
Ecological Responses of Maize Rhizosphere to Antibiotics Entering the Agricultural System in an Area with High Arsenicals Geological Background

{kind=link}
{kind=link}
{kind=link}
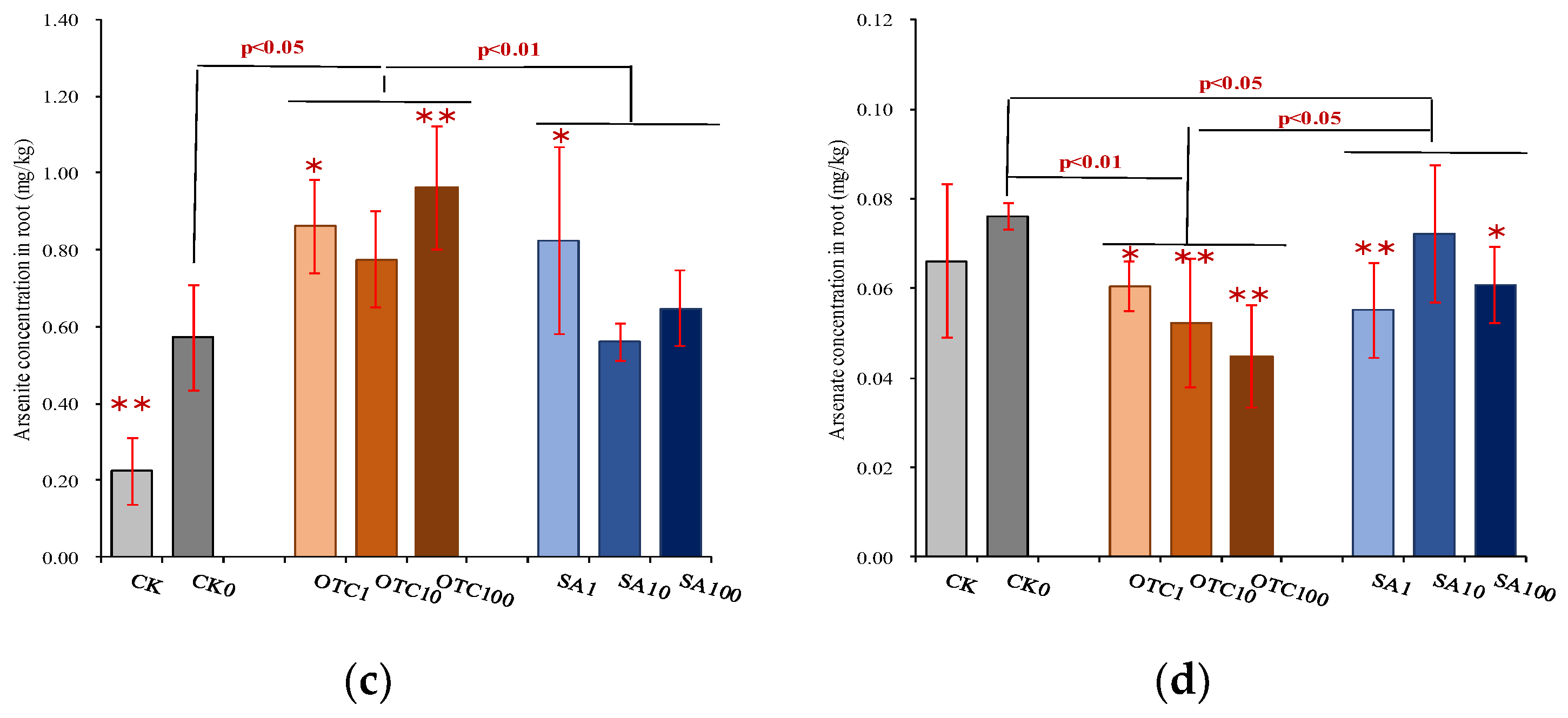{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soils and Organic Fertilizer Preparation
2.2. Pot Experiments Design
2.3. Soil Enzyme Activities
2.4. Soil Physiochemical Properties
2.5. Data Analysis
3. Results
3.1. Ecological Effects of Antibiotics on Rhizosphere Processes
3.2. Ecological Effects of Antibiotics on Soil Enzyme Activities
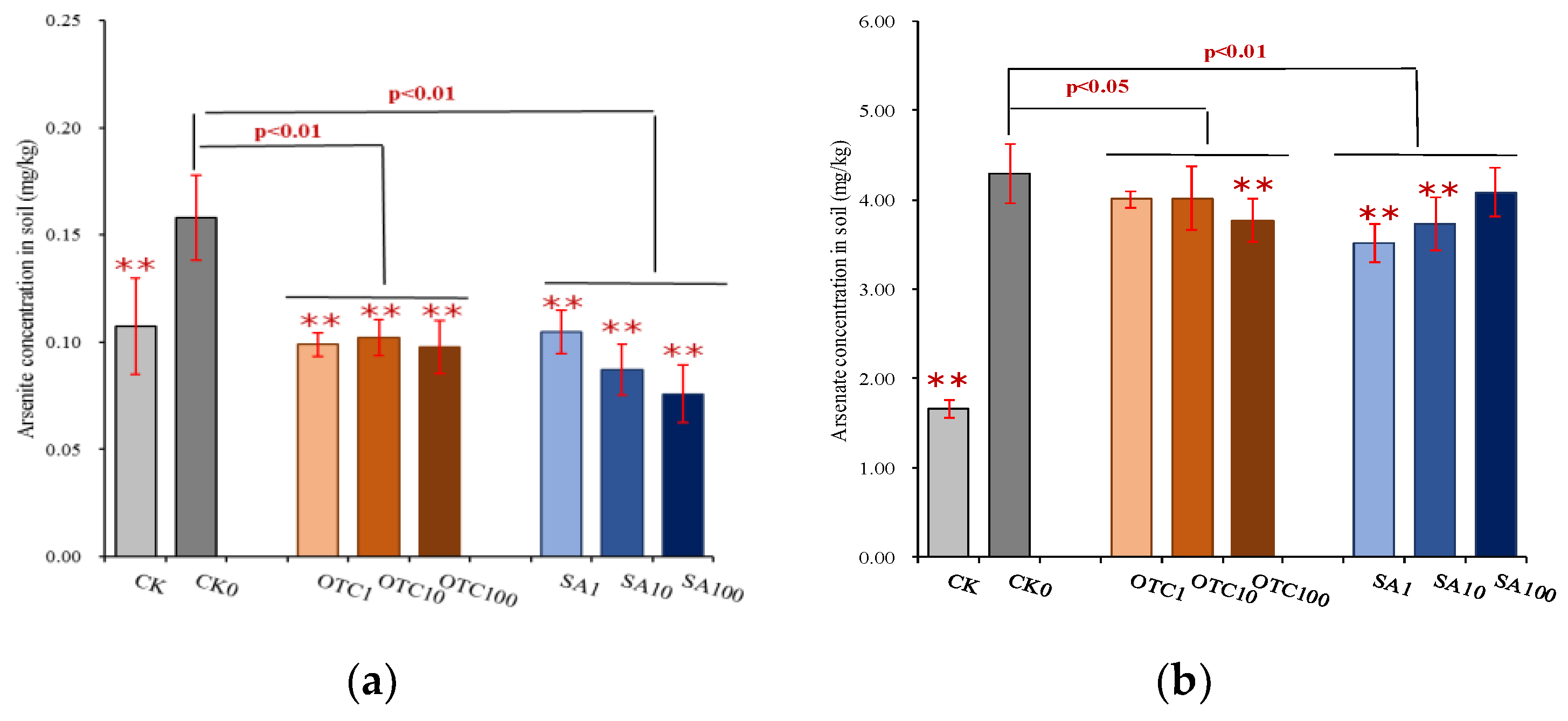
3.3. Ecological Effects of Antibiotics on Arsenic Mobilization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, X.; Wang, J.H.; Zhu, L.S.; Ge, W.L.; Wang, J. Environmental analysis of typical antibiotic-resistant bacteria and ARGs in farmland soil chronically fertilized with chicken manure. Sci. Total Environ. 2017, 593–594, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Alt, L.M.; Iverson, A.N.; Soupir, M.L.; Moorman, T.B.; Howe, A. Antibiotic resistance gene dissipation in soil microcosms amended with antibiotics and swine manure. J. Environ. Qual. 2021, 50, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Zhang, H.B.; Luo, Y.M.; Song, J. Occurrence and assessment of veterinary antibiotics in swine manures: A case study in East China. Chin. Sci. Bull. 2012, 57, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.K.; Chen, L.; Yang, L.D.; Li, S.J.; Sun, L. Bioaccumulation of antibiotics in crops under long-term manure application: Occurrence, biomass response and human exposure. Chemosphere 2019, 219, 882–895. [Google Scholar] [CrossRef]
- Tadić, Đ.; Bleda Hernandez, M.J.; Cerqueira, F.; Matamoros, V.; Piña, B.; Bayona, J.M. Occurrence and human health risk assessment of antibiotics and their metabolites in vegetables grown in field-scale agricultural systems. J. Hazard. Mater. 2021, 401, 123424. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganism 2019, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Seyoum, M.M.; Obayomi, O.; Bernstein, N.; Williams, C.F.; Gillor, O. Occurrence and distribution of antibiotics and corresponding antibiotic resistance genes in different soil types irrigated with treated wastewater. Sci. Total Environ. 2021, 782, 146835. [Google Scholar] [CrossRef]
- McRose, D.L.; Newman, D.K. Redox-active antibiotics enhance phosphorus bioavailability. Science 2021, 371, 1033–1037. [Google Scholar] [CrossRef]
- Yin, B.X.; Tan, S.; Wang, J.J.; Pan, K.; Wang, W.X.; Wang, X. Antibiotic application may raise the potential of methylmercury accumulation in fish. Sci. Total Environ. 2022, 819, 152946. [Google Scholar] [CrossRef]
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G. Antibiotic Resistance Gene Abundances Correlate with Metal and Geochemical Conditions in Archived Scottish Soils. PLoS ONE 2011, 6, e27300. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Asiani, K.; Arya, S.; Rensing, C.; Stekel, D.J.; Larsson, D.G.J.; Hobman, J.L. Metal Resistance and Its Association With Antibiotic Resistance. Adv. Microb. Physiol. 2017, 70, 261–313. [Google Scholar]
- Yazdankhah, S.; Rudi, K.; Bernhoft, A. Zinc and copper in animal feed—Development of resistance and co-resistance to antimicrobial agents in bacteria of animal origin. Microb. Ecol. Health Dis. 2014, 25, 25862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef]
- Zhao, Y.; Cocerva, T.; Cox, S.; Tardif, S.; Su, J.Q.; Zhu, Y.G.; Brandt, K.K. Evidence for co-selection of antibiotic resistance genes and mobile genetic elements in metal polluted urban soils. Sci. Total Environ. 2019, 656, 512–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, A.P.; Rajkumari, J.; Bhattacharjee, A.; Pandey, P. Development, spread and persistence of antibiotic resistance genes (ARGs) in the soil microbiomes through co-selection. Rev. Environ. Health 2020, 35, 371–378. [Google Scholar] [CrossRef]
- Song, J.; Rensing, C.; Holm, P.E.; Virta, M.; Brandt, K.K. Comparison of Metals and Tetracycline as Selective Agents for Development of Tetracycline Resistant Bacterial Communities in Agricultural Soil. Environ. Sci. Technol. 2017, 51, 3040–3047. [Google Scholar] [CrossRef]
- Wang, F.H.; Qiao, M.; Su, J.Q.; Chen, Z.; Zhou, X.; Zhu, Y.G. High Throughput Profiling of Antibiotic Resistance Genes in Urban Park Soils with Reclaimed Water Irrigation. Environ. Sci. Technol. 2014, 48, 9079–9085. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Gillings, M.; Simonet, P.; Stekel, D.; Banwart, S.; Penuelas, J. Human dissemination of genes and microorganismsin Earth’s Critical Zone. Glob. Chang. Biol. 2018, 24, 1488–1499. [Google Scholar] [CrossRef]
- Moriarty, M.M.; Koch, I.; Gordon, R.A.; Reimer, K.J. Arsenic speciation of terrestrial invertebrates. Environ. Sci. Technol. 2009, 43, 4818–4823. [Google Scholar] [CrossRef]
- Yang, H.C.; Rosen, B.P. New mechanisms of bacterial arsenic resistance. Biomed. J. 2016, 39, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Dunivin, T.K.; Yeh, S.Y.; Shade, A. A global survey of arsenic-related genes in soil microbiomes. BMC Biol. 2019, 17, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, M.J.; Jamieson, H.E.; Borčinová, R.A.; Maitland, K.; Oliver, J.; Falck, H.; Richardson, M. Mineralogical, geospatial, and statistical methods combined to estimate geochemical background of arsenic in soils for an area impacted by legacy mining pollution. Sci. Total Environ. 2021, 776, 145926. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.T.; Chi, Q.Q.; Zhu, D.; Li, G.; Ding, J.; An, X.L.; Zheng, F.; Zhu, Y.G.; Xue, X.M. Arsenic and Sulfamethoxazole Increase the Incidence of Antibiotic Resistance Genes in the Gut of Earthworm. Environ. Sci. Technol. 2019, 53, 10445–10453. [Google Scholar] [CrossRef]
- Cao, J.; Yang, G.; Mai, Q.; Zhuang, Z.; Zhuang, L. Co-selection of antibiotic-resistant bacteria in a paddy soil exposed to As (III) contamination with an emphasis on potential pathogens. Sci. Total Environ. 2020, 725, 138367. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Lu, C.; Liao, Q.; Gudda, F.O.; Ling, W. Antibiotics in animal manure and manure-based fertilizers: Occurrence and ecological risk assessment. Chemosphere 2020, 255, 127006. [Google Scholar] [CrossRef]
- Lima, T.; Domingues, S.; DaSilva, G.J. Manure as a Potential Hotspot for Antibiotic Resistance Dissemination by Horizontal Gene Transfer Events. Vet. Sci. 2020, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, Z.; Zhang, J. Improvement of indophenol blue colorimetric method on activity of urease in soil. J. Civ. Archit. Environ. Eng. 2012, 34, 102–107. [Google Scholar]
- Li, Z.G.; Luo, Y.M.; Teng, Y. Research Method of Soil and Environment Microorganisms; Science Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Chen, Y.S.; Xu, J.H.; Lv, Z.Y.; Huang, L.M.; Jiang, J.P. Impacts of biochar and oyster shells waste on the immobilization of arsenic in highly contaminated soils. J. Environ. Manag. 2018, 217, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Roose-Amsaleg, C.; Laverman, A.M. Do antibiotics have environmental side-effects? Impact of synthetic antibiotics on biogeochemical processes. Environ. Sci. Pollut. Res. 2016, 23, 4000–4012. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; Mcneill, A.M.; Combaret, C.P. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Caraccioloa, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Pashaei, R.; Zahedipour-Sheshglani, P.; Dzingelevičienė, R.; Abbasi, S.; Rees, R.M. Effects of pharmaceuticals on the nitrogen cycle in water and soil: A review. Environ. Monit. Assess. 2022, 194, 105. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.D.; Bernhardt, E. Are nitrogen and carbon cycle processes impacted by common stream antibiotics? A comparative assessment of single vs. mixture exposures. Natl. Cent. Biotechnol. Inf. 2022, 17, e0261714. [Google Scholar] [CrossRef]
- Singh, A.; Ghoshal, N. mpact of herbicide and various soil amendments on soil enzyme activities in a tropical rainfedagroecosystem. Eur. J. Soil Biol. 2013, 54, 56–62. [Google Scholar] [CrossRef]
- Veres, Z.; Kotroczó, Z.; Fekete, I.; Tóth, J.A.; Lajtha, K.; Townsend, K.; Tóthmérész, B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Appl. Soil Ecol. 2015, 92, 18–23. [Google Scholar] [CrossRef]
- McBride, S.G.; Strickland, M.S. Quorum sensing modulates microbial efficiency by regulating bacterial investment in nutrient acquisition enzymes. Soil Biol. Biochem. 2019, 136, 107514. [Google Scholar] [CrossRef]
- Feng, Y.; Hu, J.C.; Chen, Y.S.; Xu, J.H.; Jiang, J.P. Ecological response to antibiotics re-entering the aquaculture environment with possible long-term antibiotics selection based on enzyme activity in sediment. Environ. Sci. Pollut. Res. 2021, 29, 19033–19044. [Google Scholar] [CrossRef] [PubMed]
- Thiele-Bruhn, S.; Beck, I.C. Effects of sulfonamide and tetracycline antibiotics on soil microbial activity and microbial biomass. Chemosphere 2005, 59, 457–465. [Google Scholar] [CrossRef]
- Fang, H.; Han, Y.L.; Yin, Y.M.; Pan, X.; Yu, Y.L. Variations in dissipation rate, microbial function, and antibiotic resistance due to repeated introductions of manure containing sulfadiazine and chlortetracycline to soil. Chemosphere 2014, 96, 51–56. [Google Scholar] [CrossRef]
- Yao, Z.P.; Li, Z.J.; Lang, Y.C.; Zhang, Y.Q.; Yao, J.H.; Xie, X.Y. Dynamic responses of soil enzymatic activities to oxytetracycline in soil. Plant Nutr. Fertil. Sci. 2009, 15, 696–700. [Google Scholar] [CrossRef]
- Zhen, L.; Gu, J.; Hu, T.; Chen, Z.X. Effects of compost containing oxytetracycline on enzyme activities and microbial communities in maize rhizosphere soil. Environ. Sci. Pollut. Res. 2018, 25, 29459–29467. [Google Scholar] [CrossRef]
- Liu, F.; Ying, G.G.; Tao, R.; Zhao, J.L.; Yang, J.F.; Zhao, L.F. Effects of six selected antibiotics on plant growth and soil microbial and enzymatic activities. Environ. Pollut. 2009, 157, 1636–1642. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Zhang, H.; Shi, W.; Liu, Y. Bacterial Heavy-Metal and Antibiotic Resistance Genes in a Copper Tailing Dam Area in Northern China. Front. Microbiol. 2019, 10, 1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lan, B.; Fei, H.; Wang, S.; Zhu, G. Heavy metal could drive co-selection of antibiotic resistance in terrestrial subsurface soils. J. Hazard. Mater. 2021, 411, 124848. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Dinh, Q.T.; Chevreuil, M.; Garnier, J.; Roose-Amsaleg, C.; Labadie, P.; Laverman, A.M. The effect of environmental and therapeutic concentrations of antibiotics on nitrate reduction rates in river sediment. Water Res. 2013, 47, 3654–3662. [Google Scholar] [CrossRef] [PubMed]
- DeVries, S.L.; Zhang, P.F. Antibiotics and the Terrestrial Nitrogen Cycle: A Review. Curr. Pollut. Rep. 2016, 2, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.; Hou, L.; Liu, M.; Zheng, Y.; Li, X.; Lin, X.; Gao, J.; Jiang, X.; Wang, R.; Yu, C. Effects of multiple antibiotics exposure on denitrification process in the Yangtze Estuary sediments. Chemosphere 2017, 171, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, X.L.; Yu, Y.; Zhou, Y.; Chen, Q.; Sun, W. Impacts of engineered nanoparticles and antibiotics on denitrification: Element cycling functional genes and antibiotic resistance genes. Ecotoxicol. Environ. Saf. 2022, 241, 113787. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, R.; Rosen, B.P.; Phung, L.T.; Silver, S. Microbial arsenic: From geocycles to genes and enzymes. FEMS Microbiol. Rev. 2002, 26, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Massimo, P.; Antonio, G.C.; Lucia, C.; Alessia, S.; Violante, A. Arsenic in the Soil Environment: Mobility and Phytoavailability. Environ. Eng. Sci. 2015, 32, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Yamamura, S.; Watanabe, K.; Suda, W.; Tsuboi, S.; Watanabe, M. Effect of antibiotics onredox transformations of arsenic and diversity of arsenite-oxidizing bacteria in sediment microbial communities. Environ. Sci. Technol. 2014, 48, 350–357. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef] [Green Version]
- Kaviani, R.A.; Astaykina, A.; Streletskii, R.; Afsharyzad, Y.; Etesami, H.; Zarei, M.; Balasundram, S.K. An Overview of Antibiotic Resistance and Abiotic Stresses Affecting Antimicrobial Resistance in Agricultural Soils. Int. J. Environ. Res. Public Health 2022, 19, 4666. [Google Scholar] [CrossRef]
- Zhao, F.J.; Ma, J.F.; Meharg, A.A.; McGrath, S.P. Arsenic uptake and metabolism in plants. New Phytol. 2009, 181, 777–794. [Google Scholar] [CrossRef] [PubMed]
- Coelho, D.G.; Marinato, C.S.; de Matos, L.P.; de Andrade, H.M.; da Silva, V.M.; Santos-Neves, P.H.; Araújo, S.C.; Oliveira, J.A. Is arsenite more toxic than arsenate in plants? Ecotoxicology 2020, 29, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; You, G.; Zhang, M.; Peng, D.; Jiang, Z.; Qi, S.; Yang, S.; Hou, J. Antibiotic resistance genes alternation in soils modified with neutral and alkaline salts: Interplay of salinity stress and response strategies of microbes. Sci. Total Environ. 2022, 809, 152246. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Sui, Q.W.; Tong, J.; Zhang, H.; Wang, Y.W. Soil types influence the fate of antibiotic-resistant bacteria and antibiotic resistance genes following the land application of sludge composts. Environ. Int. 2018, 118, 34–43. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fiere, N.; Dantas, G. Bacterial phylogeny structures soil resistomes across habitats. Nature 2014, 509, 612–615. [Google Scholar] [CrossRef] [Green Version]
- Qiao, M.; Ying, G.G.; Singer, A.C. Review of antibiotic resistance in China and its environment. Environ. Int. 2018, 110, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Das, K.R.; Naik, M.M. Co-selection of multi-antibiotic resistance in bacterial pathogens in metal and microplastic contaminated environments: An emerging health threat. Chemosphere 2018, 215, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Pavón, A.; Riquelme, D.; Jaña, V.; Iribarren, C.; Manzano, C.; Lopez-Joven, C.; Reyes-Cerpa, S.; Navarrete, P.; Pavez, L.; García, K. The High Risk of Bivalve Farming in Coastal Areas With Heavy Metal Pollution and Antibiotic-Resistant Bacteria: A Chilean Perspective. Front. Cell. Infect. Microbiol. 2022, 12, 867446. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pawitwar, S.S.; Rosen, B.P. The organoarsenical biocycle and the primordial antibiotic methylarsenite. Metallomics 2016, 8, 1047–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, A.; Rana, R.; Saroha, T.; Patil, P.B. Discerning the role of a functional arsenic-resistance cassette in the evolution and adaptation of a rice pathogen. Microb. Genom. 2021, 7, 000608. [Google Scholar] [CrossRef]
- Wepking, C.; Badgley, B.; Barrett, J.E.; Knowlton, K.F.; Lucas, J.M.; Minick, K.J.; Ray, P.P.; Shawver, S.E.; Strickland, M.S. Prolonged exposure to manure from livestock-administered antibiotics decreases ecosystem carbon-use efficiency and alters nitrogen cycling. Ecol. Lett. 2019, 22, 2067–2076. [Google Scholar] [CrossRef]
- Zhang, P.; Mao, D.Q.; Gao, H.H.; Zheng, L.Y.; Chen, Z.Y.; Gao, Y.T.; Duan, Y.T.; Guo, J.H.; Luo, Y.; Ren, H.Q. Colonization of gut microbiota by plasmid-carrying bacteria is facilitated by evolutionary adaptation to antibiotic treatment. ISME J. 2021, 16, 1284–1293. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Chen, Y.; Feng, Y.; Li, X.; Ye, L.; Jiang, J. Ecological Responses of Maize Rhizosphere to Antibiotics Entering the Agricultural System in an Area with High Arsenicals Geological Background. Int. J. Environ. Res. Public Health 2022, 19, 13559. https://doi.org/10.3390/ijerph192013559
Li M, Chen Y, Feng Y, Li X, Ye L, Jiang J. Ecological Responses of Maize Rhizosphere to Antibiotics Entering the Agricultural System in an Area with High Arsenicals Geological Background. International Journal of Environmental Research and Public Health. 2022; 19(20):13559. https://doi.org/10.3390/ijerph192013559
Chicago/Turabian StyleLi, Mengli, Yongshan Chen, Ying Feng, Xiaofeng Li, Lili Ye, and Jinping Jiang. 2022. "Ecological Responses of Maize Rhizosphere to Antibiotics Entering the Agricultural System in an Area with High Arsenicals Geological Background" International Journal of Environmental Research and Public Health 19, no. 20: 13559. https://doi.org/10.3390/ijerph192013559