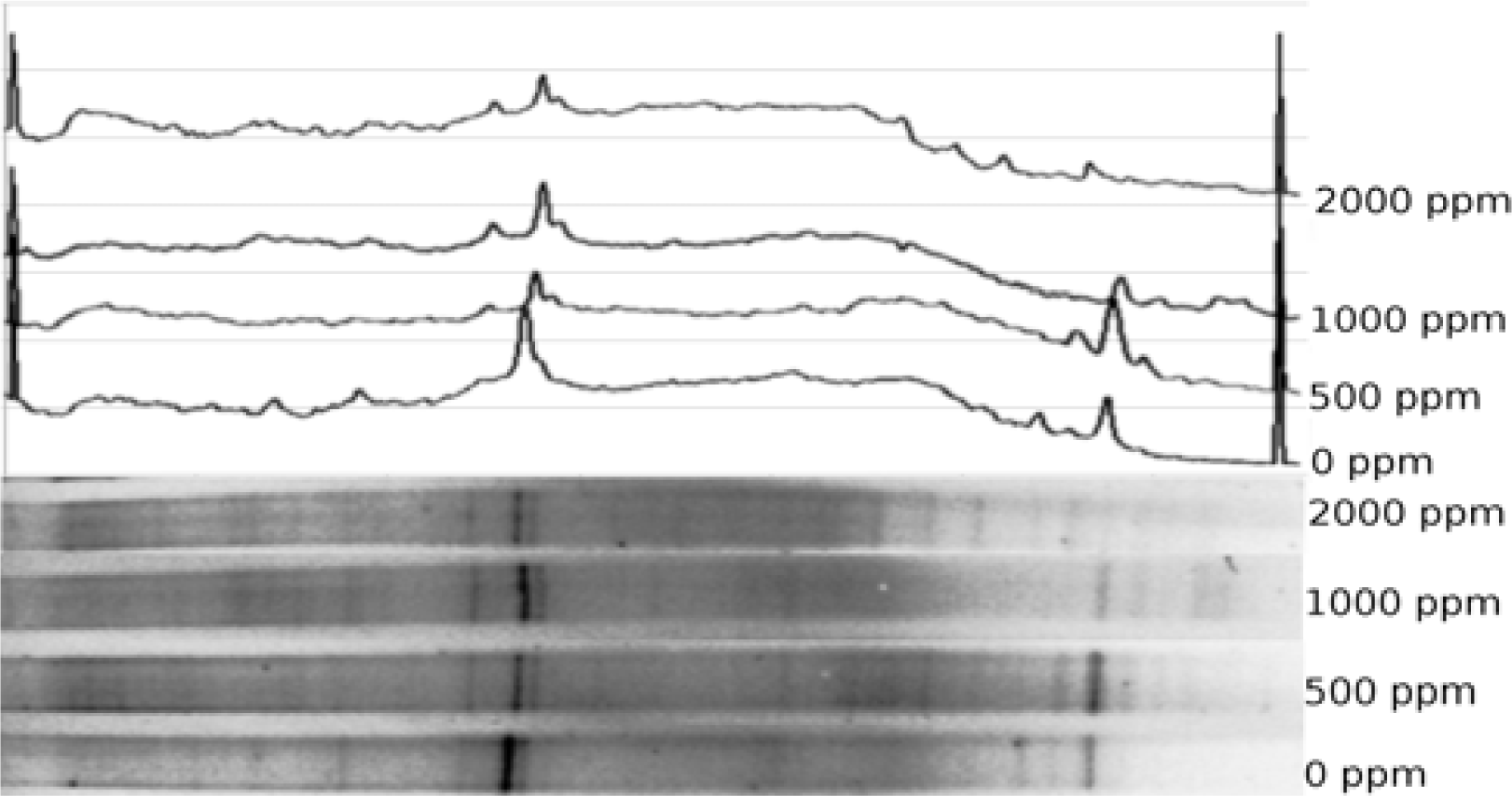
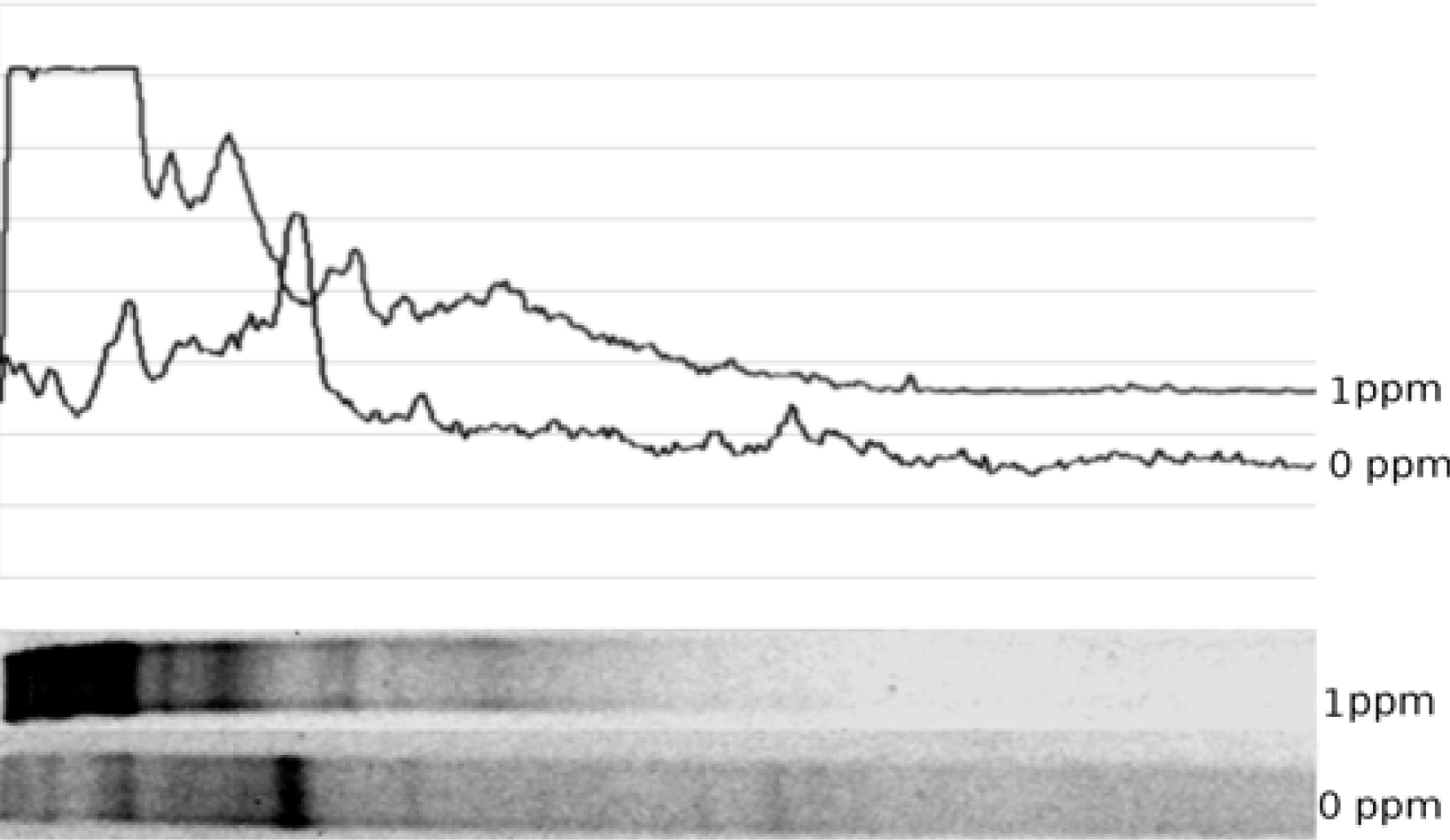
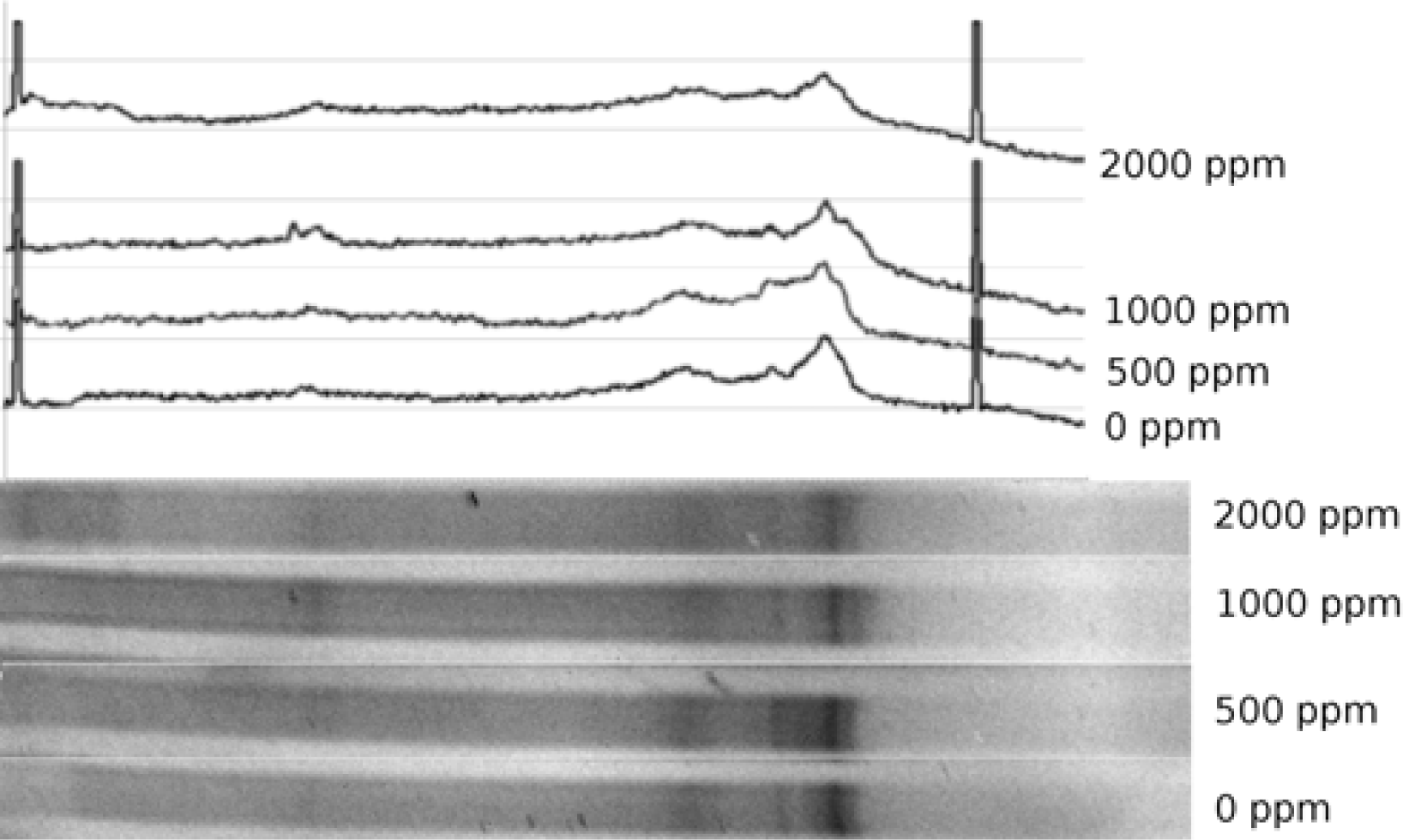
Vertical gradient DGGE gel was cast using a Northstar gradient maker coupled to a peristaltic pump. Gel concentration was set at 60% denaturant at the bottom and 20% at the top. Details on casting a similar gel are provided elsewhere [
35], except that 0.5x TAE was used both in the gel and in the electrophoresis tank. Freshly cast gel was allowed to polymerize for ca. 1 hour and the sides of the gel cassette were sealed with silicone sealant to avoid lateral lane drift. Gel was equilibrated in the buffer tank at the run temperature (70°C) for 2 hours. About 15 μL of amplified sample was loaded on the gel and voltage applied to the tank at 60V for 18 hours. Following the run, the gel was extracted from the cassette, rinsed and stained with ethidium bromide solution (1 μg/mL) for one hour, destained in distilled water for 90 minutes and photographed with UV transillumination. Diversity of the microbial community was analyzed from the recorded gel images.
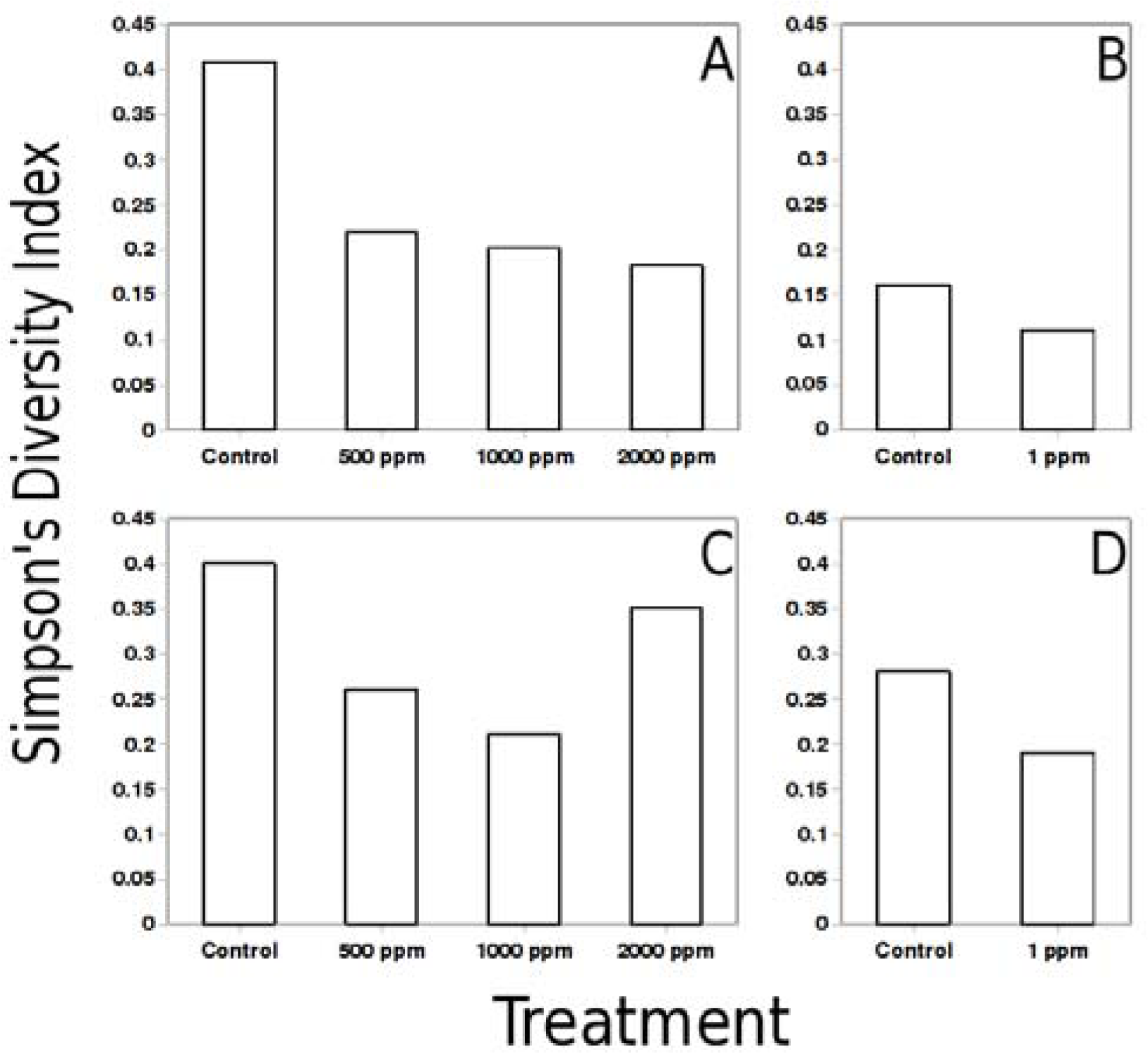
Diversity analysis was conducted by recording density profile data of a DGGE gel lane using Scion Image for Windows. Density profile was converted into a string of values and a plot generated in Open Office for Linux. Peaks recorded in the plotted profile were matched to the gel image and peak position and height measured using built-in measurement tool in GIMP for Linux. Aggregate and individual peak heights were used to compute Simpson’s diversity index [45]. Jaccard similarity index [
24] was computed on the basis of presence/absence of matched peaks in the samples.
Discussion
Heavy metals have been known to disrupt ecosystem structure and functioning for a long time. In the multicellular organisms, heavy metal target specific organs and pathways [
18,
19], resulting in disruption of definitive metabolic functions. By the same token, one can expect that some pathways in microbes would be more sensitive to metal than others, resulting in selective inhibition as metal concentration increases [
17,
18]. Selective inhibition of specific pathways would result in decline of both numbers and diversity of organisms relying on those pathways, and one could expect such shift to occur at lower metal concentrations than needed to effect community changes evidenced by the 16S profile. Such shift in a community composition could lead to profound changes in the biogeochemical processes in the environment [
26].
All metal levels produced at least some change in the community. Invariably, such change resulted in decrease of genetic diversity of the community compared to the control, regardless of the level of metal used and marker observed. Almost in every case, higher metal levels lead to lower community diversity, with one notable exception discussed below.
One of the more important observations of this study is the substantial change produced in both 16S and nirK community profiles by Pb addition at 1 ppm. Whereas change was detected by use of both those markers, change in the nirK pool was substantially more profound, as evidenced by the fact that post-exposure nirK profile differed (according to Jaccard index) from its pre-exposure counterpart to a greater extent than the 16S.This observation suggests that denitrification pathway may be the one uniquely sensitive to metal exposure. If true, disruption of denitrification could potentially enhance nitrogen retention by the soil, leading to a host of related phenomena, from increased nonpoint nutrient pollution in the nearby water bodies to enhanced plant growth and easier remediation of the metal-contaminated soils.
Exposure of soils to higher levels of Pb (500–2000 ppm) resulted in substantial community shifts. Consistently declining community diversity at higher metal levels was evidenced by both 16S and nirK marker profiles. Such a decline could be taken as the evidence of microbial population activities being substantially disrupted by the metal presence. On the other hand, modest rebound of nirK diversity at 2000 ppm Pb can not be adequately explained within this framework.
The highest degree of difference (or lowest degree of similarity) was observed between control samples and those that had lower levels of metal added. This suggests that above a certain threshold, addition of more metal has only a limited effect, a notion supported by the fact that 500 and 1000 ppm samples were the most similar ones. It appears that the further increase in metal content to 2000 ppm triggers another change in community diversity, suggesting that another threshold value was passed. The threshold-like effects of heavy metals could be explained by potential presence of several groups of organisms capable of the same type of metabolism, yet having various metal tolerances. More metal-tolerant bacteria would be outcompeted in the absence of metal, whereas presence of metal would shift the community towards more metal-tolerant guilds.
In the preliminary manner, this study demonstrates that both low (1 ppm) and high (500–2000 ppm) levels of heavy metals in soul could dramatically shift the makeup of a microbial community, almost certainly decreasing community diversity. Some classes of organisms (such as denitrification-capable bacteria) appear to be affected to a greater extent than generalized microbial community; such selective inhibition could have profound effects on community function and structure, as well as on biogeochemical phenomena in situ. It also appears that the heavy metals affect microbial communities in a threshold-like manner, suggesting the presence of sub-communities with different levels of metal tolerance.
Our data show that heavy metals have profound effect upon the general and denitrifying microbial community, as observed by others [
16,
17,
26] and as our original hypothesis suggested. In turn, denitrifying community appeared to adjust to elevated metals by selecting for putatively metal-resistant enzyme forms. Further studies, including sequencing of the recovered DNA fragments are needed to identify the changes conferring metal resistance to denitrification-related enzymes.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}