
Enhancing Mechanisms of the Plant Growth-Promoting Bacterial Strain Brevibacillus sp. SR-9 on Cadmium Enrichment in Sweet Sorghum by Metagenomic and Transcriptomic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. PGPB and Cd-Contaminated Soil
2.2. Potted Plant Experimental Design and Treatments
2.3. Collection of Soil Samples
2.4. Analysis of the Soil Microbiota Composition
2.5. Metagenomic Analysis
2.6. Transcriptome Analysis of Sorghum Roots
2.7. Data Analysis
3. Results and Analysis
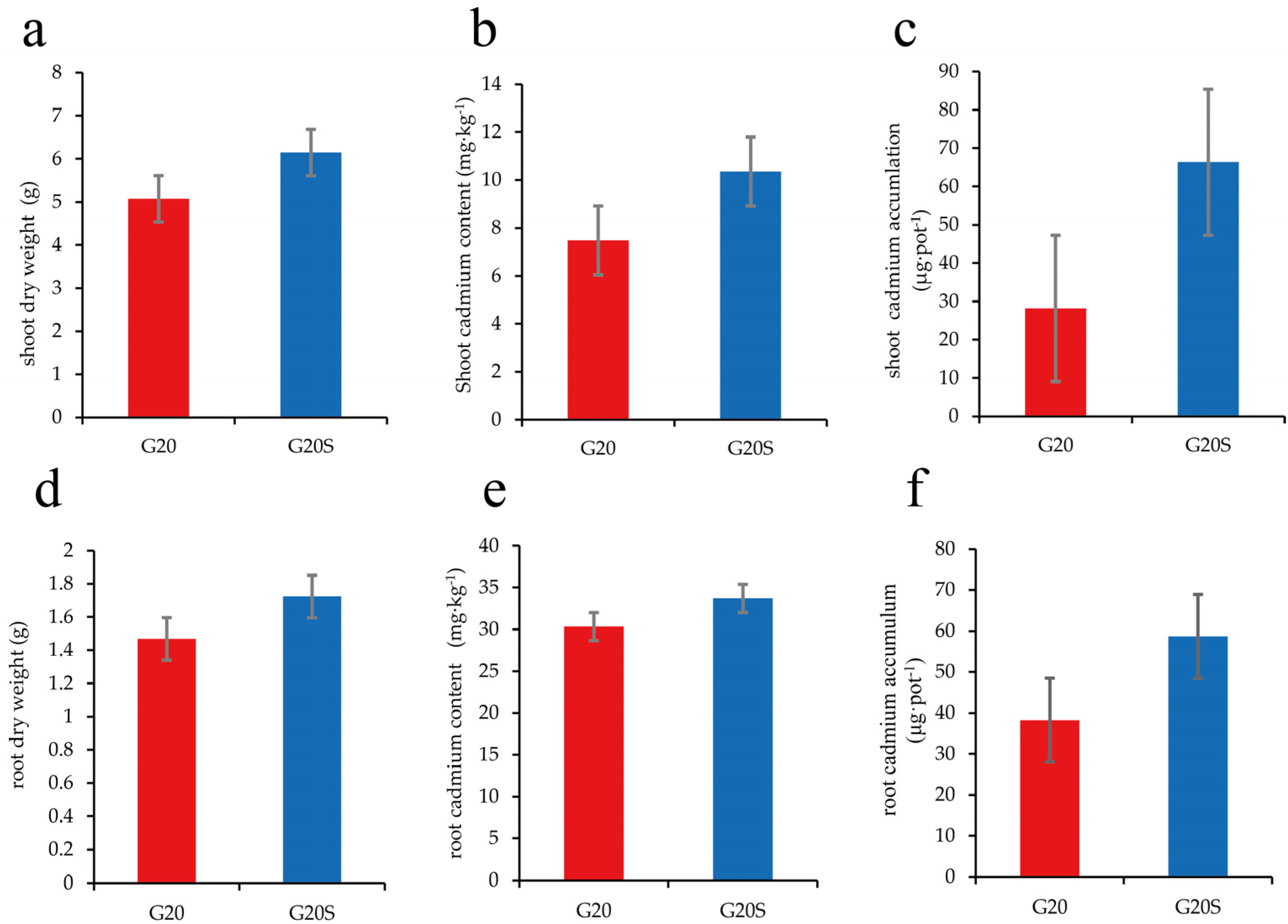
3.1. Effects of Brevibacillus SR-9 Inoculation on Sorghum Biomass and Cd Accumulation
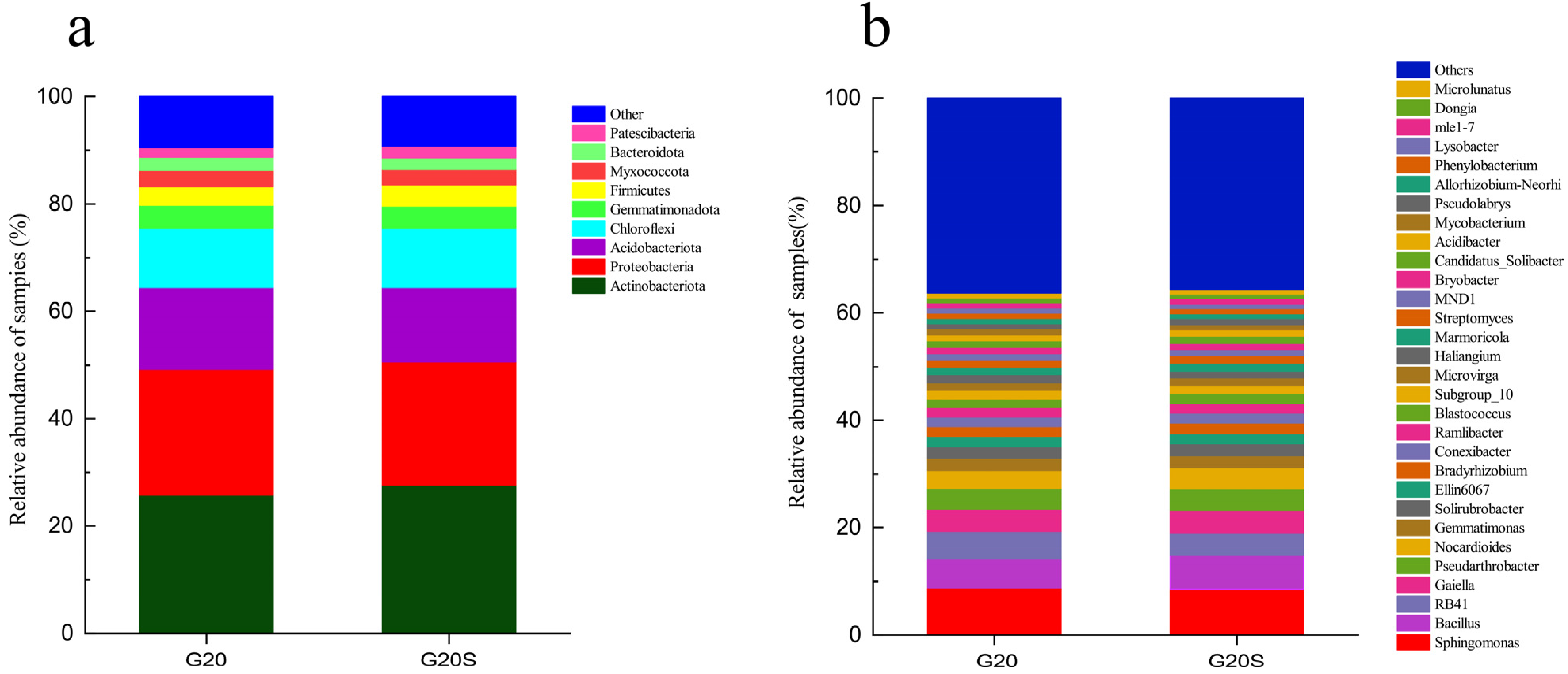
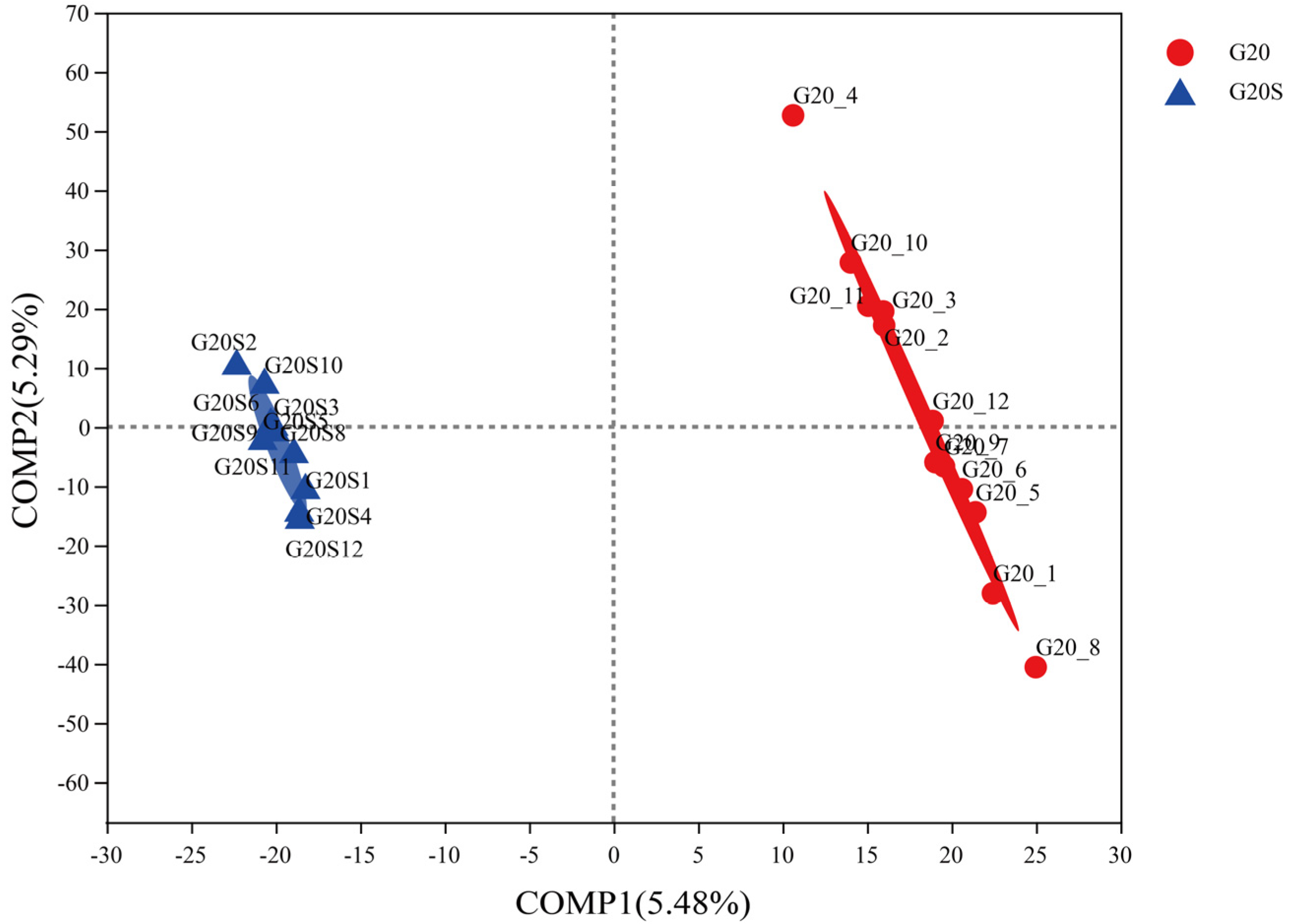
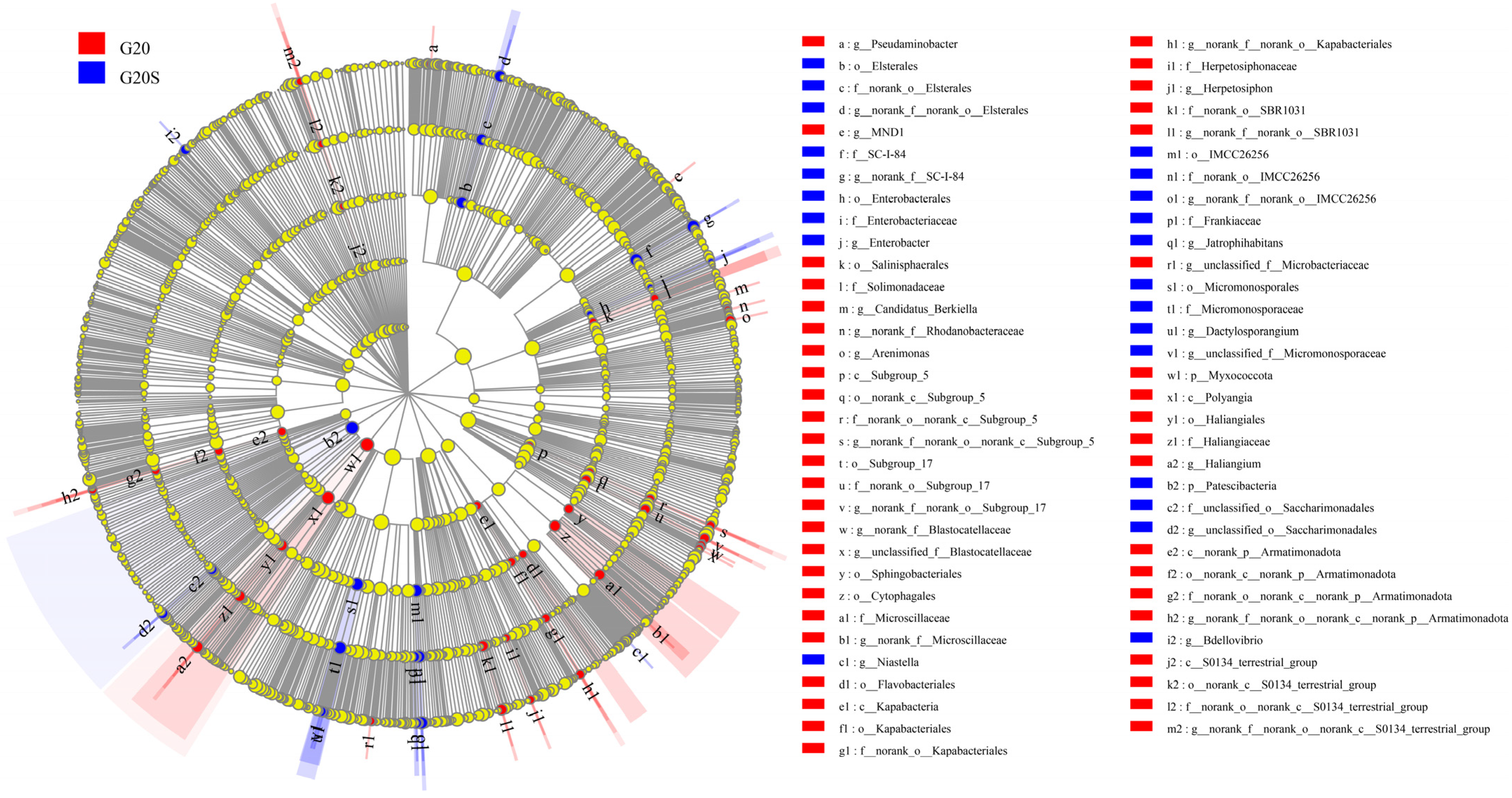
3.2. Microbial Community Composition
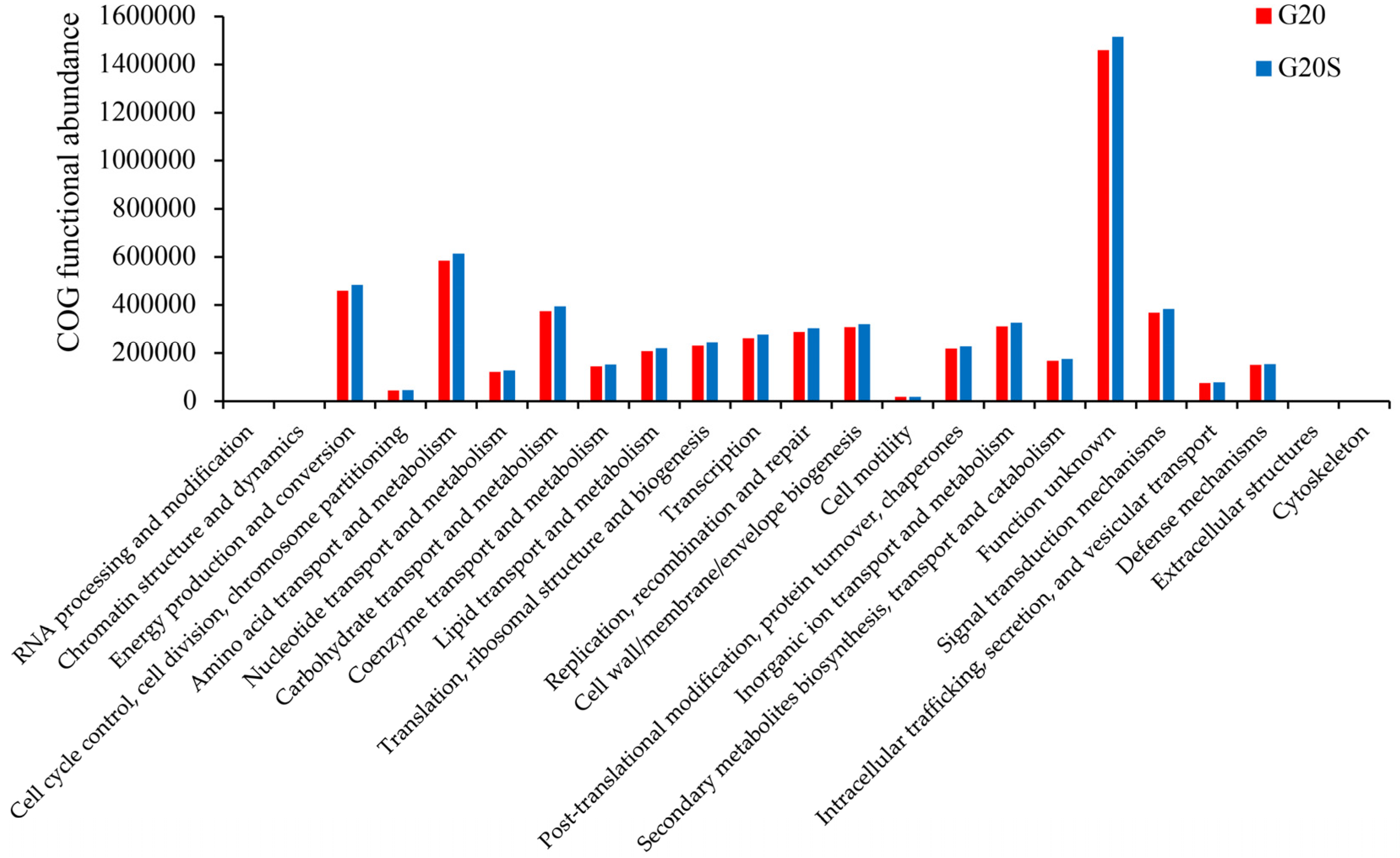
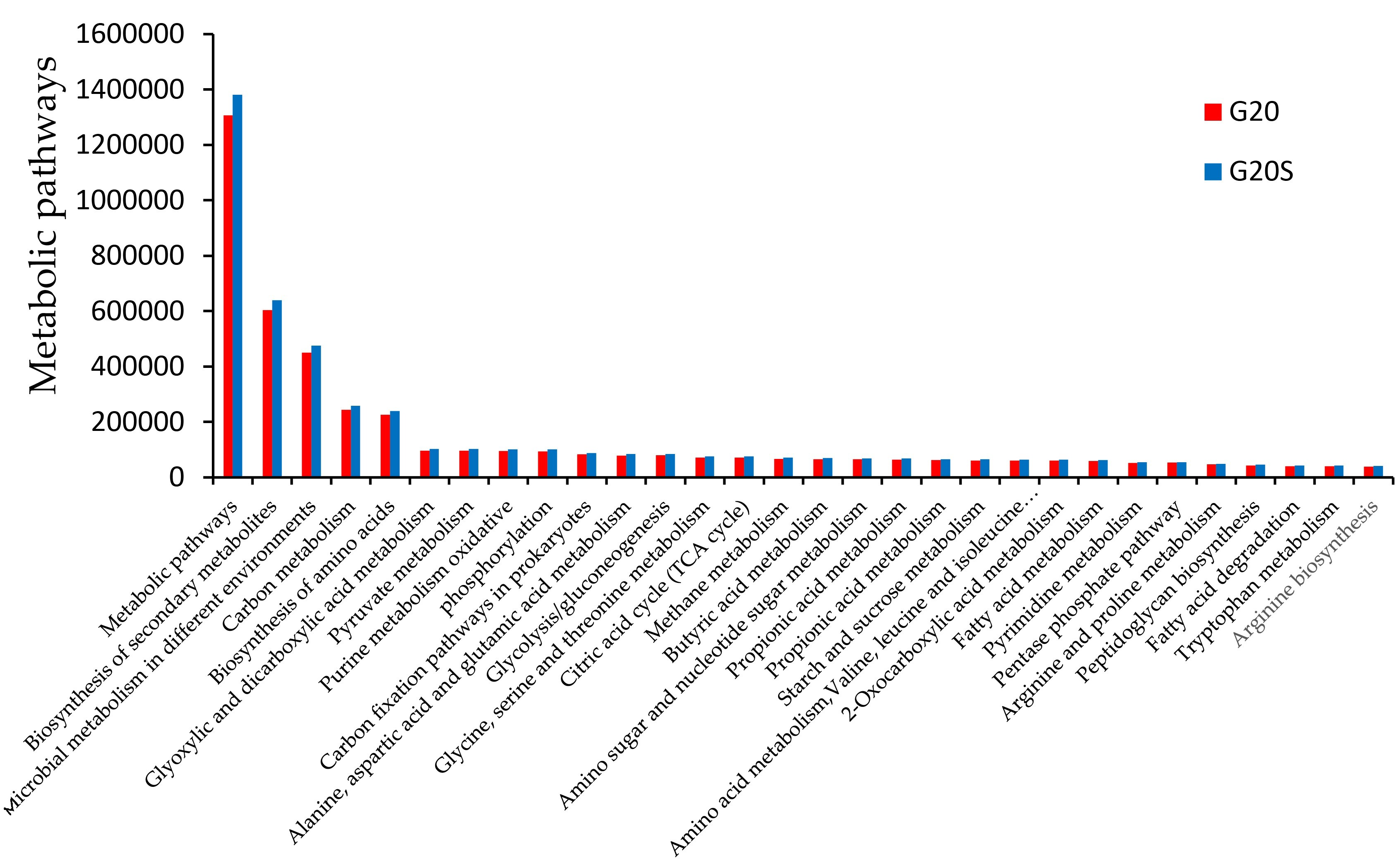
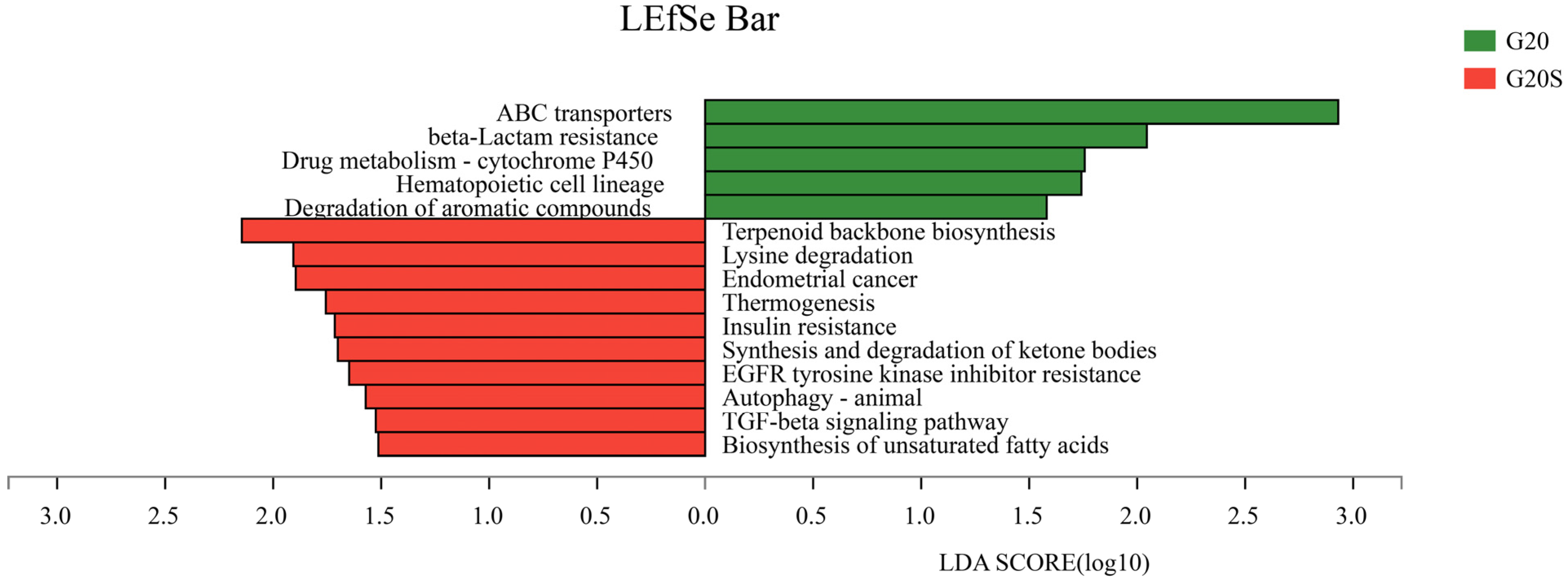
3.3. Metagenomic Analysis
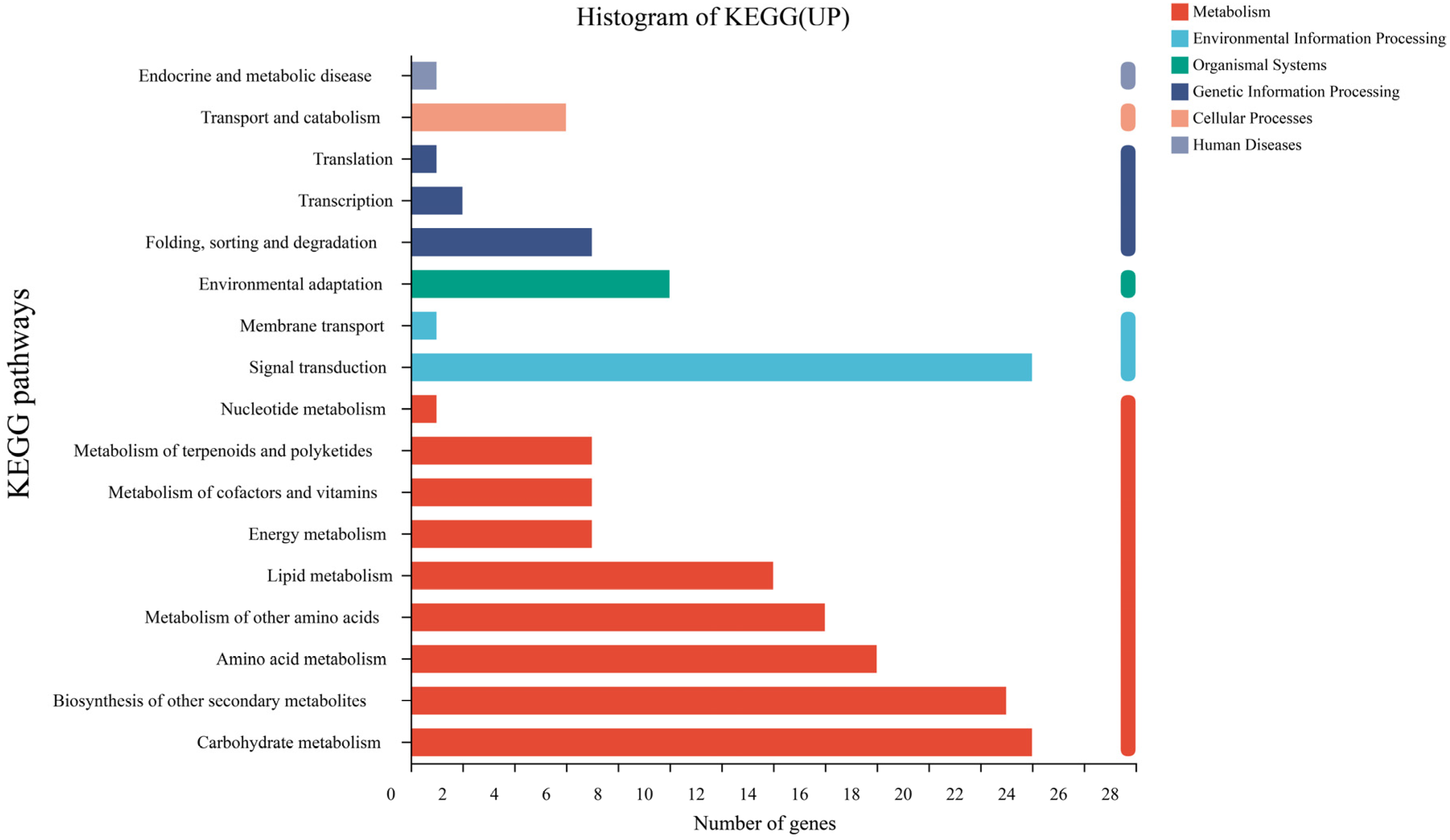
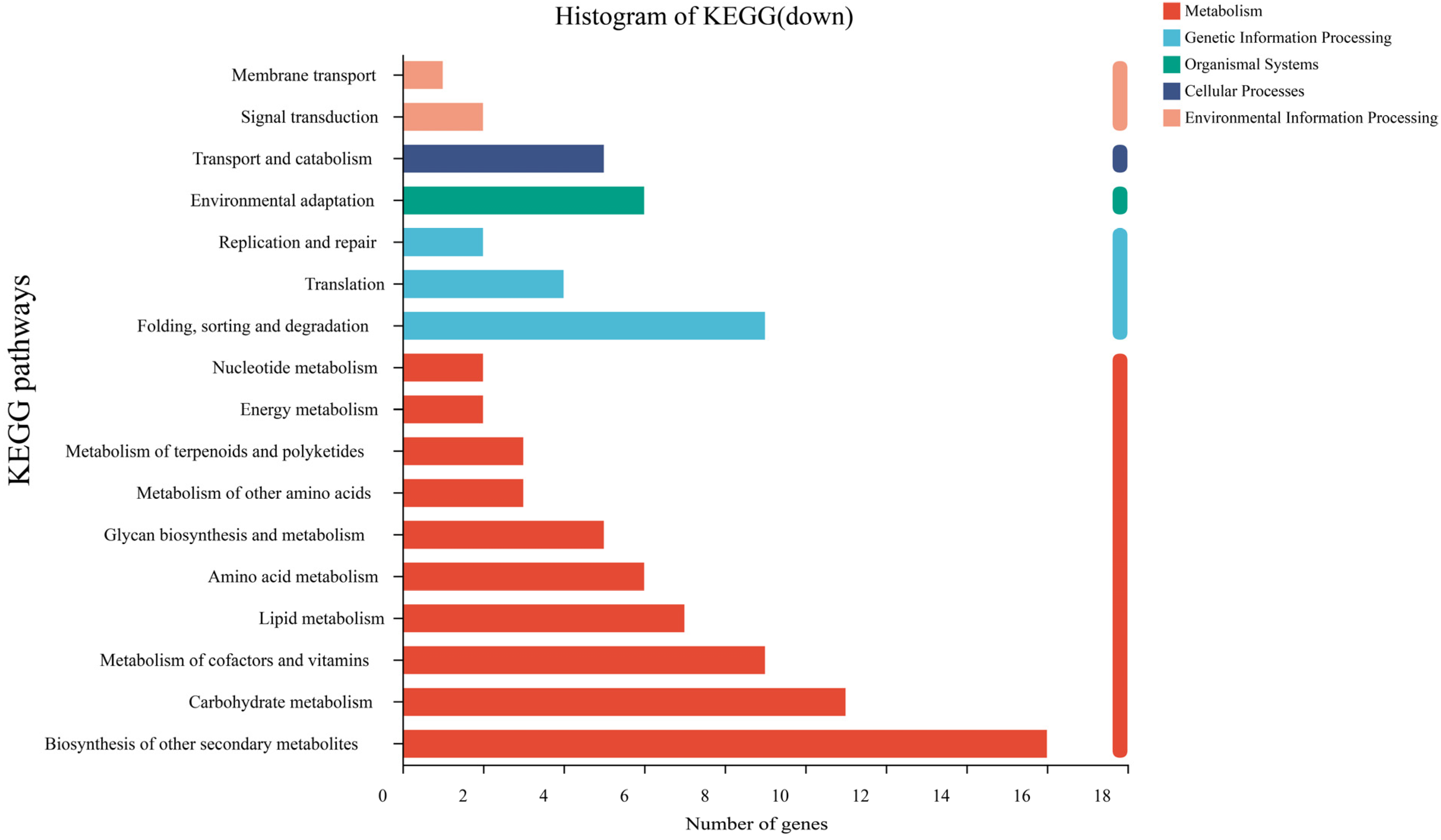
3.4. Transcriptome Gene Expression
4. Discussion
4.1. Strain SR-9 Inoculation Improves Sorghum Biomass and Cd Uptake
4.2. Strain SR-9 Inoculation Changes the Soil Microbial Community Composition
4.3. Strain SR-9 Inoculation Changes Soil Microbial Function According to Metagenomic Analysis
4.4. Strain SR-9 Inoculation Changes Gene Expression in Sorghum Roots According to Transcriptomic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, J.J.; Kim, Y.S.; Kumar, V. Heavy metal toxicity: An update of chelating therapeutic strategies. J. Trace Elem. Med. Biol. 2019, 54, 226–231. [Google Scholar] [CrossRef]
- Lao, Q.B.; Su, Q.Z.; Liu, G.Q.; Shen, Y.L.; Chen, F.J.; Lei, X.T.; Qing, S.M.; Wei, C.L.; Zhang, C.H.; Gao, J.S. Spatial distribution of and historical changes in heavy metals in the surface seawater and sediments of the Beibu Gulf, China. Mar. Pollut. Bull. 2019, 146, 427–434. [Google Scholar] [CrossRef]
- Vardhan, K.H.; Kumar, P.S.; Panda, R.C. A review on heavy metal pollution, toxicity and remedial measures: Current trends and future perspectives. J. Mol. Liq. 2019, 209, 111197. [Google Scholar] [CrossRef]
- Ministry of Environmental Protection; Ministry of Land and Resources. National soil pollution survey bulletin. China Environ. Prot. Ind. 2014, 5, 10–11.
- Belleghem, F.V.; Cuypers, A.; Semane, B.; Smeets, K.; Vangronsveld, J.; d’Haen, J.; Valcke, R. Subcellular localization of cadmium in roots and leaves of Arabidopsis thaliana. New Phytol. 2007, 173, 495–508. [Google Scholar] [CrossRef] [PubMed]
- He, J.L.; Luo, J.; Ma, C.F.; Li, S.J.; Qu, L.; Gai, Y.; Jiang, X.G.; Janz, D.; Polle, A. A Transcriptomic Network Underlies Microstructural and Physiological Responses to Cadmium in Populus × canescens. Plant Physiol. 2013, 162, 424–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Phytol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Mclaughlin, M.J.; Parker b, D.R.; Clarke, C. Metals and micronutrients—Food safety issues. Field Crop. Res. 1999, 60, 143–163. [Google Scholar] [CrossRef]
- Dias, M.C.; Monteiro, C.; Moutinho-Pereira, J.; Correia, C.; Gonçalves, B.; Santos, C. Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol. Plant 2013, 35, 1281–1289. [Google Scholar] [CrossRef]
- He, C.Q.; Zhao, Y.P.; Wang, F.F.; Oh, K.K.; Zhao, Z.Z.; Wu, C.L.; Zhang, X.Y.; Chen, X.Y.; Chen, X.L. Phytoremediation of soil heavy metals (Cd and Zn) by castor seedlings: Tolerance, accumulation and subcellular distribution. Chemosphere 2020, 252, 126471. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Huang, J.; Lu, J.; Sun, Y. Study on the influence of soil microbial community on the long-term heavy metal pollution of different land use types and depth layers in mine. Ecotoxicol. Environ. Saf. 2019, 170, 218–226. [Google Scholar] [CrossRef]
- Yan, A.; Wang, Y.M.; Tan, S.N.; Yusof, M.L.M.; Ghosh, S.; Chen, Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front Plant Sci. 2020, 11, 00359. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Sun, H.; Munis, M.F.H.; Fahad, S.; Yang, X.Y. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Shameer, S.; Prasad, V. Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regul. 2018, 84, 603–615. [Google Scholar] [CrossRef]
- Marques, A.P.; Moreira, H.; Franco, A.R.; Rangel, A.O.; Castro, P.M. Inoculating Helianthus annuus (sunflower) grown in zinc and cadmium contaminated soils with plant growth promoting bacteria—Effects on phytoremediation strategies. Chemosphere 2013, 92, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, F.Y.; Ge, J. Effect and mechanism of plant root exudates on soil remediation. Acta Ecol. Sin. 2022, 3, 1–14. [Google Scholar]
- Jia, W.T.; Lv, S.L.; Feng, J.J.; Li, J.H.; Li, Y.X.; Li, S.Z. Morphophysiological characteristic analysis demonstrated the potential of sweet sorghum (Sorghum bicolor (L.) Moench) in the phytoremediation of cadmium-contaminated soils. Environ. Sci. Pollut. Res. 2016, 23, 18823–18831. [Google Scholar] [CrossRef]
- Luo, S.L.; Xu, T.Y.; Chen, J.L.; Rao, C.; Xiao, X.; Wan, Y.; Zeng, G.M.; Liu, C.B.; Liu, Y.T. Endophyte-assisted promotion of biomass production and metal-uptake of energy crop sweet sorghum by plant-growth-promoting endophyte Bacillus sp. SLS18. Applied Microbiology and Biotechnology. Appl. Microbiol. Biot. 2011, 93, 1745–1753. [Google Scholar] [CrossRef]
- Gnansounou, E.; Dauriat, A.; Wyman, C.E. Refining sweet sorghum to ethanol and sugar: Economic trade-offs in the context of North China. Bioresour. Technol. 2005, 96, 985–1002. [Google Scholar] [CrossRef]
- Ratnavathi, C.V.; Chakravarthy, S.K.; Komala, V.V.; Chavan, U.D.; Patil, J.V. Sweet Sorghum as Feedstock for Biofuel Production: A Review. Sugar. Tech. 2011, 13, 399–407. [Google Scholar] [CrossRef]
- Zhuang, P.; Shu, W.S.; Li, Z.; Liao, B.; Li, J.T.; Shao, J.S. Removal of metals by sorghum plants from contaminated land. J. Environ. Manag. 2009, 21, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Metwali, M.R.; Gowayed, S.H.; Al-Maghrabi, O.; Mosleh, Y. Evaluation of toxic effect of copper and cadmium on growth, physiological traits and protein profile of wheat (Triticum aestivium L.), maize (Zea mays L.). World Appl. Sci. J. 2013, 21, 301–314. [Google Scholar]
- EI-Meihy, R.M.; Abou-Aly, H.M.; Youssef, A.M.; Tewfike, T.A.; EI-Alkshar, E.A. Efficiency of heavy metals-tolerant plant growth promoting bacteria for alleviating heavy metals toxicity on sorghum. Environ. Exp. Bot. 2019, 162, 295–301. [Google Scholar] [CrossRef]
- Bao, S.D. Siol Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Chen, Y.; Tian, W.; Shao, Y.; Li, Y.J.; Lin, L.A.; Zhang, Y.J.; Han, H.; Chen, Z.L. Miscanthus cultivation shapes rhizosphere microbial community structure and function as assessed by Illumina MiSeq sequencing combined with PICRUSt and FUNGUIld analyses. Arch. Microbiol. 2020, 202, 1157–1171. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, Y.M.; Huang, H.Y.; Mou, L.C.; Ru, J.L.; Xiao, S. Long-term and high-concentration heavy-metal contamination strongly influences the microbiome and functional genes in Yellow River sediments. Sci. Total Environ. 2018, 637–638, 1400–1412. [Google Scholar] [CrossRef]
- Abdollahi, S.; Golchin, A.; Shahryar, F.; Alamdari, P. PGPR inoculation of a contaminated soil affects plant growth and phytoavailability of Cd and Pb. Arch. Agron. Soil Sci. 2020, 68, 579–596. [Google Scholar] [CrossRef]
- Liu, A.; Wang, W.J.; Zheng, X.Y.; Chen, X.C.; Fu, W.T.; Wang, G.; Ji, J.; Guan, C.F. Improvement of the Cd and Zn phytoremediation efficiency of rice (Oryza sativa) through the inoculation of a metal-resistant PGPR strain. Chemosphere 2022, 302, 134900. [Google Scholar] [CrossRef]
- Ji, S.G.; Yan, Y.L.; Lyu, P. Study on heavy metal uptake in polluted soil by different sorghum germplasm. Chin. J. Eco-Agric. 2014, 5, 57–58. [Google Scholar]
- Yuan, X.Z.; Xiong, T.; Yao, C.; Yin, Y.N.; Li, H.C.; Li, N.S. A real filed phytoremediation of multi-metals contaminated soils by selected hybrid sweet sorghum with high biomass and high accumulation ability. Chemosphere 2019, 237, 124536. [Google Scholar] [CrossRef]
- Kong, Z.Y.; Glick, B.R. Chapter Two—The Role of Plant Growth-Promoting Bacteria in Metal Phytoremediation. Adv. Microb. Physiol. 2017, 71, 97–132. [Google Scholar]
- Zhang, L. Reinforcement of sweet sorghum by siderophore producing bacteria for remediation of heavy metal pollution in soil. Environ. Sci. Technol. 2014, 4, 74–79. [Google Scholar]
- Zhang, J.; Wang, L.H.; Yang, J.C.; Liu, H.; Dai, L.J. Health risk to residents and stimulation to inherent bacteria of various heavy metals in soil. Sci. Total Environ. 2015, 508, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.M.; Zhang, S.R.; Zhong, Q.M.; Gong, G.S.; Wang, G.Y.; Guo, X.; Xu, X.X. Effects of soil chemical properties and fractions of Pb, Cd, and Zn on bacterial and fungal communities. Sci. Total Environ. 2020, 715, 136904. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Bu, H.S.; Feng, Q.J.; Wassie, M.; Amee, M.; Jiang, Y.; Bi, Y.F.; Hu, L.X.; Chen, L. Identification of Cd-resistant microorganisms from heavy metal-contaminated soil and its potential in promoting the growth and Cd accumulation of bermudagrass. Environ. Res. 2021, 200, 111730. [Google Scholar] [CrossRef]
- Song, L.; Pan, Z.Z.; Dai, Y.; Chen, L.; Zhang, L.; Liao, Q.L.; Yu, X.Z.; Guo, H.Y.; Zhou, G.S. High-throughput sequencing clarifies the spatial structures of microbial communities in cadmium-polluted rice soils. Environ. Sci. Pollut. Res. 2021, 28, 47086–47098. [Google Scholar] [CrossRef]
- Chen, Q.L.; Ding, J.; Zhu, Y.G.; He, J.Z.; Hu, H.W. Soil bacterial taxonomic diversity is critical to maintaining the plant productivity. Environ. Int. 2020, 140, 105766. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Safe 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Rani, A.; Shouche, Y.S.; Goel, R. Declination of Copper Toxicity in Pigeon Pea and Soil System by Growth-Promoting Proteus vulgaris KNP3 Strain. Curr. Microbiol. 2008, 57, 78–82. [Google Scholar] [CrossRef]
- Li, Q.; Xing, Y.; Fu, X.W.; Ji, L.; Li, T.Y.; Wang, J.N.; Chen, Z.; Qi, Z.C.; Zhang, Q. Biochemical mechanisms of rhizospheric Bacillus subtilis-facilitated phytoextraction by alfalfa under cadmium stress—Microbial diversity and metabolomics analyses. Ecotox Environ. Safe 2021, 212, 112016. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Themaat, E.V.L.V.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant. Biol 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Hortal, S.; Bastida, F.; Armas, C.; Lozano, Y.M.; Moreno, J.L.; García, C.; Pugnaire, F.I. Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows. Soil Biol. Biochem. 2013, 64, 139–146. [Google Scholar] [CrossRef]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot 2013, 100, 1692–1705. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microb. Biot. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Raiz, M.; Shahzad, S.M.; Imran, Q.; Ali, I. Combined ability of chromium (Cr) tolerant plant growth promoting bacteria (PGPB) and salicylic acid (SA) in attenuation of chromium stress in maize plants. Plant. Physiol. Bioch. 2016, 108, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Srivastava, A.K.; Rajput, V.D.; Chauhan, P.K.; Bhojiya, A.A.; Jain, D.; Chaubey, G.; Dwivedi, P.; Sharma, B.; Minkina, T. Root Exudates: Mechanistic Insight of Plant Growth Promoting Rhizobacteria for Sustainable Crop Production. Front. Microbiol. 2022, 13, 916488. [Google Scholar] [CrossRef] [PubMed]
- Avalos, M.; Garbeva, P.; Vader, L.; Wezel, G.P.V.; Dickschat, J.S.; Ulanova, D. Biosynthesis, evolution and ecology of microbial terpenoids. Nat. Prod. Rep. 2022, 39, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.M.; Kang, B.R.; Kim, Y.C. Transcriptome Analysis of Induced Systemic Drought Tolerance Elicited by Pseudomonas chlororaphis O6 in Arabidopsis thaliana. Plant. Pathol. J. 2013, 29, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant. 2013, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Qiao, K.; Liang, S.; Wang, F.H.; Wang, H.; Hu, Z.L.; Chai, T.Y. Effects of cadmium toxicity on diploid wheat (Triticum urartu) and the molecular mechanism of the cadmium response. J. Hazard Mater. 2019, 374, 1–10. [Google Scholar] [CrossRef]
- Wang, J.; Duan, X.C.; Wang, Y.Z.; Sheng, J.J. Transcriptomic and physiological analyses of Miscanthus lutarioriparius in response to plumbum stress. Ind Crop. Prod. 2022, 176, 114305. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-Q.; Liu, Y.-Q.; Li, Y.-J.; Han, H.; Zhang, H.; Ji, M.-F.; Chen, Z.-J. Enhancing Mechanisms of the Plant Growth-Promoting Bacterial Strain Brevibacillus sp. SR-9 on Cadmium Enrichment in Sweet Sorghum by Metagenomic and Transcriptomic Analysis. Int. J. Environ. Res. Public Health 2022, 19, 16309. https://doi.org/10.3390/ijerph192316309
Li X-Q, Liu Y-Q, Li Y-J, Han H, Zhang H, Ji M-F, Chen Z-J. Enhancing Mechanisms of the Plant Growth-Promoting Bacterial Strain Brevibacillus sp. SR-9 on Cadmium Enrichment in Sweet Sorghum by Metagenomic and Transcriptomic Analysis. International Journal of Environmental Research and Public Health. 2022; 19(23):16309. https://doi.org/10.3390/ijerph192316309
Chicago/Turabian StyleLi, Xiao-Qi, Yong-Qi Liu, Ying-Jun Li, Hui Han, Hao Zhang, Ming-Fei Ji, and Zhao-Jin Chen. 2022. "Enhancing Mechanisms of the Plant Growth-Promoting Bacterial Strain Brevibacillus sp. SR-9 on Cadmium Enrichment in Sweet Sorghum by Metagenomic and Transcriptomic Analysis" International Journal of Environmental Research and Public Health 19, no. 23: 16309. https://doi.org/10.3390/ijerph192316309