
Effects of Meteorological Factors on Waterbird Functional Diversity and Community Composition in Liaohe Estuary, China

Abstract
:1. Introduction
2. Study Area
3. Material and Methods
3.1. Data Collection
3.1.1. Waterbird Observation
3.1.2. Functional Traits
3.1.3. Meteorological Variables
3.2. Statistical Analyses
3.2.1. Functional Alpha and Beta Diversity
3.2.2. Changes in Meteorological Factors
3.2.3. Relationship between Functional Diversity and Meteorological Variables
4. Results
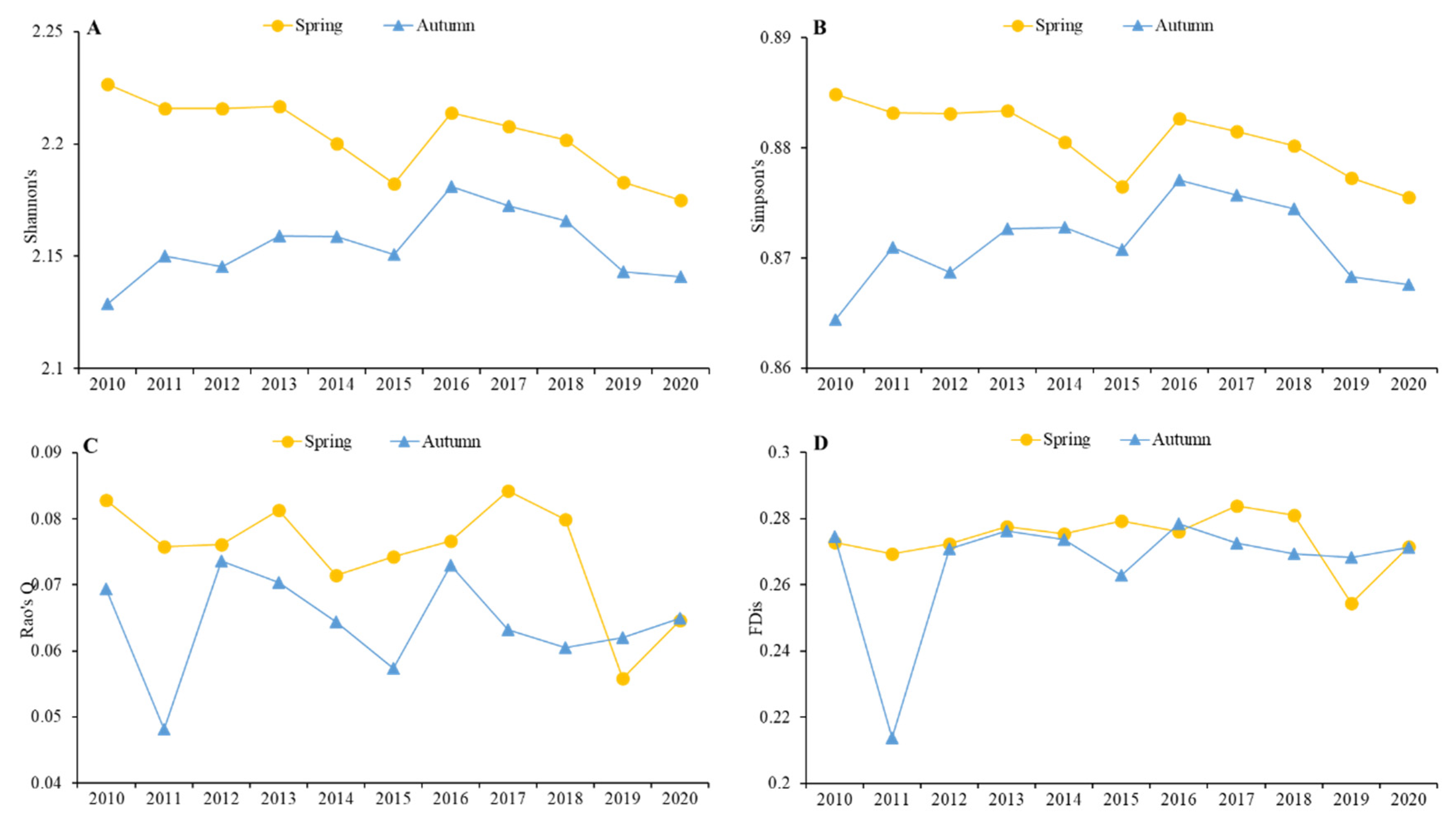
4.1. Waterbird Functional Diversity Dynamics
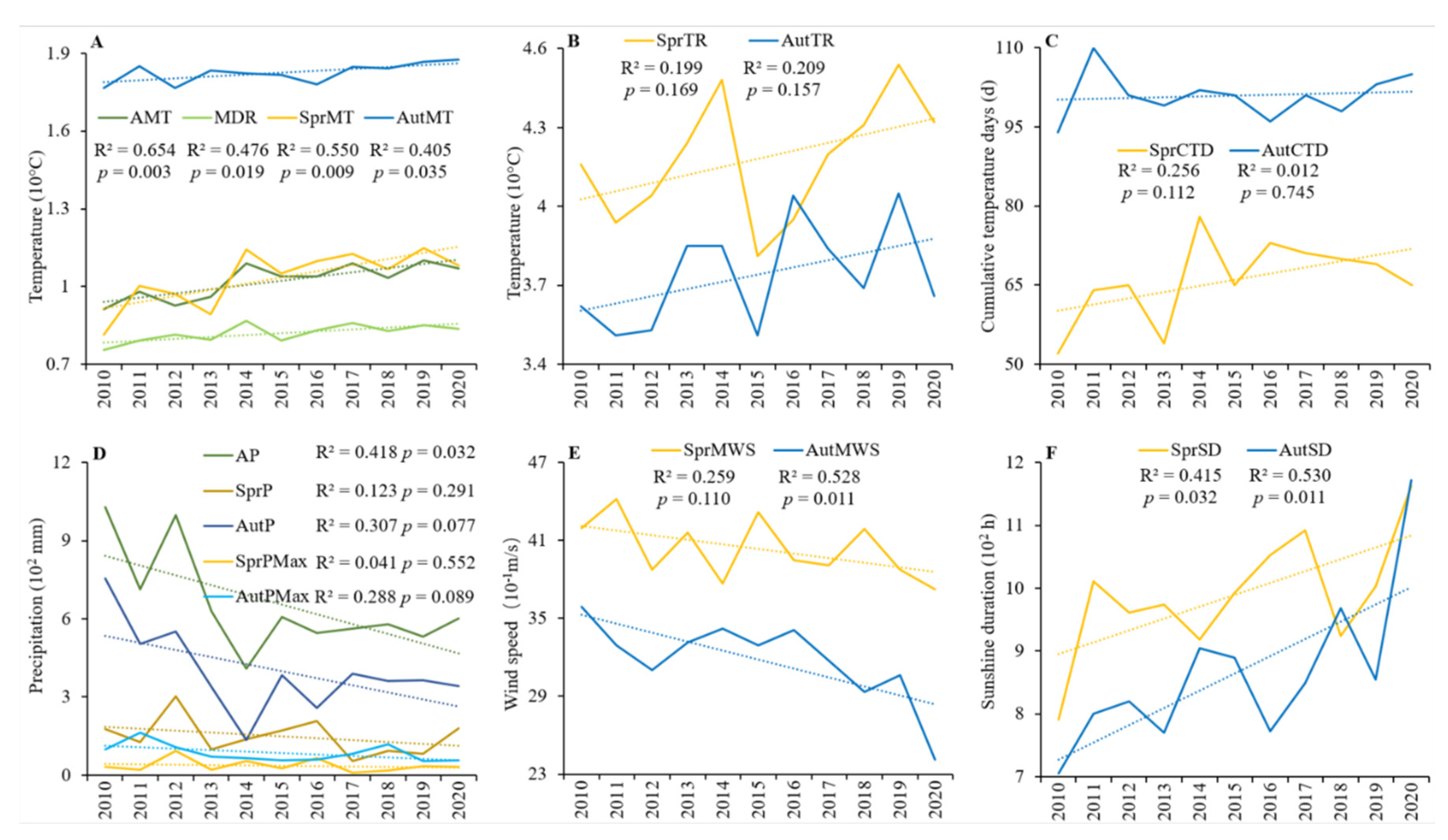
4.2. Meteorological Changes
4.3. Relationship between Functional Diversity and Meteorological Factors
5. Discussion
5.1. Trends in Functional Diversity
5.2. Responses of Waterbird Functional Diversity to Meteorological Factors
5.3. Seasonal Differences in Response to Meteorological Factors
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Name | Species | Families | Residence | Breeding | Diet | Nest Site |
|---|---|---|---|---|---|---|---|
| 1 | Little Grebe | Tachybaptus ruficollis | PA | R | B | PIS | Float |
| 2 | Great Crested Grebe | Podiceps cristatus | PA | S | B | PIS | Float |
| 3 | Great Cormorant | Phalacrocorax carbo | PA | S | B | PIS | Cliff |
| 4 | Gray Heron | Ardea cinerea | CG | R | B | PIS | Tree |
| 5 | Purple Heron | Ardea purpurea | CG | S | B | PIS | Grass |
| 6 | Great White Egret | Ardea alba | CG | S | B | PIS | Grass |
| 7 | Little Egret | Egretta garzetta | CG | S | B | PIS | Grass |
| 8 | Black-crowned Night heron | Nycticorax nycticorax | CG | S | B | PIS | Tree |
| 9 | Eurasian Spoonbill | Platalea leucorodia | CG | S | B | PIS | Grass |
| 10 | Oriental Stork | Ciconia boyciana | CG | S | B | PIS | Grass |
| 11 | Common Crane | Grus grus | CG | W | NB | OMN | Grass |
| 12 | Siberian Crane | Leucogeranus | CG | P | NB | HER | Grass |
| 13 | Red-crowned Crane | Grus japonensis | CG | R | B | OMN | Grass |
| 14 | Eurasian/common Coot | Fulica atra | CG | S | B | OMN | Grass |
| 15 | Whooper Swan | Cygnus cygnus | PA | S | B | HER | Grass |
| 16 | Greater White-fronted Goose | Anser albifrons | PA | P | NB | HER | Shrub |
| 17 | Swan Goose | Anser cygnoid | PA | W | NB | HER | Grass |
| 18 | Bean Goose | Anser fabalis | PA | P | NB | HER | Shrub |
| 19 | Common Shelduck | Tadorna tadorna | PA | S | B | OMN | Tree |
| 20 | Mallard | Anas platyrhynchos | PA | R | B | OMN | Grass |
| 21 | Chinese Spot-billed Duck | Anas zonorhyncha | PA | W | NB | OMN | Grass |
| 22 | Northern Shoveler | Spatula clypeata | PA | P | NB | OMN | Grass |
| 23 | Gadwall | Mareca strepera | PA | S | B | HER | Shrub |
| 24 | Ruddy Shelduck | Tadorna ferruginea | PA | S | B | HER | Tree |
| 25 | Eurasian Wigeon | Mareca penelope | PA | S | B | HER | Shrub |
| 26 | Common Goldeneye | Bucephala clangula | PA | P | NB | PIS | Tree |
| 27 | Falcated Duck | Mareca falcata | PA | S | B | HER | Shrub |
| 28 | Garganey | Spatula querquedula | PA | S | B | HER | Grass |
| 29 | Common Teal | Anas crecca | PA | S | B | HER | Grass |
| 30 | Baikal Teal | Anas formosa | PA | P | NB | HER | Shrub |
| 31 | Tufted Duck | Aythya fuligula | PA | P | NB | OMN | Shrub |
| 32 | Common Pochard | Aythya ferina | PA | P | NB | OMN | Float |
| 33 | Goosander | Mergus merganser | PA | S | B | PIS | Tree |
| 34 | Smew | Mergellus albellus | PA | W | NB | PIS | Tree |
| 35 | Northern Pintail | Anas acuta | PA | P | NB | HER | Grass |
| 36 | Northern Lapwing | Vanellus vanellus | Cha | S | B | OMN | Tree |
| 37 | Pacific Golden Plover | Pluvialis fulva | Cha | P | NB | INS | Bare |
| 38 | Grey Plover | Pluvialis squatarola | Cha | P | NB | BEN | Bare |
| 39 | Lesser Sand Plover | Charadrius mongolus | Cha | P | NB | BEN | Bare |
| 40 | Kentish Plover | Charadrius alexandrinus | Cha | S | B | BEN | Bare |
| 41 | Eurasian Oystercatcher | Haematopus ostralegus | Cha | S | B | BEN | Bare |
| 42 | Black-winged Stilt | Himantopus himantopus | Cha | S | B | BEN | Grass |
| 43 | Pied Avocet | Recurvirostra avosetta | Cha | S | B | BEN | Bare |
| 44 | Far Eastern Curlew | Numenius madagascariensis | Cha | P | NB | CAR | Bare |
| 45 | Whimbrel | Numenius phaeopus | Cha | P | NB | BEN | Grass |
| 46 | Little Curlew | Numenius minutus | Cha | P | NB | OMN | Bare |
| 47 | Great Knot | Calidris tenuirostris | Cha | P | NB | BEN | Shrub |
| 48 | Sharp-tailed Sandpiper | Calidris acuminata | Cha | P | NB | OMN | Shrub |
| 49 | Marsh Sandpiper | Tringa stagnatilis | Cha | P | NB | BEN | Grass |
| 50 | Spotted Redshank | Tringa erythropus | Cha | P | NB | BEN | Grass |
| 51 | Wood Sandpiper | Tringa glareola | Cha | P | NB | OMN | Bare |
| 52 | Eurasian Curlew | Numenius arquata | Cha | P | NB | BEN | Bare |
| 53 | Bar-tailed Godwit | Limosa lapponica | Cha | P | NB | BEN | Bare |
| 54 | Black-tailed Godwit | Limosa limosa | Cha | S | B | BEN | Shrub |
| 55 | Common Redshank | Tringa totanus | Cha | S | B | BEN | Grass |
| 56 | Common Greenshank | Tringa nebularia | Cha | P | NB | BEN | Bare |
| 57 | Terek Sandpiper | Xenus cinereus | Cha | P | NB | BEN | Bare |
| 58 | Red-necked Stint | Calidris ruficollis | Cha | P | NB | BEN | Grass |
| 59 | Red Knot | Calidris canutus | Cha | P | NB | BEN | Grass |
| 60 | Dunlin | Calidris alpina | Cha | P | NB | BEN | Grass |
| 61 | Arctic/American Herring Gull | Larus smithsonianus | Lar | P | NB | CAR | Cliff |
| 62 | Slaty-backed Gull | Larus schistisagus | Lar | W | NB | CAR | Cliff |
| 63 | Black-tailed Gull | Larus crassirostris | Lar | S | B | CAR | Cliff |
| 64 | Pallas’s Gull | Ichthyaetus ichthyaetus | Lar | P | NB | CAR | Cliff |
| 65 | Mew Gull | Larus canus | Lar | W | NB | PIS | Grass |
| 66 | Relict Gull | Larus relictus | Lar | P | NB | PIS | Bare |
| 67 | Caspian Tern | Hydroprogne caspia | Lar | S | B | CAR | Bare |
| 68 | White-winged Tern | Chlidonias leucopterus | Lar | S | B | PIS | Float |
| 69 | Black-headed Gull | Larus ridibundus | Lar | S | B | CAR | Grass |
| 70 | Saunders’s Gull | Larus saundersi | Lar | S | B | INS | Bare |
| 71 | Whiskered Tern | Chlidonias hybrida | Lar | S | B | PIS | Float |
| 72 | Gull-billed Tern | Gelochelidon nilotica | Lar | S | B | PIS | Bare |
| 73 | Little Tern | Sternula albifrons | Lar | S | B | PIS | Bare |
| 74 | Common Tern | Sterna hirundo | Lar | S | B | CAR | Grass |
Appendix B
| Years | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | Mean |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Spring | 0.25 | 0.07 | 0.1 | 0.08 | 0.04 | 0.04 | 0.10 | 0.10 | 0.07 | 0.06 | 0.09 |
| Autumn | 0.18 | 0.12 | 0.17 | 0.06 | 0.03 | 0.06 | 0.09 | 0.05 | 0.04 | 0.02 | 0.08 |
| Inter Years | 10–11 | 11–12 | 12–13 | 13–14 | 14–15 | 15–16 | 16–17 | 17–18 | 18–19 | 19–20 | Mean |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Spr.–Spr. | 0.21 | 0.05 | 0.03 | 0.07 | 0.03 | 0.07 | 0.02 | 0.04 | 0.12 | 0.06 | 0.07 |
| Aut.–Aut. | 0.11 | 0.11 | 0.13 | 0.06 | 0.09 | 0.05 | 0.05 | 0.03 | 0.07 | 0.02 | 0.07 |
| Aut.–Spr. | 0.11 | 0.09 | 0.08 | 0.05 | 0.08 | 0.18 | 0.07 | 0.10 | 0.03 | 0.07 | 0.09 |
| Years | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| βsor | 0.19 | 0.09 | 0.08 | 0.08 | 0.07 | 0.08 | 0.18 | 0.07 | 0.11 | 0.03 | 0.07 | 0.10 |
References
- Mantyka-Pringle, C.S.; Visconti, P.; Di Marco, M.; Martin, T.; Rondinini, C.; Rhodes, J. Climate change modifies risk of global biodiversity loss due to land-cover change. Biol. Conserv. 2015, 187, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Opdam, P.; Wascher, D. Climate change meets habitat fragmentation: Linking landscape and biogeographical scale levels in research and conservation. Biol. Conserv. 2004, 117, 285–297. [Google Scholar] [CrossRef]
- Bibby, C.J. Making the most of birds as environmental indicators. Ostrich 1999, 70, 81–88. [Google Scholar] [CrossRef]
- Stanley, T.A.; John, W.A. Bird populations and environmental changes: Can birds be bio-indicators. Am. Birds 1989, 2, 260–270. [Google Scholar]
- Naeem, S.; Duffy, J.E.; Zavaleta, E. The Functions of Biological Diversity in an Age of Extinction. Science 2012, 336, 1401–1406. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D. Functional diversity. Enclopedia of Biodiversity; Academic Press: Cambridge, MA, USA, 2001; Volume 3, pp. 109–121. [Google Scholar]
- Boersma, K.S.; Dee, L.E.; Miller, S.J.; Bogan, M.T.; Lytle, D.A.; Gitelman, A.I. Linking multidimensional functional diversity to quantitative methods: A graphical hypothesise—Evaluation framework. Ecology 2016, 97, 583–593. [Google Scholar] [CrossRef]
- Almeida, B.D.A.; Sebastian-Gonzalez, E.; Anjos, L.D.; Green, A.J.; Botella, F. A functional perspective for breeding and wintering waterbird communities: Temporal trends in species and trait diversity. OIKOS 2019, 128, 1103–1115. [Google Scholar] [CrossRef]
- Daneshvar, F.; Nejadhashemi, A.P.; Herman, M.R.; Abouali, M. Response of benthic macroinvertebrate communities to climate change. Ecohydrol. Hydrobiol. 2017, 17, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Che, X.; Chen, D.; Zhang, M.; Quan, Q.; Møller, A.P.; Zou, F. Seasonal dynamics of waterbird assembly mechanisms revealed by patterns in phylogenetic and functional diversity in a subtropical wetland. BioTropica 2019, 51, 421–431. [Google Scholar] [CrossRef]
- Austin, G.E.; Rehfisch, M.M. Shifting nonbreeding distributions of migratory fauna in relation to climatic change. Glob. Chang. Biol. 2005, 11, 31–38. [Google Scholar] [CrossRef]
- Barnagaud, J.-Y.; Gaüzère, P.; Zuckerberg, B.; Princé, K.; Svenning, J.-C. Temporal changes in bird functional diversity across the United States. Oecologia 2017, 185, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Natalia, M.S.; Patricia, K. Do taxonomic, phylogenetic and functional plant α-and β-diversity reflect environmental patterns in the Lower Paraná River floodplain. Plant Ecol. Divers. 2017, 10, 153–165. [Google Scholar]
- Mendez, V.; Gill, J.A.; Burton, N.H.K.; Austin, G.E.; Petchey, O.L.; Davies, R.G. Functional diversity across space and time: Trends in wader communities on British estuaries. Divers. Distrib. 2011, 18, 356–365. [Google Scholar] [CrossRef]
- Day, J.W.; Christian, R.R.; Boesch, D.; Yáñez-Arancibia, A.; Morris, J.; Twilley, R.R.; Naylor, L.; Schaffner, L.; Stevenson, C. Consequences of Climate Change on the Ecogeomorphology of Coastal Wetlands. Estuaries Coasts 2008, 31, 477–491. [Google Scholar] [CrossRef]
- Rogers, K.; Saintilan, N.; Woodroffe, C.D. Surface elevation change and vegetation distribution dynamics in a subtropical coastal wetland: Implications for coastal wetland response to climate change. Estuar. Coast. Shelf Sci. 2014, 149, 46–56. [Google Scholar] [CrossRef]
- Brix, H.; Ye, S.; Laws, E.A.; Sun, D.; Li, G.; Ding, X.; Yuan, H.; Zhao, G.; Wang, J.; Pei, S. Large-scale management of common reed, Phragmites australis, for paper production: A case study from the Liaohe Delta, China. Ecol. Eng. 2014, 73, 760–769. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, G.; New, T. Abiotic Factors Influencing the Distribution of Vegetation in Coastal Estuary of the Liaohe Delta, Northeast China. Estuaries Coasts 2009, 32, 937–942. [Google Scholar] [CrossRef]
- Li, X.; Christopher, J.A.; Wang, Y.; Lei, G. Waterbird diversity and abundance in response to variations in climate in the Liaohe Estuary, China. Ecol. Indic. 2021, 132, 108286. [Google Scholar] [CrossRef]
- Zhu, L.; Wu, J.; Xu, Y.; Hu, R.; Wang, N. Recent geomorphic changes in the Liaohe Estuary. J. Geogr. Sci. 2010, 20, 31–48. [Google Scholar] [CrossRef]
- Du, J.; Song, K. Validation of Global Evapotranspiration Product (MOD16) Using Flux Tower Data from Panjin Coastal Wetland, Northeast China. Chin. Geogr. Sci. 2018, 28, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, L.; Su, F.; Wang, T.; Gao, P. The Periodic Response of Tidal Flat Sediments to Runoff Variation of Upstream Main River: A Case Study in the Liaohe Estuary Wetland, China. Water 2019, 12, 61. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Baoshu, Y.; Yanqing, X.; Dezhou, Y. Numerical simulation of tides and tidal currents in Liaodong Bay with POM. Prog. Nat. Sci. 2005, 15, 47–55. [Google Scholar] [CrossRef]
- Yan, X.; Hu, Y.; Chang, Y.; Zhang, D.; Liu, M.; Guo, J.; Ren, B. Monitoring Wetland Changes Both Outside and Inside Reclamation Areas for Coastal Management of the Northern Liaodong Bay, China. Wetlands 2017, 37, 885–897. [Google Scholar] [CrossRef]
- Li, D.; Chen, S.; Guan, L.; Lloyd, H.; Liu, Y.; Lv, J.; Zhang, Z. Patterns of waterbird community composition across a natural and restored wetland landscape mosaic, Yellow River Delta, China. Estuar. Coast. Shelf Sci. 2011, 91, 325–332. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Dendrograms and measuring functional diversity. OIKOS 2007, 116, 1422–1426. [Google Scholar] [CrossRef]
- Luck, G.W.; Carter, A.; Smallbone, L. Changes in Bird Functional Diversity across Multiple Land Uses: Interpretations of Functional Redundancy Depend on Functional Group Identity. PLoS ONE 2013, 8, e63671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blendinger, P.G.; Villegas, M. Crop size is more important than neighborhood fruit availability for fruit removal of Eugenia uniflora (Myrtaceae) by bird seed dispersers. Plant Ecol. 2011, 212, 889–899. [Google Scholar] [CrossRef]
- Şekercioğlu Çağan, H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, W.; Zeng, Q.; Tian, H.; Jia, Y.; Lei, G.; Wen, L. Lake productivity and waterbird functional diversity across geographic and environmental gradients in temperate China. Ecol. Evol. 2020, 10, 11237–11250. [Google Scholar] [CrossRef]
- Che, X.; Zhang, M.; Zhao, Y.; Zhang, Q.; Quan, Q.; Møller, A.; Zou, F. Phylogenetic and Functional Structure of Wintering Waterbird Communities Associated with Ecological Differences. Sci. Rep. 2018, 8, 1232. [Google Scholar] [CrossRef] [Green Version]
- Eric, G.; Jacques, C.; Georges, B.; Marie-Laure, N.; Catherine, R.; Max, D.; Laurent, G.; Alain, B.; David, A.; Astrid, B.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Gorelick, R. Combining richness and abundance into a single diversity index using matrix analogues of Shannon’s and Simpson’s indices. Ecography 2006, 29, 525–530. [Google Scholar] [CrossRef]
- Botta-Dukat, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Baselga, A. Multiple site dissimilarity quantifies compositional heterogeneity among several sites, while average pairwise dissimilarity may be misleading. Ecography 2013, 36, 124–128. [Google Scholar] [CrossRef]
- Lichtenberg, J.M.; Şimşek, Ö. Simple Regression Models. Proc. Mach. Learn. Res. 2017, 58, 13–25. [Google Scholar]
- Brown, A.M.; Warton, D.; Andrew, N.R.; Binns, M.; Cassis, G.; Gibb, H. The fourth-corner solution—Using predictive models to understand how species traits interact with the environment. Methods Ecol. Evol. 2014, 5, 344–352. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the Species Traits–Environment Relationships: The Fourth-Corner Problem Revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef]
- Studds, C.E.; Kendall, B.E.; Murray, N.J.; Straw, P.; Woehler, E.J.; Fuller, R.A. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 2017, 8, 14895. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.; Villéger, S.; Mason, N.W.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, F.; Silbernagel, J.; Larson, H. Community structure, abundance variation and population trends of waterbirds in relation to water level fluctuation in Poyang Lake. J. Great Lakes Res. 2019, 45, 976–985. [Google Scholar] [CrossRef]
- Götmark, F.; Blomqvist, D.; Johansson, O.C.; Bergkvist, J. Nest Site Selection: A Trade-Off between Concealment and View of the Surroundings? J. Avian Biol. 1995, 26, 305. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Xin, Y.-Z.; Gao, D.-M.; Li, F.-M.; Morgan, J.; Xing, B.-S. Microbial Community Characteristics in a Degraded Wetland of the Yellow River Delta. Pedosphere 2010, 20, 466–478. [Google Scholar] [CrossRef]
- Fan, J.; Xu, Y.; Ge, H.; Yang, W. Vegetation growth variation in relation to topography in Horqin Sandy Land. Ecol. Indic. 2020, 113, 106215. [Google Scholar] [CrossRef]
- Hua, Y.; Cui, B.; He, W. Changes in Water Birds Habitat Suitability Following Wetland Restoration in the Yellow River Delta, China. CLEAN—Soil Air Water 2012, 40, 1076–1084. [Google Scholar] [CrossRef]
- Raposa, K.B.; McKinney, R.A.; Beaudette, A. Effects of Tide Stage on the Use of Salt Marshes by Wading Birds in Rhode Island. Northeast. Nat. 2009, 16, 209–224. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Gaston, K.J.; Loder, N. Geographic gradients in body size: A clarification of Bergmann’s rule. Divers. Distrib. 1999, 5, 165–174. [Google Scholar] [CrossRef]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.J.; Sabatier, P. Bird Migration Routes and Risk for Pathogen Dispersion into Western Mediterranean Wetlands. Emerg. Infect. Dis. 2007, 13, 365–372. [Google Scholar] [CrossRef]
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenstroöm, J.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Global Patterns of Influenza A Virus in Wild Birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Rogers, K.; Saintilan, N.; Ling, J. The influences of climate and hydrology on population dynamics of waterbirds in the lower Murrumbidgee River floodplains in Southeast Australia: Implications for environmental water management. Ecol. Model. 2011, 222, 154–163. [Google Scholar] [CrossRef]
- Saino, N.; Romano, M.; Ambrosini, R.; Ferrari, R.P.; Moller, A.P. Timing of reproduction and egg quality covary with temperature in the insectivorous Barn Swallow, Hirundo rustica. Funct. Ecol. 2004, 18, 50–57. [Google Scholar] [CrossRef]
- Arbeiter, S.; Schulze, M.; Tamm, P.; Hahn, S. Strong cascading effect of weather conditions on prey availability and annual breeding performance in European bee-eaters Merops apiaster. J. Ornithol. 2015, 157, 155–163. [Google Scholar] [CrossRef]
- Li, X.; Lei, G.; Li, Y.; Wang, Y.; Tan, Z. Assessing hydrodynamic effects of ecological restoration scenarios for a tidal-dominated wetland in Liaodong Bay (China). Sci. Total Environ. 2021, 752, 142339. [Google Scholar] [CrossRef] [PubMed]





| Categories | Type | Trait | Description | Source |
|---|---|---|---|---|
| Categories | C | Families | Divided into four groups according to their families and habits | b and c |
| C | Residence | Four groups: summer visitor, winter visitor, passage, and resident | c and d | |
| C | Breeding | Two groups: breeding and non-breeding | b, c, and d | |
| C | Diet | Six groups: herbivore, insectivore, carnivore, piscivore, benthivore, and omnivore | b | |
| C | Nest sites | Five groups: bare ground, cliff, float, grass or shrub, and tree | a and b | |
| Body | Q | Mass (g) | a and b | |
| Q | Size (mm) | a and b | ||
| Flying ability | Q | Wingspan (mm) | a and b | |
| Q | Wing length (mm) | a and b | ||
| Q | Tail length (mm) | b | ||
| Foraging ability | Q | Bill length (mm) | b | |
| Q | Tarsus length (mm) | b | ||
| Reproduction | Q | Incubation (day) | a and b | |
| Q | Clutch (number) | a and b | ||
| Q | Nest size (mm) | a and b |
| Traits | Meteorological Variables | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AMT | MDR | SMT | STR | SCTD | AP | SP | SPMax | SMWS | SSD | ||
| (a) Spring | |||||||||||
| Families | *** | ** | *** | ° | ** | *** | ° | ° | ° | * | |
| PA | −−− | −− | −−− | ° | −−− | +++ | ° | ° | ° | − | |
| CG | ° | ° | − | ° | − | − | ° | ° | ° | ° | |
| Cha | +++ | +++ | +++ | ° | +++ | −−− | ° | ° | ° | +++ | |
| Lar | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Residence | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| S | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| W | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| P | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| R | − | − | ° | − | ° | + | ° | ° | ° | ° | |
| Diet | * | ° | ** | ° | ** | ** | ° | ° | ° | * | |
| Herbivores | −−− | − | −−− | ° | −− | +++ | ° | ° | ° | ° | |
| Insectivores | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Carnivores | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Piscivores | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Benthivores | ++ | +++ | +++ | ° | +++ | −−− | ° | ° | ° | −−− | |
| Omnivores | ° | − | ° | ° | ° | ° | ° | ° | ° | ° | |
| Nest sites | ** | ** | *** | ° | *** | ** | ° | ° | ° | * | |
| Bare ground | +++ | ++ | +++ | ° | +++ | --- | ° | ° | ° | ++ | |
| Cliff | − | − | −− | ° | −− | ° | ° | + | ° | −− | |
| Float | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Grass/shrub | −− | − | −− | ° | −− | ++ | ° | ° | ° | ° | |
| Tree | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Body | Mass | −−− | −−− | −−− | ° | −−− | +++ | ° | ° | + | −− |
| Size | −−− | −− | −−− | ° | −−− | +++ | ° | ° | ° | ||
| Flying | Wingspan | − | −− | −− | ° | −− | ++ | ° | ° | ° | −− |
| Wing length | − | −− | −− | ° | −− | ++ | ° | ° | ° | −− | |
| Tail length | −− | −− | −− | ° | −−− | ++ | ° | ° | ° | −− | |
| Foraging | Bill length | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° |
| Tarsus length | − | − | − | ° | − | + | ° | ° | ° | ° | |
| Reproduction | Incubation | −−− | −−− | −−− | ° | −−− | +++ | ° | ° | ° | − |
| Clutch | − | ° | −− | ° | −− | + | ° | ° | ° | ° | |
| Nest size | −−− | −−− | −−− | ° | −− | +++ | ° | ° | ° | −− | |
| (b) Autumn | |||||||||||
| Families | *** | ** | * | ° | ° | *** | *** | ° | ° | ° | |
| PA | −−− | −−− | −− | − | ° | +++ | +++ | ++ | ° | − | |
| CG | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Cha | ++ | + | ° | ° | ° | −− | −− | − | ° | ° | |
| Lar | + | ° | + | ° | ° | −− | −− | ° | ° | ° | |
| Residence | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| S | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| W | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| P | ° | ° | ° | ° | ° | ° | ° | - | ° | ° | |
| R | ° | ° | ° | ° | ° | ° | ° | + | ° | ° | |
| Diet | *** | *** | ** | ° | ° | *** | *** | ** | ° | ° | |
| Herbivores | −−− | −−− | −−− | ° | − | +++ | ++ | ° | + | ++ | |
| Insectivores | ° | ° | ° | ° | + | ° | ° | + | ° | ° | |
| Carnivores | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Piscivores | ++ | ++ | + | ° | ° | −− | − | ° | ° | ° | |
| Benthivores | ++ | + | ° | ° | ° | −− | − | −−− | ° | + | |
| Omnivores | − | − | ° | − | ° | ++ | ++ | ++ | ° | ° | |
| Nest sites | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Bare ground | + | ° | ° | ° | ° | − | − | ° | ° | ° | |
| Cliff | ° | ° | ° | ° | + | ° | ° | + | ° | ° | |
| Float | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Grass/shrub | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Tree | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Body | Mass | −−− | − | −− | ° | ° | ++ | +++ | ++ | ° | − |
| Size | −− | − | ° | ° | ° | ++ | ++ | + | ° | ° | |
| Flying | Wingspan | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° |
| Wing length | ° | ° | ° | ° | ° | + | ° | ° | ° | ° | |
| Tail length | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
| Foraging | Bill length | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° |
| Tarsus length | ° | ° | ° | ° | ° | + | + | ° | ° | ° | |
| Reproduction | Incubation | − | ° | ° | ° | ° | ++ | ++ | + | ° | ° |
| Clutch | −−− | −− | −− | −− | ° | +++ | +++ | ° | ° | ° | |
| Nest size | ° | ° | ° | ° | ° | ° | ° | ° | ° | ° | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zeng, Q.; Lei, G.; Sun, G. Effects of Meteorological Factors on Waterbird Functional Diversity and Community Composition in Liaohe Estuary, China. Int. J. Environ. Res. Public Health 2022, 19, 5392. https://doi.org/10.3390/ijerph19095392
Li X, Zeng Q, Lei G, Sun G. Effects of Meteorological Factors on Waterbird Functional Diversity and Community Composition in Liaohe Estuary, China. International Journal of Environmental Research and Public Health. 2022; 19(9):5392. https://doi.org/10.3390/ijerph19095392
Chicago/Turabian StyleLi, Xiuzhong, Qing Zeng, Guangchun Lei, and Gongqi Sun. 2022. "Effects of Meteorological Factors on Waterbird Functional Diversity and Community Composition in Liaohe Estuary, China" International Journal of Environmental Research and Public Health 19, no. 9: 5392. https://doi.org/10.3390/ijerph19095392