
Untargeted Metabolomics Profiling of Bioactive Compounds under Varying Digestate Storage Conditions: Assessment of Antioxidant and Antifungal Activity


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material
2.2. Experimental Set-Up
2.3. Antioxidant Activity of Digestate
2.4. Antifungal Activity of Digestate
2.5. Analytical Methods
2.5.1. Sample Preparation for GC–MS
2.5.2. GC–MS Analysis
2.5.3. Data Processing and Statistical Analysis
2.5.4. Pathway Analysis
3. Results and Discussion
3.1. Digestate Storage under Varying Conditions Changes Its Metabolites Profile
3.2. Digestate Storage under Varying Conditions Generates Different Types of Metabolites
3.3. Changes in the Metabolite’s Derivative Pathways under Varying Digestate Storage Conditions
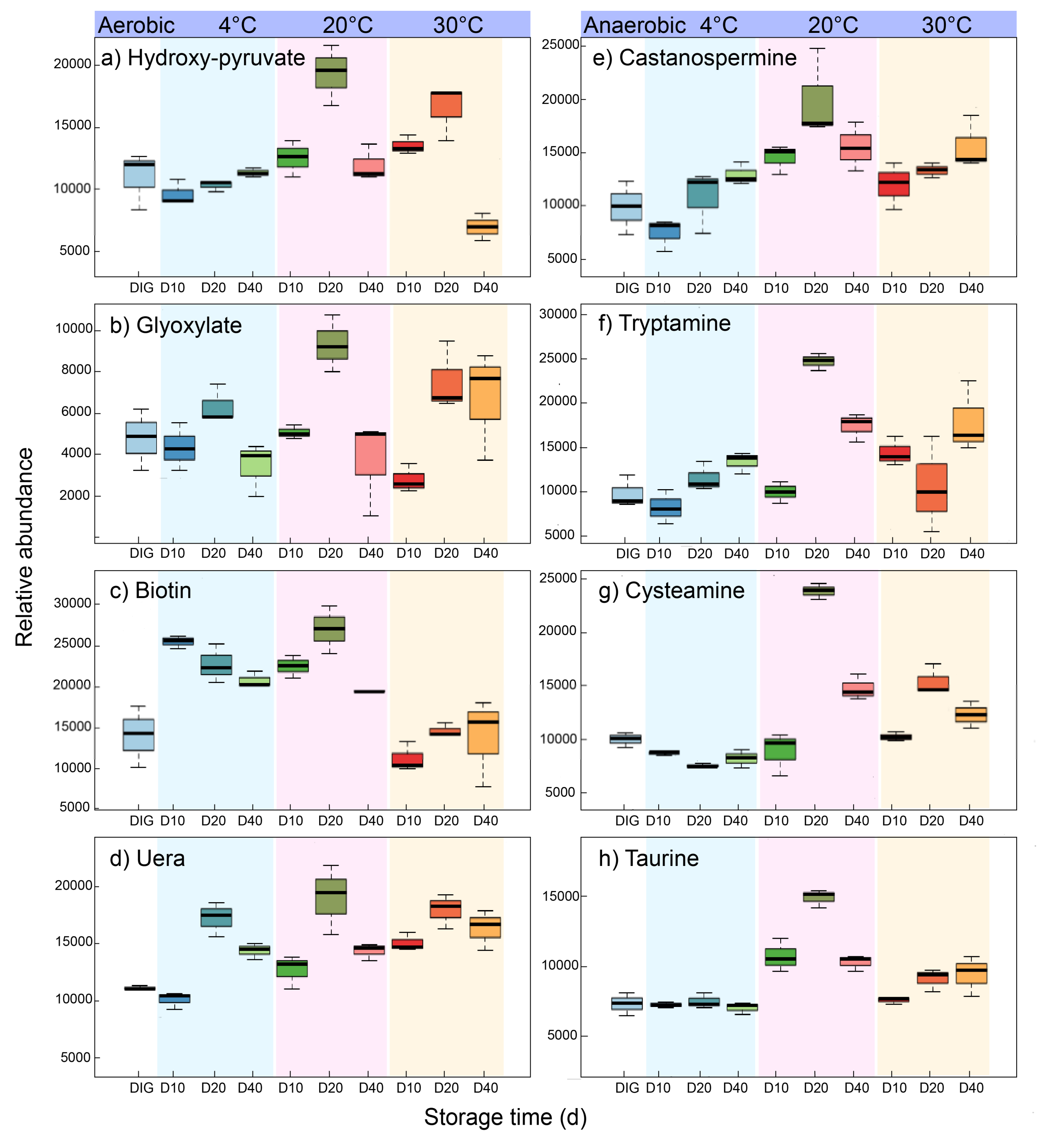
3.3.1. Aerobic Storage of Digestate Promotes the Biotin Metabolic Pathway for Vitamin Production
3.3.2. The Aerobic Storage Mode Promoted Glycine, Serine, and Threonine Metabolism to Enhance the Antimicrobial Capacity
3.3.3. Anaerobic Storage Enhances the Tryptophan and Lysine Metabolic Pathway to Produce Indole Alkaloids
3.3.4. Anaerobic Storage Promotes the Taurine and Hypotaurine Metabolism Pathways to Produce Antioxidants
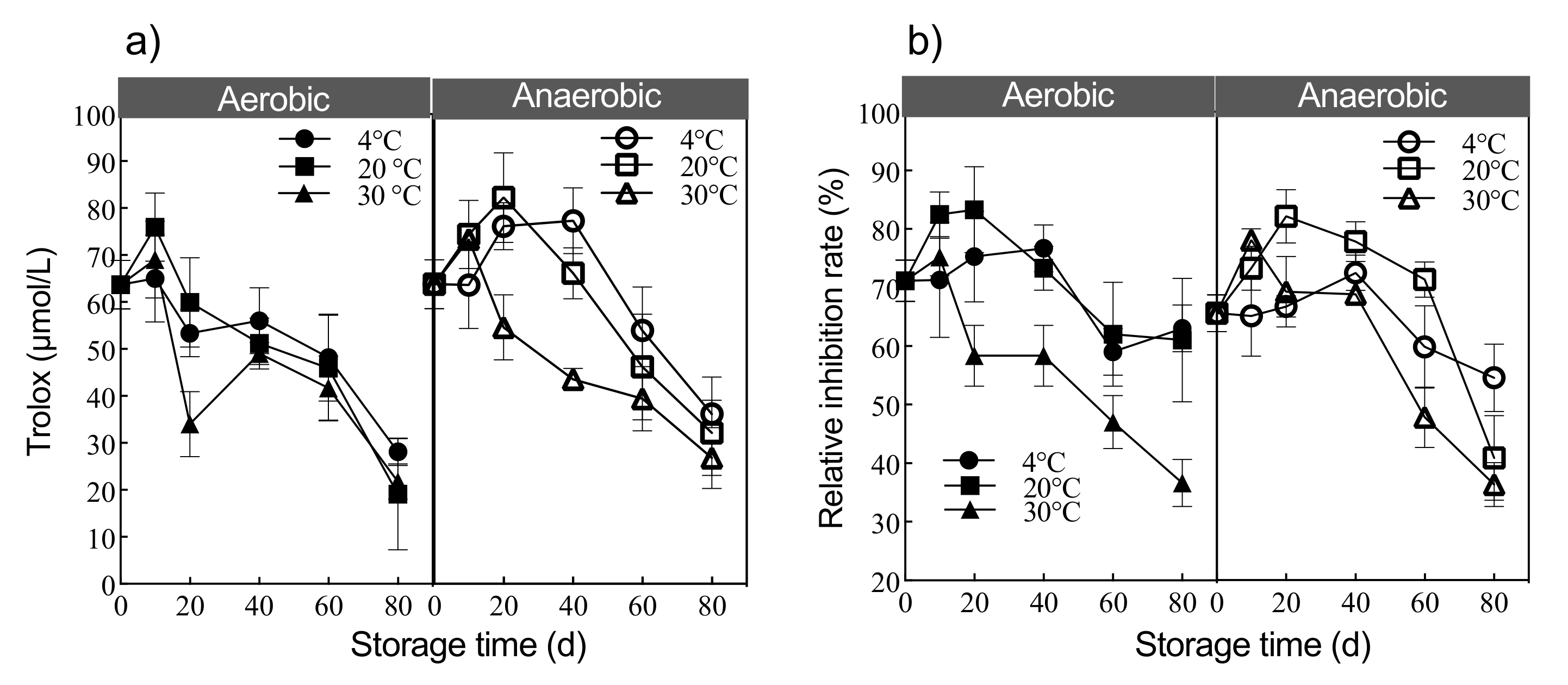
3.4. Antioxidant and Antifungal Properties of Digestate under Different Storage Modes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sobhi, M.; Gaballah, M.S.; Han, T.; Cui, X.; Li, B.; Sun, H.; Guo, J.; Dong, R. Nutrients recovery from fresh liquid manure through an airlift reactor to mitigate the greenhouse gas emissions of open anaerobic lagoons. J. Environ. Manag. 2021, 294, 112956. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Clark, M. Global diets link environmental sustainability and human health. Nature 2014, 515, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, A.J.; Assefa, Y.; Bond, H.D.; Haag, L.A.; Stone, L.R. Changes in soil nutrients after 10 years of cattle manure and swine effluent application. Soil Tillage Res. 2017, 172, 48–58. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, J.; Pan, M.; Hao, Y.; Hu, R.; Xiao, W.; Li, G.; Lyu, T. Valorisation of microalgae residues after lipid extraction: Pyrolysis characteristics for biofuel production. Biochem. Eng. J. 2022, 179, 108330. [Google Scholar] [CrossRef]
- Khoshnevisan, B.; Tsapekos, P.; Alvarado-Morales, M.; Rafiee, S.; Tabatabaei, M.; Angelidaki, I. Life cycle assessment of different strategies for energy and nutrient recovery from source sorted organic fraction of household waste. J. Clean. Prod. 2018, 180, 360–374. [Google Scholar] [CrossRef]
- Jadhav, P.; Muhammad, N.; Bhuyar, P.; Krishnan, S.; Abd Razak, A.S.; Zularisam, A.; Nasrullah, M. A review on the impact of conductive nanoparticles (CNPs) in anaerobic digestion: Applications and limitations. Environ. Technol. Innov. 2021, 23, 101526. [Google Scholar] [CrossRef]
- Dutta, S.; He, M.; Xiong, X.; Tsang, D.C. Sustainable management and recycling of food waste anaerobic digestate: A review. Bioresour. Technol. 2021, 341, 125915. [Google Scholar] [CrossRef]
- Li, G.; Bai, X.; Li, H.; Lu, Z.; Zhou, Y.; Wang, Y.; Cao, J.; Huang, Z. Nutrients removal and biomass production from anaerobic digested effluent by microalgae: A review. Int. J. Agric. Biol. Eng. 2019, 12, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Khoshnevisan, B.; Duan, N.; Tsapekos, P.; Awasthi, M.K.; Liu, Z.; Mohammadi, A.; Angelidaki, I.; Tsang, D.C.; Zhang, Z.; Pan, J. A critical review on livestock manure biorefinery technologies: Sustainability, challenges, and future perspectives. Renew. Sustain. Energy Rev. 2021, 135, 110033. [Google Scholar] [CrossRef]
- Tsapekos, P.; Khoshnevisan, B.; Alvarado-Morales, M.; Zhu, X.; Pan, J.; Tian, H.; Angelidaki, I. Upcycling the anaerobic digestion streams in a bioeconomy approach: A review. Renew. Sustain. Energy Rev. 2021, 151, 111635. [Google Scholar] [CrossRef]
- Paavola, T.; Rintala, J. Effects of storage on characteristics and hygienic quality of digestates from four co-digestion concepts of manure and biowaste. Bioresour. Technol. 2008, 99, 7041–7050. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, P.J.; Tompkins, D.; Pollard, S.J.; Hough, R.L.; Chambers, B.; Gale, P.; Tyrrel, S.; Villa, R.; Taylor, M.; Wu, S. Risk assessments for quality-assured, source-segregated composts and anaerobic digestates for a circular bioeconomy in the UK. Environ. Int. 2019, 127, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Styles, D.; Adams, P.; Thelin, G.; Vaneeckhaute, C.L.; Chadwick, D.; Withers, P.J. Life cycle assessment of biofertilizer production and use compared with conventional liquid digestate management. Environ. Sci. Technol. 2018, 52, 7468–7476. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, Y.; Akunna, J.C.; White, N.A.; Hallett, P.D.; Wheatley, R. Investigating the effects of anaerobic and aerobic post-treatment on quality and stability of organic fraction of municipal solid waste as soil amendment. Bioresour. Technol. 2008, 99, 8631–8636. [Google Scholar] [CrossRef]
- Vergote, T.L.; Bodé, S.; De Dobbelaere, A.E.; Buysse, J.; Meers, E.; Volcke, E.I. Monitoring methane and nitrous oxide emissions from digestate storage following manure mono-digestion. Biosyst. Eng. 2020, 196, 159–171. [Google Scholar] [CrossRef]
- Widyasari-Mehta, A.; Suwito, H.R.K.A.; Kreuzig, R. Laboratory testing on the removal of the veterinary antibiotic doxycycline during long-term liquid pig manure and digestate storage. Chemosphere 2016, 149, 154–160. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Li, G.; Luo, W.; Sun, Y. Manure digestate storage under different conditions: Chemical characteristics and contaminant residuals. Sci. Total Environ. 2018, 639, 19–25. [Google Scholar] [CrossRef]
- Tambone, F.; Adani, F.; Gigliotti, G.; Volpe, D.; Fabbri, C.; Provenzano, M.R. Organic matter characterization during the anaerobic digestion of different biomasses by means of CPMAS 13C NMR spectroscopy. Biomass Bioenergy 2013, 48, 111–120. [Google Scholar] [CrossRef]
- Li, G.; Bai, X.; Huo, S.; Huang, Z. Fast pyrolysis of LERDADEs for renewable biofuels. IET Renew. Power Gener. 2020, 14, 959–967. [Google Scholar] [CrossRef]
- Popp, D.; Schrader, S.; Kleinsteuber, S.; Harms, H.; Sträuber, H. Biogas production from coumarin-rich plants—inhibition by coumarin and recovery by adaptation of the bacterial community. FEMS Microbiol. Ecol. 2015, 91, 1–8. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Jannatizadeh, A.; Nojadeh, M.S.; Ebrahimzadeh, A. Exogenous melatonin ameliorates chilling injury in cut anthurium flowers during low temperature storage. Postharvest Biol. Technol. 2019, 148, 184–191. [Google Scholar] [CrossRef]
- Tao, Y.; Bie, X.-M.; Lv, F.-X.; Zhao, H.-Z.; Lu, Z.-X. Antifungal activity and mechanism of fengycin in the presence and absence of commercial surfactin against Rhizopus stolonifer. J. Microbiol. 2011, 49, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Muhmood, A.; Liu, H.; Dong, R.; Pang, S.; Wu, S. Exploring Bioactive Compounds in Anaerobically Digested Slurry: Extraction, Characterization, and Assessment of Antifungal Activity. Waste Biomass Valorization 2020, 11, 1863–1872. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [Green Version]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nuclc Acids Res. 2015, 43, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehsheikh, A.B.; Sourestani, M.M.; Zolfaghari, M.; Enayatizamir, N. Changes in soil microbial activity, essential oil quantity, and quality of Thai basil as response to biofertilizers and humic acid. J. Clean. Prod. 2020, 256, 120439. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, Y.; Guan, H.; Cui, T.; Liao, Y.; Wei, W.; Li, J.; Hassan, B.H.; Zhang, H.; Jia, X. Biochemical and structural characterization of the BioZ enzyme engaged in bacterial biotin synthesis pathway. Nat. Commun. 2021, 12, 2056. [Google Scholar] [CrossRef]
- Jog, R.; Nareshkumar, G.; Rajkumar, S. Enhancing soil health and plant growth promotion by actinomycetes. In Plant Growth Promoting Actinobacteria; Springer: Berlin/Heidelberg, Germany, 2016; pp. 33–45. [Google Scholar]
- Zhang, J.; Morris-Natschke, S.L.; Ma, D.; Shang, X.; Lee, K. Biologically active indolizidine alkaloids. Med. Res. Rev. 2020, 41, 928–960. [Google Scholar] [CrossRef]
- Malik, M.A.; Raza, M.K.; Dar, O.A.; Amadudin; Abid, M.; Wani, M.Y.; Al-Bogami, A.S.; Hashmi, A.A. Probing the antibacterial and anticancer potential of tryptamine based mixed ligand Schiff base Ruthenium(III) complexes. Bioorganic Chem. 2019, 87, 773–782. [Google Scholar] [CrossRef]
- Rajeswari, B.; Santhi, R. Antibacterial and antifungal activities of some novel imines of tryptamine. Mater. Today: Proc. 2019, 14, 323–331. [Google Scholar] [CrossRef]
- Surai, P.F.; Earle-Payne, K.; Kidd, M.T. Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models. Antioxidants 2021, 10, 1876. [Google Scholar] [CrossRef] [PubMed]
- Michalak, I. Marine Biologically Active Compounds as Feed Additives; MDPI: Basel, Switzerland, 2020. [Google Scholar]
- Ramli, A.N.M.; Sukri, N.A.M.; Azelee, N.I.W.; Bhuyar, P. Exploration of antibacterial and antioxidative activity of seed/peel extracts of Southeast Asian fruit Durian (Durio zibethinus) for effective shelf-life enhancement of preserved meat. J. Food Processing Preserv. 2021, 45, e15662. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | COD (g/L) | DOC (mg/L) | TN (g/L) | NH4+ (g/L) | TS (g/L) | VS (g/L) |
|---|---|---|---|---|---|---|
| 7.17 ± 0.06 | 4.43 ± 0.02 | 1.2 ± 0.04 | 3.09 ± 0.06 | 1.09 ± 0.08 | 45.4 ± 0.1 | 28.4 ± 0.1 |
| Metabolic Pathway | Total | Expected | Hits | Raw p | −log(p) | Holm Adjust | FDR | Impact |
|---|---|---|---|---|---|---|---|---|
| Steroid hormone biosynthesis | 99 | 4.73 | 17 | 2.25 × 10−6 | 1.3 × 101 | 1.8 × 10−4 | 1.8 × 10−4 | 0.4 |
| Purine metabolism | 20 | 0.96 | 5 | 1.99 × 10−3 | 6.22 | 1.57 × 10−1 | 7.95 × 10−2 | 0.26 |
| Cysteine and methionine metabolism | 56 | 2.68 | 8 | 4.41 × 10−3 | 5.42 | 3.44 × 10−1 | 1.18 × 10−1 | 0.19 |
| Biotin metabolism | 11 | 0.53 | 3 | 1.32 × 10−2 | 4.33 | 1 | 2.65 × 10−1 | 0.33 |
| Galactose metabolism | 41 | 1.96 | 5 | 4.32 × 10−2 | 3.14 | 1 | 6.92 × 10−1 | 0.13 |
| Pyruvate metabolism | 32 | 1.53 | 4 | 6.34 × 10−2 | 2.76 | 1 | 6.99 × 10−1 | 0.06 |
| Purine metabolism | 92 | 4.4 | 8 | 6.94 × 10−2 | 2.67 | 1 | 6.99 × 10−1 | 0.04 |
| Glyoxylate metabolism | 22 | 1.05 | 3 | 8.45 × 10−2 | 2.47 | 1 | 6.99 × 10−1 | 0.25 |
| Glycine, serine, and threonine metabolism | 50 | 2.39 | 5 | 8.74 × 10−2 | 2.44 | 1 | 6.99 × 10−1 | 0.5 |
| Starch and sucrose metabolism | 50 | 2.39 | 5 | 8.74 × 10−2 | 2.44 | 1 | 6.99 × 10−1 | 0.17 |
| Limonene and pinene degradation | 59 | 2.82 | 5 | 1.48 × 10−1 | 1.91 | 1 | 1 | 0.12 |
| Pyrimidine metabolism | 60 | 2.87 | 5 | 1.56 × 10−1 | 1.86 | 1 | 1 | 0.06 |
| Arachidonic acid metabolism | 62 | 2.96 | 5 | 1.72 × 10−1 | 1.76 | 1 | 1 | 0.1 |
| Methane metabolism | 34 | 1.62 | 3 | 2.2 × 10−1 | 1.52 | 1 | 1 | 0.39 |
| Caffeine metabolism | 21 | 1 | 2 | 2.65 × 10−1 | 1.33 | 1 | 1 | 0.11 |
| Nitrogen metabolism | 39 | 1.86 | 3 | 2.85 × 10−1 | 1.26 | 1 | 1 | 0.14 |
| Arginine and proline metabolism | 77 | 3.68 | 5 | 3.06 × 10−1 | 1.18 | 1 | 1 | 0.06 |
| Thiamine metabolism | 24 | 1.15 | 2 | 3.19 × 10−1 | 1.14 | 1 | 1 | 0.12 |
| Nicotinate and nicotinamide metabolism | 44 | 2.1 | 3 | 3.52 × 10−1 | 1.04 | 1 | 1 | 0 |
| One carbon pool by folate | 9 | 0.43 | 1 | 3.57 × 10−1 | 1.03 | 1 | 1 | 0 |
| Lysine degradation | 47 | 2.25 | 3 | 3.92 × 10−1 | 9.38 × 10−1 | 1 | 1 | 0.17 |
| Selenoamino acid metabolism | 48 | 2.29 | 3 | 4.05 × 10−1 | 9.05 × 10−1 | 1 | 1 | 0 |
| alpha-Linolenic acid metabolism | 29 | 1.39 | 2 | 4.07 × 10−1 | 8.99 × 10−1 | 1 | 1 | 0.38 |
| Terpenoid backbone biosynthesis | 33 | 1.58 | 2 | 4.73 × 10−1 | 7.48 × 10−1 | 1 | 1 | 0.16 |
| Propanoate metabolism | 35 | 1.67 | 2 | 5.05 × 10−1 | 6.84 × 10−1 | 1 | 1 | 0.01 |
| Tryptophan metabolism | 79 | 3.77 | 4 | 5.28 × 10−1 | 6.39 × 10−1 | 1 | 1 | 0.05 |
| Glutathione metabolism | 38 | 1.82 | 2 | 5.5 × 10−1 | 5.99 × 10−1 | 1 | 1 | 0.01 |
| Sulfur metabolism | 18 | 0.86 | 1 | 5.87 × 10−1 | 5.33 × 10−1 | 1 | 1 | 0.03 |
| Ascorbate and aldarate metabolism | 45 | 2.15 | 2 | 6.43 × 10−1 | 4.42 × 10−1 | 1 | 1 | 0.22 |
| Fructose and mannose metabolism | 48 | 2.29 | 2 | 6.78 × 10−1 | 3.89 × 10−1 | 1 | 1 | 0.02 |
| Alanine aspartate and glutamate metabolism | 24 | 1.15 | 1 | 6.93 × 10−1 | 3.67 × 10−1 | 1 | 1 | 0.02 |
| Tyrosine metabolism | 76 | 3.63 | 3 | 7.14 × 10−1 | 3.37 × 10−1 | 1 | 1 | 0.02 |
| Pantothenate and CoA biosynthesis | 27 | 1.29 | 1 | 7.35 × 10−1 | 3.07 × 10−1 | 1 | 1 | 0 |
| Glycerolipid metabolism | 32 | 1.53 | 1 | 7.93 × 10−1 | 2.31 × 10−1 | 1 | 1 | 0.01 |
| Lysine biosynthesis | 32 | 1.53 | 1 | 7.93 × 10−1 | 2.31 × 10−1 | 1 | 1 | 0.1 |
| Glycerophospholipid metabolism | 39 | 1.86 | 1 | 8.54 × 10−1 | 1.58 × 10−1 | 1 | 1 | 0.03 |
| Folate biosynthesis | 42 | 2.01 | 1 | 8.74 × 10−1 | 1.34 × 10−1 | 1 | 1 | 0.11 |
| Aminoacyl-tRNA biosynthesis | 75 | 3.58 | 2 | 8.83 × 10−1 | 1.25 × 10−1 | 1 | 1 | 0.06 |
| Metabolism of xenobiotics by cytochrome | 65 | 3.11 | 1 | 9.6 × 10−1 | 4.05 × 10−2 | 1 | 1 | 0 |
| Porphyrin and chlorophyll metabolism | 104 | 4.97 | 1 | 9.95 × 10−1 | 5.5 × 10−3 | 1 | 1 | 0.01 |
| Metabolic Pathway | Total | Expected | Hits | Raw p | −log(p) | Holm Adjust | FDR | Impact |
|---|---|---|---|---|---|---|---|---|
| Cyanoamino acid metabolism | 99 | 5.63 | 17 | 2.57 × 10−5 | 1.06 × 101 | 2.06 × 10−3 | 2.06 × 10−3 | 0.40 |
| Trypophane metabolism | 20 | 1.14 | 5 | 4.30 × 10−3 | 5.45 | 3.40× 10−1 | 1.72 × 10−1 | 0.26 |
| Typtophane biosynthesis | 32 | 1.82 | 6 | 8.08 × 10−3 | 4.82 | 6.31 × 10−1 | 2.16 × 10−1 | 0.06 |
| Cysteine and methionine metabolism | 56 | 3.19 | 8 | 1.25 × 10−2 | 4.38 | 9.62 × 10−1 | 2.50 × 10−1 | 0.19 |
| Tyrosine metabolism | 11 | 0.63 | 3 | 2.12 × 10−2 | 3.85 | 1.00 | 3.22 × 10−1 | 0.33 |
| Glutathione metabolism | 41 | 2.33 | 6 | 2.64 × 10−2 | 3.64 | 1.00 | 3.22 × 10−1 | 0.13 |
| Cyanoamino acid metabolism | 21 | 1.20 | 4 | 2.82 × 10−2 | 3.57 | 1.00 | 3.22 × 10−1 | 0.11 |
| Glycine, serine, and threonine metabolism | 48 | 2.73 | 6 | 5.20 × 10−2 | 2.96 | 1.00 | 5.20 × 10−1 | 0.19 |
| Methane metabolism | 39 | 2.22 | 5 | 6.73 × 10−2 | 2.70 | 1.00 | 5.97 × 10−1 | 0.14 |
| Purine metabolism | 92 | 5.24 | 9 | 7.46 × 10−2 | 2.60 | 1.00 | 5.97 × 10−1 | 0.04 |
| Pyrimidine metabolism | 60 | 3.42 | 6 | 1.23 × 10−1 | 2.10 | 1.00 | 7.08 × 10−1 | 0.07 |
| Galactose metabolism | 34 | 1.94 | 4 | 1.24 × 10−1 | 2.08 | 1.00 | 7.08 × 10−1 | 0.39 |
| Lysine degradation | 47 | 2.68 | 5 | 1.26 × 10−1 | 2.07 | 1.00 | 7.08 × 10−1 | 0.17 |
| Selenoamino acid metabolism | 22 | 1.25 | 3 | 1.26 × 10−1 | 2.07 | 1.00 | 7.08 × 10−1 | 0.25 |
| Starch and sucrose metabolism | 50 | 2.85 | 5 | 1.52 × 10−1 | 1.88 | 1.00 | 7.08 × 10−1 | 0.17 |
| Glyoxylate and dicarboxylate metabolism | 50 | 2.85 | 5 | 1.52 × 10−1 | 1.88 | 1.00 | 7.08 × 10−1 | 0.50 |
| Thiamine metabolism | 24 | 1.37 | 3 | 1.53 × 10−1 | 1.88 | 1.00 | 7.08 × 10−1 | 0.12 |
| Tryptophan metabolism | 79 | 4.50 | 7 | 1.59 × 10−1 | 1.84 | 1.00 | 7.08 × 10−1 | 0.23 |
| Nicotinate and nicotinamide metabolism | 44 | 2.50 | 4 | 2.39 × 10−1 | 1.43 | 1.00 | 9.71 × 10−1 | 0.00 |
| Limonene and pinene degradation | 59 | 3.36 | 5 | 2.43 × 10−1 | 1.42 | 1.00 | 9.71 × 10−1 | 0.12 |
| Arachidonic acid metabolism | 62 | 3.53 | 5 | 2.76 × 10−1 | 1.29 | 1.00 | 1.00 | 0.10 |
| Terpenoid backbone biosynthesis | 33 | 1.88 | 3 | 2.89 × 10-1 | 1.24 | 1.00 | 1.00 | 0.21 |
| Biotin metabolism | 38 | 2.16 | 3 | 3.69 × 10−1 | 9.98 × 10−1 | 1.00 | 1.00 | 0.01 |
| Glycerophospholipid metabolism | 39 | 2.22 | 3 | 3.85 × 10−1 | 9.55 × 10−1 | 1.00 | 1.00 | 0.10 |
| One carbon pool by folate | 9 | 0.51 | 1 | 4.10 × 10−1 | 8.91 × 10−1 | 1.00 | 1.00 | 0.00 |
| Arginine and proline metabolism | 77 | 4.38 | 5 | 4.49 × 10−1 | 8.01 × 10−1 | 1.00 | 1.00 | 0.06 |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | 27 | 1.54 | 2 | 4.61 × 10−1 | 7.75 × 10−1 | 1.00 | 1.00 | 0.00 |
| alpha-Linolenic acid metabolism | 29 | 1.65 | 2 | 4.98 × 10−1 | 6.96 × 10−1 | 1.00 | 1.00 | 0.38 |
| Fructose and mannose metabolism | 48 | 2.73 | 3 | 5.21 × 10−1 | 6.52 × 10−1 | 1.00 | 1.00 | 0.06 |
| Taurine and hypotaurine metabolism | 31 | 1.76 | 2 | 5.35 × 10−1 | 6.26 × 10−1 | 1.00 | 1.00 | 0.10 |
| Glycerolipid metabolism | 32 | 1.82 | 2 | 5.52 × 10−1 | 5.94 × 10−1 | 1.00 | 1.00 | 0.01 |
| Pentose and glucuronate interconversions | 53 | 3.02 | 3 | 5.90 × 10−1 | 5.28 × 10−1 | 1.00 | 1.00 | 0.04 |
| Propanoate metabolism | 35 | 1.99 | 2 | 6.02 × 10−1 | 5.08 × 10−1 | 1.00 | 1.00 | 0.01 |
| Steroid hormone biosynthesis | 16 | 0.91 | 1 | 6.10 × 10−1 | 4.95 × 10−1 | 1.00 | 1.00 | 0.00 |
| Aminoacyl-tRNA biosynthesis | 75 | 4.27 | 4 | 6.28 × 10−1 | 4.66 × 10−1 | 1.00 | 1.00 | 0.06 |
| Sulfur metabolism | 18 | 1.02 | 1 | 6.53 × 10−1 | 4.26 × 10−1 | 1.00 | 1.00 | 0.03 |
| Citrate cycle (TCA cycle) | 20 | 1.14 | 1 | 6.92 × 10−1 | 3.69 × 10−1 | 1.00 | 1.00 | 0.00 |
| Ascorbate and aldarate metabolism | 45 | 2.56 | 2 | 7.37 × 10−1 | 3.05 × 10−1 | 1.00 | 1.00 | 0.22 |
| Alanine, aspartate, and glutamate metabolism | 24 | 1.37 | 1 | 7.57 × 10−1 | 2.79 × 10−1 | 1.00 | 1.00 | 0.02 |
| Primary bile acid biosynthesis | 47 | 2.68 | 2 | 7.59 × 10−1 | 2.76 × 10−1 | 1.00 | 1.00 | 0.02 |
| Sphingolipid metabolism | 25 | 1.42 | 1 | 7.71 × 10−1 | 2.60 × 10−1 | 1.00 | 1.00 | 0.01 |
| Pantothenate and CoA biosynthesis | 27 | 1.54 | 1 | 7.96 × 10−1 | 2.28 × 10−1 | 1.00 | 1.00 | 0.00 |
| beta-Alanine metabolism | 28 | 1.59 | 1 | 8.08 × 10−1 | 2.13 × 10−1 | 1.00 | 1.00 | 0.10 |
| Tyrosine metabolism | 76 | 4.33 | 3 | 8.19 × 10−1 | 2.00 × 10−1 | 1.00 | 1.00 | 0.02 |
| Lysine biosynthesis | 32 | 1.82 | 1 | 8.49 × 10−1 | 1.64 × 10−1 | 1.00 | 1.00 | 0.10 |
| Inositol phosphate metabolism | 39 | 2.22 | 1 | 9.00 × 10−1 | 1.05 × 10−1 | 1.00 | 1.00 | 0.01 |
| Folate biosynthesis | 42 | 2.39 | 1 | 9.17 × 10−1 | 8.72 × 10−2 | 1.00 | 1.00 | 0.11 |
| Metabolism of xenobiotics by cytochrome | 65 | 3.70 | 1 | 9.79 × 10−1 | 2.12 × 10−2 | 1.00 | 1.00 | 0.00 |
| Porphyrin and chlorophyll metabolism | 104 | 5.92 | 2 | 9.85 × 10−1 | 1.50 × 10−2 | 1.00 | 1.00 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Muhmood, A.; Tsapekos, P.; Cui, X.; Guo, Y.; Zheng, Y.; Qiu, Y.; Wang, P.; Ren, L. Untargeted Metabolomics Profiling of Bioactive Compounds under Varying Digestate Storage Conditions: Assessment of Antioxidant and Antifungal Activity. Int. J. Environ. Res. Public Health 2022, 19, 4923. https://doi.org/10.3390/ijerph19084923
Lu J, Muhmood A, Tsapekos P, Cui X, Guo Y, Zheng Y, Qiu Y, Wang P, Ren L. Untargeted Metabolomics Profiling of Bioactive Compounds under Varying Digestate Storage Conditions: Assessment of Antioxidant and Antifungal Activity. International Journal of Environmental Research and Public Health. 2022; 19(8):4923. https://doi.org/10.3390/ijerph19084923
Chicago/Turabian StyleLu, Jiaxin, Atif Muhmood, Panagiotis Tsapekos, Xian Cui, Yuwen Guo, Yi Zheng, Yizhan Qiu, Pan Wang, and Lianhai Ren. 2022. "Untargeted Metabolomics Profiling of Bioactive Compounds under Varying Digestate Storage Conditions: Assessment of Antioxidant and Antifungal Activity" International Journal of Environmental Research and Public Health 19, no. 8: 4923. https://doi.org/10.3390/ijerph19084923