
Marathon Performance Depends on Pacing Oscillations between Non Symmetric Extreme Values



Abstract
:1. Introduction
2. Materials and Methods
3. Results
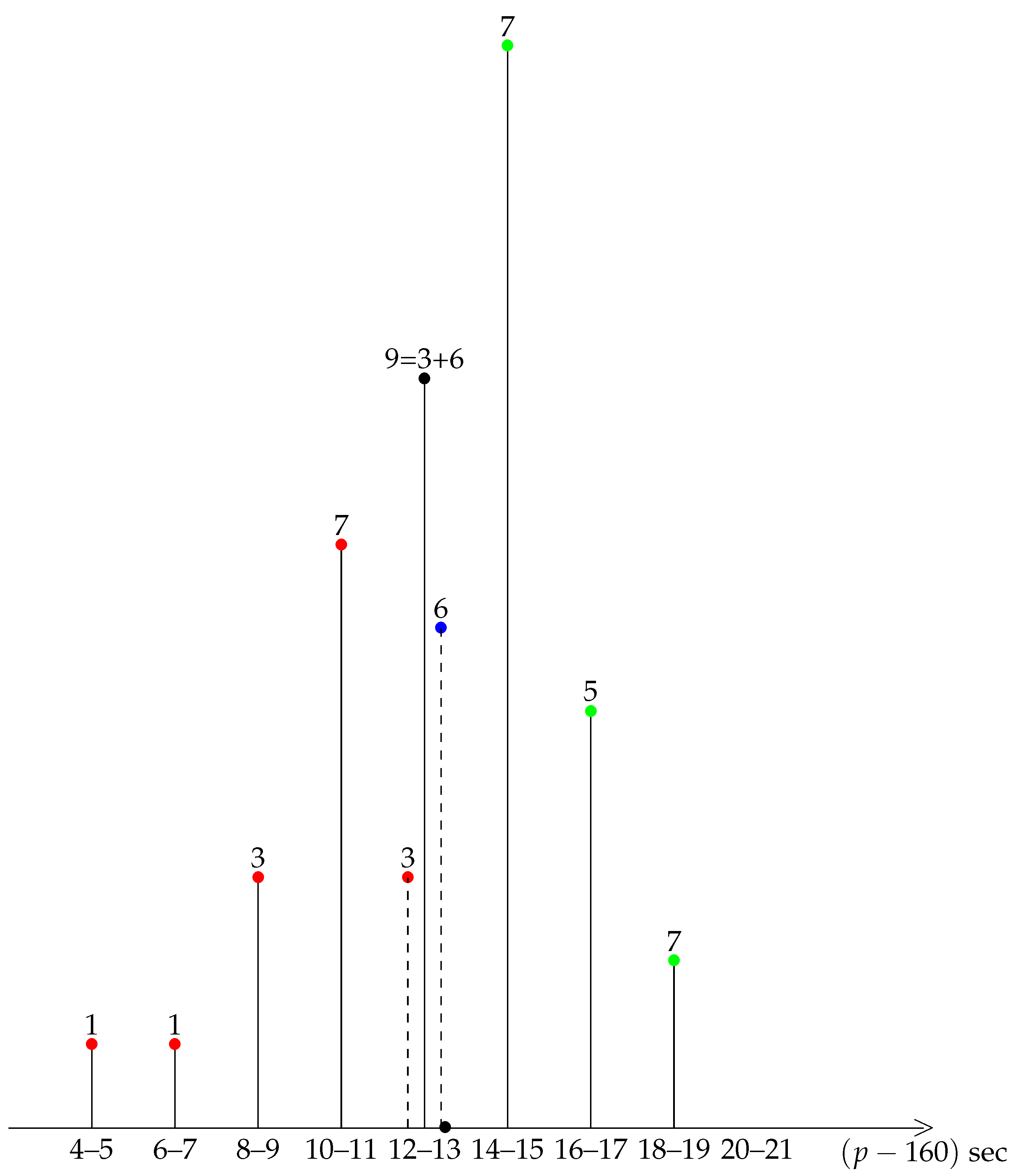
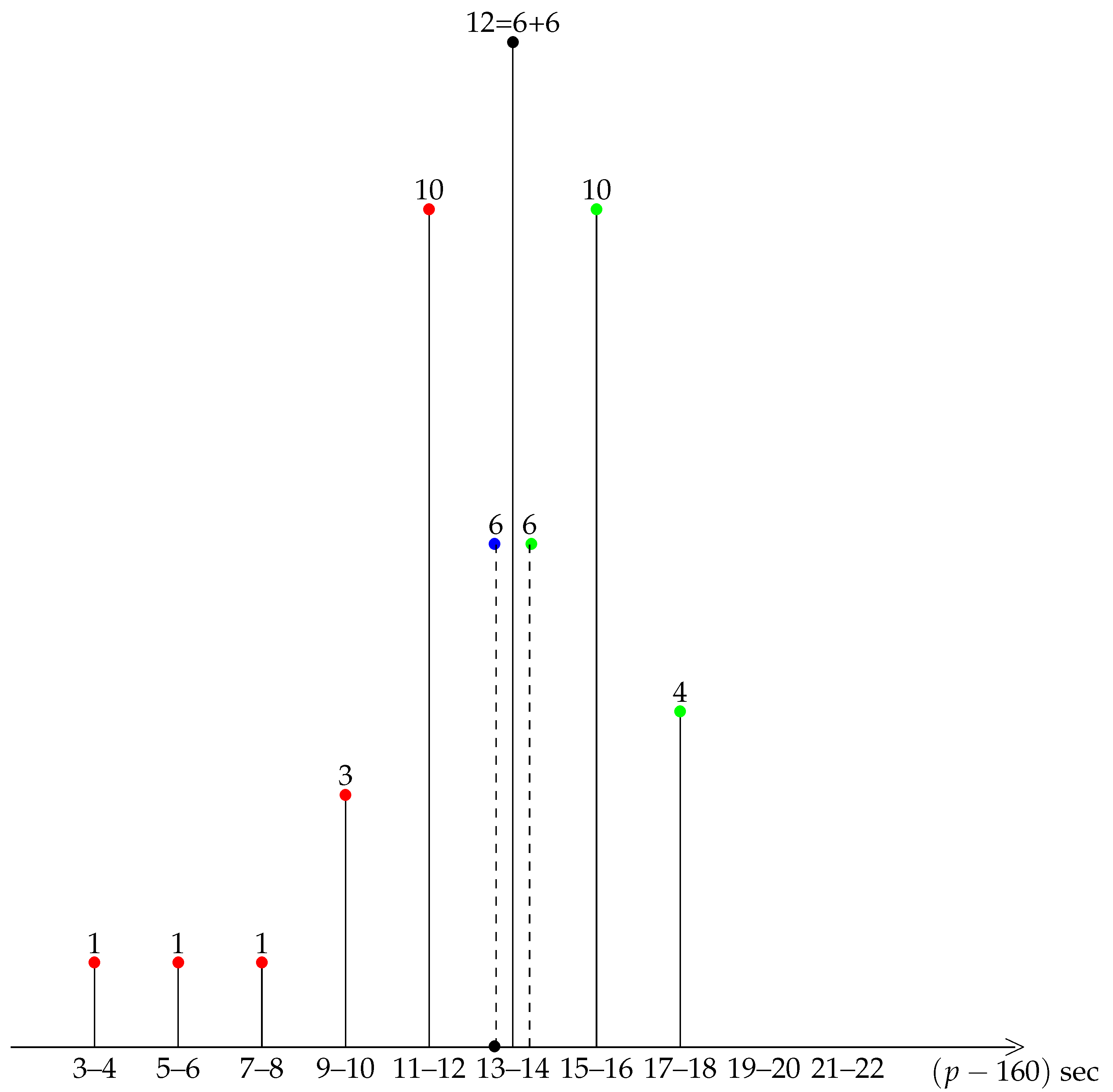
3.1. The Pace Is Not Uniform
3.2. The Pace Is Not Symmetric
3.3. The Sign of Asymmetry Is Not Due to Random Sampling Fluctuations
4. Discussion
4.1. The Pace Is Variable, Not Uniform
4.2. The Pace Is Not Symmetric, and the Sign of Asymmetry Is Explained via a “Mirror Race“, by Physiology, and Not by Sampling Fluctuations
4.3. Some Mathematical Speed Functions as the Bathtub with Negatively Skewed Pace Distributions
4.4. Oscillations between Extremes Values Allow the Interplay between Aerobic and Anaerobic Metabolisms, Optimizing the Recovery of Energy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, A. The Physiological Basis of Athletic Records. Nature 1925, 116, 544–548. [Google Scholar] [CrossRef]
- Emig, T.; Peltonen, J. Human running performance from real-world big data. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.B. A theory of competitive running. Phys. Today 1973, 26, 42–47. [Google Scholar] [CrossRef]
- Kennelly, A.E. An approximate law of fatigue in the speeds of racing animals. Proc. Am. Acad. Arts Sci. 1906, 42, 275–331. [Google Scholar] [CrossRef]
- Bernard, C. Introduction à la Médecine Expérimentale; Flammarion: Paris, France, 1984. [Google Scholar]
- Billat, V.; Carbillet, T.; Correa, M.; Pycke, J.R. Detecting the marathon asymmetry with a statistical signature. Phys. Stat. Mech. Appl. 2019, 515, 240–247. [Google Scholar] [CrossRef]
- Foster, C.; Hendrickson, K.J.; Peyer, K.; Reiner, B.; deKoning, J.J.; Lucia, A.; Battista, R.A.; Hettinga, F.J.; Porcari, J.P.; Wright, G. Pattern of developing the performance template. Br. J. Sport. Med. 2009, 43, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Lluch, J.; Rebollo, M.; Calduch-Losa, Á.; Mollá, R. Precision of wearable GPS in marathon races. IEEE Consum. Electron. Mag. 2020, 10, 32–38. [Google Scholar] [CrossRef]
- Stuart, A. Kendall’s Advanced Theory of Statistics; John Wiley & Sons: New York, NY, USA, 1994; Volume 1. [Google Scholar]
- David, H.A.; Nagaraja, H.N. Order Statistics; John Wiley & Sons: New York, NY, USA, 2004. [Google Scholar]
- Udny Yule, G.; Kendall, M. An Introduction to the Theory of Statistics. Charles Griffin & Company Limited: London, UK, 1950. [Google Scholar]
- Zar, J. Biostatistical Analysis, 4th ed.; Prentice-Hall: Englewood Cliffs, NJ, USA, 1999. [Google Scholar]
- Sheskin, D.J. Handbook of Parametric and Nonparametric Statistical Procedures; Chapman and Hall/CRC: London, UK, 2003. [Google Scholar]
- Johnson, N.L.; Kemp, A.W.; Kotz, S. Univariate Discrete Distributions; John Wiley & Sons: New York, NY, USA, 2005; Volume 444. [Google Scholar]
- Léger, L.A.; Ferguson, R.J. Effect of pacing on oxygen uptake and peak lactate for a mile run. Eur. J. Appl. Physiol. Occup. Physiol. 1974, 32, 251–257. [Google Scholar] [CrossRef]
- Ariyoshi, M.; Yamaji, K.; Shephard, R. Influence of running pace upon performance: Effects upon treadmill endurance time and oxygen cost. Eur. J. Appl. Physiol. Occup. Physiol. 1979, 41, 83–91. [Google Scholar] [CrossRef]
- Billat, V.L.; Slawinski, J.; Danel, M.; Koralsztein, J.P. Effect of free versus constant pace on performance and oxygen kinetics in running. Med. Sci. Sport. Exerc. 2001, 33, 2082–2088. [Google Scholar] [CrossRef] [Green Version]
- Cottin, F.; Papelier, Y.; Durbin, F.; Koralsztein, J.P.; Billat, V.L. Effect of fatigue on spontaneous velocity variations in human middle-distance running: Use of short-term Fourier transformation. Eur. J. Appl. Physiol. 2002, 87, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Sandals, L.E.; Wood, D.; Draper, S.B.; James, D. Influence of pacing strategy on oxygen uptake during treadmill middle-distance running. Int. J. Sport. Med. 2006, 27, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Garcin, M.; Danel, M.; Billat, V. Perceptual responses in free vs. constant pace exercise. Int. J. Sport. Med. 2008, 29, 453–459. [Google Scholar] [CrossRef]
- Ely, M.R.; Martin, D.E.; Cheuvront, S.N.; Montain, S.J. Effect of ambient temperature on marathon pacing is dependent on runner ability. Med. Sci. Sports Exerc. 2008, 40, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Vanhatalo, A.; Burnley, M.; Morton, R.H.; Poole, D.C. Critical power: Implications for determination of V O2max and exercise tolerance. Med. Sci. Sport. Exerc. 2010, 42, 1876–1890. [Google Scholar] [CrossRef]
- Fukuba, Y.; Whipp, B.J. A metabolic limit on the ability to make up for lost time in endurance events. J. Appl. Physiol. 1999, 87, 853–861. [Google Scholar] [CrossRef]
- Morton, R.H. A new modelling approach demonstrating the inability to make up for lost time in endurance running events. IMA J. Manag. Math. 2009, 20, 109–120. [Google Scholar] [CrossRef]
- Blondel, N.; Berthoin, S.; Billat, V.; Lensel, G. Relationship between run times to exhaustion at 90, 100, 120, and 140% of V O2max and velocity expressed relatively to critical velocity and maximal velocity. Int. J. Sport. Med. 2001, 22, 27–33. [Google Scholar] [CrossRef]
- Bassett, D.R., Jr. Scientific contributions of AV Hill: Exercise physiology pioneer. J. Appl. Physiol. 2002, 93, 1567–1582. [Google Scholar] [CrossRef]
- Billat, V.; Koralsztein, J.P.; Jacquot, S.; Brunel, N.J. Humans can freely sustain long-term constant acceleration in running via stochastic short-term corrections. Electron. J. Appl. Stat. Anal. 2018, 506, 290–304. [Google Scholar]
- Billat, V.L.; Demarle, A.; Sawinsky, J.; Paiva, M.; Koralszeit, J.P. Physical and training characteristics of top-class marathon runners. Med. Sci. Sport. Exerc. 2001, 33, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Kirby, B.S.; Clark, I.E.; Rice, H.M.; Fulkerson, E.; Wylie, L.J.; Wilkerson, D.P.; Vanhatalo, A.; Wilkins, B.W. Physiological demands of running at 2-hour marathon race pace. J. Appl. Physiol. 2021, 130, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.; Vitiello, D.; Palacin, F.; Correa, M.; Pycke, J.R. Race analysis of the world’s best female and male marathon runners. Int. J. Environ. Res. Public Health 2020, 17, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, F.Y.; Okuno, N.M.; Perandini, L.A.; Caldeira, L.F.S.; Simões, H.G.; Cardoso, J.R.; Bishop, D.J. Critical power can be estimated from nonexhaustive tests based on rating of perceived exertion responses. J. Strength Cond. Res. 2008, 22, 937–943. [Google Scholar] [CrossRef]
- Marino, F.E. The limitations of the constant load and self-paced exercise models of exercise physiology. Comp. Exerc. Physiol. 2010, 7, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Ulmer, H.V. Concept of an extracellular regulation of muscular metabolic rate during heavy exercise in humans by psychophysiological feedback. Experientia 1996, 52, 416–420. [Google Scholar] [CrossRef]
- Billman, G.E. Homeostasis: The underappreciated and far too often ignored central organizing principle of physiology. Front. Physiol. 2020, 11, 200. [Google Scholar] [CrossRef]
- Amann, M.; Eldridge, M.W.; Lovering, A.T.; Stickland, M.K.; Pegelow, D.F.; Dempsey, J.A. Arterial oxygenation influences central motor output and exercise performance via effects on peripheral locomotor muscle fatigue in humans. J. Physiol. 2006, 575, 937–952. [Google Scholar] [CrossRef]
- Casado, A.; Hanley, B.; Jiménez-Reyes, P.; Renfree, A. Pacing profiles and tactical behaviors of elite runners. J. Sport Health Sci. 2020, 10, 537–549. [Google Scholar] [CrossRef]
- Bramble, D.M.; Lieberman, D.E. Endurance running and the evolution of Homo. Nature 2004, 432, 345–352. [Google Scholar] [CrossRef]
- Cheval, B.; Boisgontier, M.P. The Theory of Effort Minimization in Physical Activity. Exerc. Sport Sci. Rev. 2021, 49, 168. [Google Scholar] [CrossRef] [PubMed]
- Abbiss, C.R.; Peiffer, J.J.; Meeusen, R.; Skorski, S. Role of ratings of perceived exertion during self-paced exercise: What are we actually measuring? Sport. Med. 2015, 45, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Andronov, A.; Vitt, A.; Khaikin, S. Theory of Oscillators, Translation; Pergamon Press: Oxford, UK, 1966. [Google Scholar]
- Farley, C.T.; Gonzalez, O. Leg stiffness and stride frequency in human running. J. Biomech. 1996, 29, 181–186. [Google Scholar] [CrossRef]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar] [CrossRef]
- Boullosa, D.; Medeiros, A.R.; Flatt, A.A.; Esco, M.R.; Nakamura, F.Y.; Foster, C. Relationships between Workload, Heart Rate Variability, and Performance in a Recreational Endurance Runner. J. Funct. Morphol. Kinesiol. 2021, 6, 30. [Google Scholar] [CrossRef]
- Foster, C.; Schrager, M.; Snyder, A.C.; Thompson, N.N. Pacing strategy and athletic performance. Sport. Med. 1994, 17, 77–85. [Google Scholar] [CrossRef]
- Myrkos, A.; Smilios, I.; Zafeiridis, A.; Iliopoulos, S.; Kokkinou, E.M.; Douda, H.; Tokmakidis, S.P. Effects of Work and Recovery Duration and Their Ratio on Cardiorespiratory and Metabolic Responses During Aerobic Interval Exercise. J. Strength Cond. Res. 2021. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Race | Total Time | V | |||
|---|---|---|---|---|---|
| Tokyo 2021 (E. K.) | 2 h 08 min 38 s | 182.4 s | 7.2 | 4% | − 0.31 |
| Berlin 2014 (D.K) | 2 h 2 min 57 s | 174.8 | 4.1 | 2% | − 0.47 |
| Berlin 2019 (K.B) | 2 h 01 min 41 s | 173.0 | 3.0 | 2% | − 0.55 |
| Berlin 2018 (E.K) | 2 h 01 min 39 s | 173.1 | 3.0 | 2% | − 0.82 |
| Vienna 2019 (E.K) | 1 h 59 min 40 s | 169.7 | 1.9 | 1% | − 2.32 |
| Monza 2017 (E.K) | 2 h 00 min 25 s | 171.2 | 1.8 | 1% | +0.56 |
| Race | ||||||||
|---|---|---|---|---|---|---|---|---|
| Tokyo 2021 | 167 | 194 | 27 | 189 | 254 | 15% | 10% | 7% |
| Berlin 2014 | 165 | 181 | 16 | 106 | 137 | 9% | 6% | 4% |
| Berlin 2019 | 165 | 179 | 14 | 76 | 98 | 8% | 4% | 3% |
| Berlin 2018 | 164 | 178 | 14 | 73 | 124 | 8% | 4% | 3% |
| Vienna 2019 | 161 | 172 | 11 | 43 | 43 | 6% | 2% | 1% |
| Monza 2017 | 168 | 176 | 8 | 43 | 58 | 5% | 3% | 2% |
| Race | z | p-Value of z | |
|---|---|---|---|
| Tokyo 2021 | −0.31 | −0.92 | 18% |
| Berlin 2014 | −0.47 | −1.36 | 9% |
| Berlin 2019 | −0.55 | −1.50 | 7% |
| Berlin 2018 | −0.82 | −2.25 | 1% |
| Vienna 2019 | −2.32 | −4.79 | |
| Monza 2017 | 0.56 | 2.07 | 2% |
| Race | ||||
|---|---|---|---|---|
| Tokyo 2021 | 27 | 15.4 | 11.6 | −3.8 |
| Berlin 2014 | 16 | 9.8 | 6.2 | −3.6 |
| Berlin 2018 | 14 | 9.1 | 4.9 | −4.2 |
| Vienna 2019 | 11 | 8.7 | 2.3 | −5.4 |
| Monza 2017 | 8 | 3.3 | 4.7 | 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pycke, J.-R.; Billat, V. Marathon Performance Depends on Pacing Oscillations between Non Symmetric Extreme Values. Int. J. Environ. Res. Public Health 2022, 19, 2463. https://doi.org/10.3390/ijerph19042463
Pycke J-R, Billat V. Marathon Performance Depends on Pacing Oscillations between Non Symmetric Extreme Values. International Journal of Environmental Research and Public Health. 2022; 19(4):2463. https://doi.org/10.3390/ijerph19042463
Chicago/Turabian StylePycke, Jean-Renaud, and Véronique Billat. 2022. "Marathon Performance Depends on Pacing Oscillations between Non Symmetric Extreme Values" International Journal of Environmental Research and Public Health 19, no. 4: 2463. https://doi.org/10.3390/ijerph19042463