
Coal Mining Activities Driving the Changes in Microbial Community and Hydrochemical Characteristics of Underground Mine Water
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
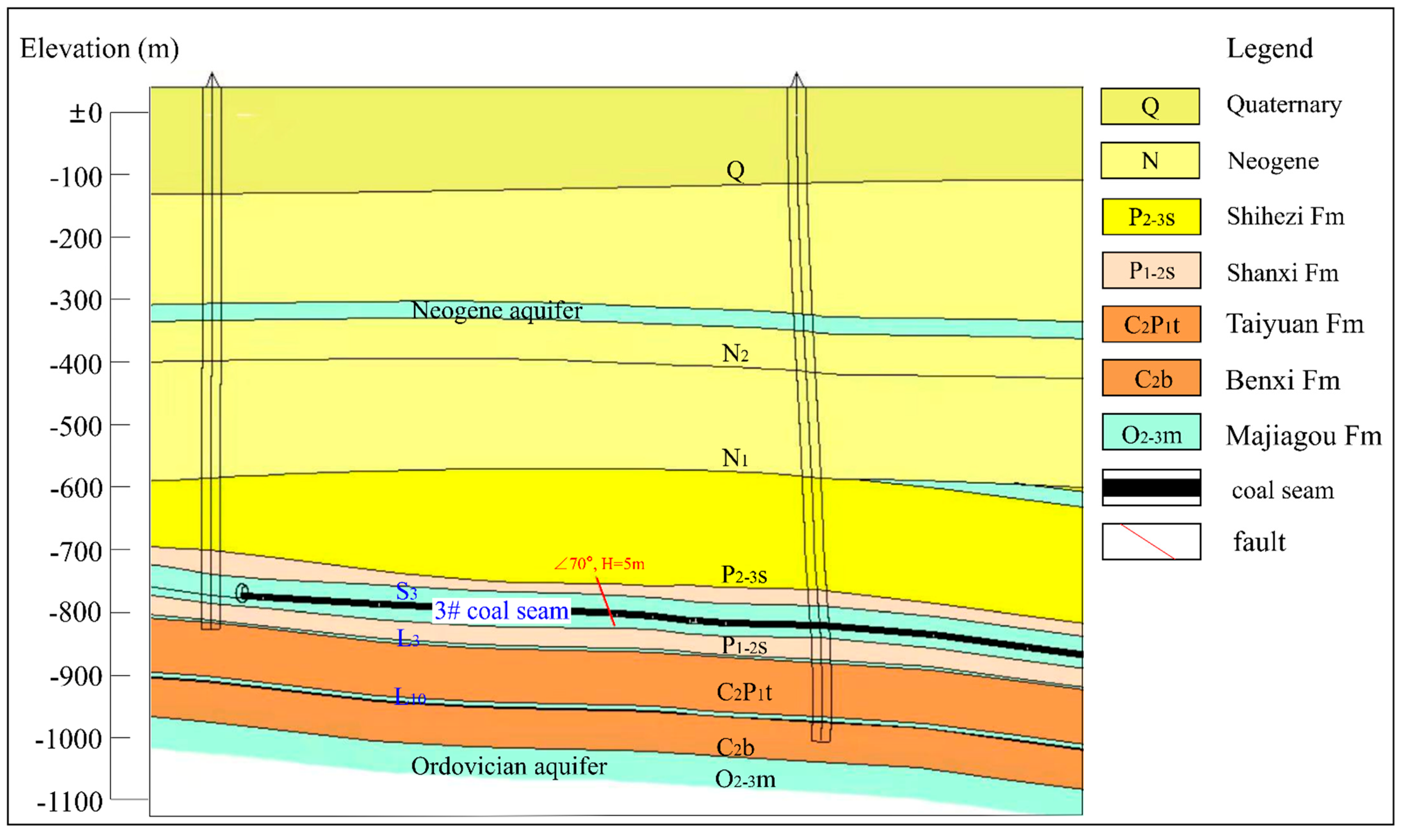
2.1. Study Sites and Sample Collection
2.2. Hydrochemical Characterization
2.3. DNA Extraction and 16S rRNA Gene Sequencing
2.4. Processing and Statistical Analyses of Sequence Data
3. Results and Discussion
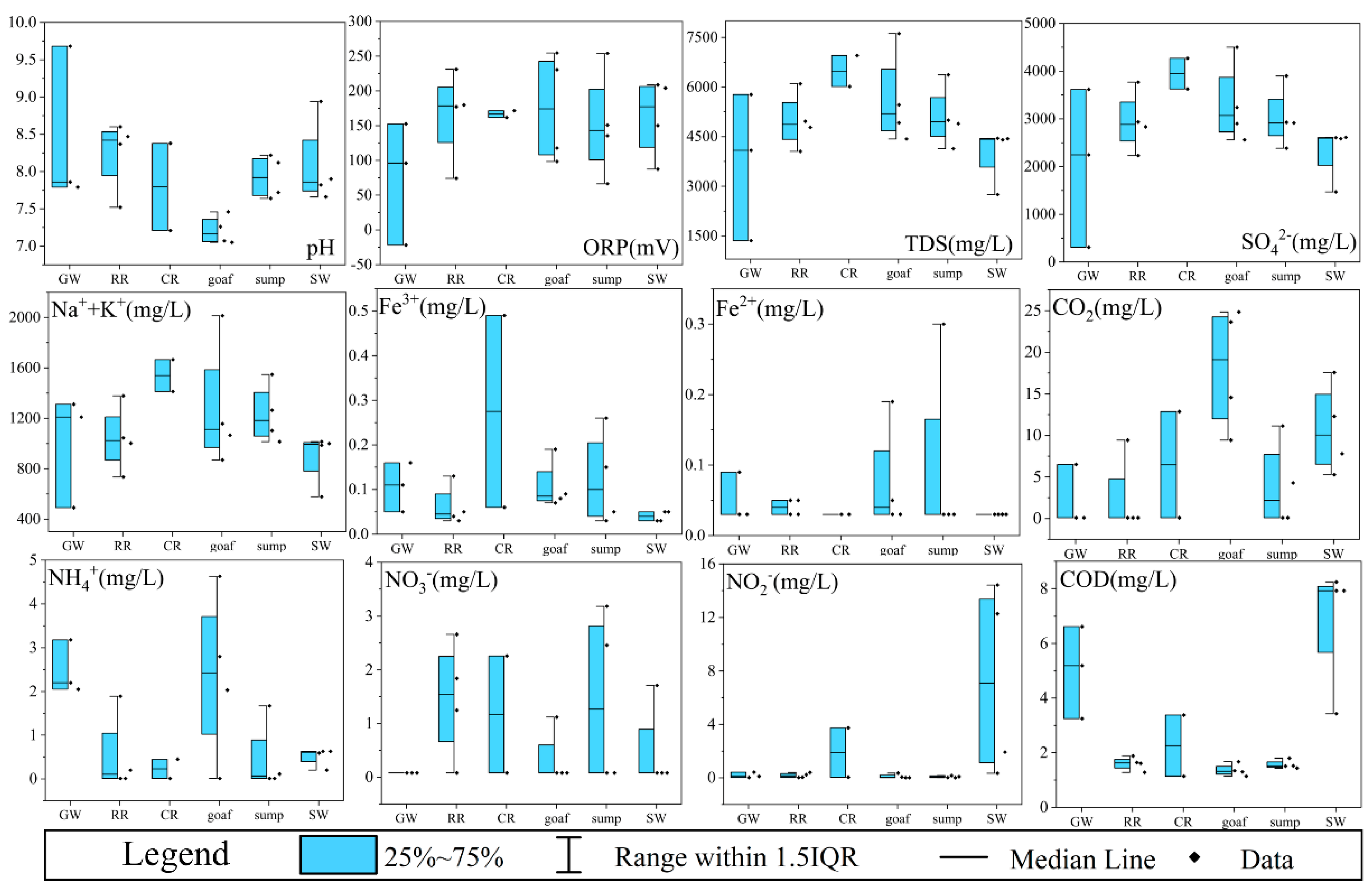
3.1. Spatial Changes in Hydrochemical Characteristic
3.2. Overall Microbial Diversity and Taxonomic Composition Variation across Different Zones
3.3. Relationships between Microbial Community Structure and Hydrochemical Components
3.4. Microbial Community Variation Mechanism across Four Typical Environmental Processes in Coal Mine
3.4.1. Vertical Distribution Characteristics of Microbial Communities in Groundwater Aquifers
3.4.2. Variation Characteristics of Microbial Communities during Coal Mining
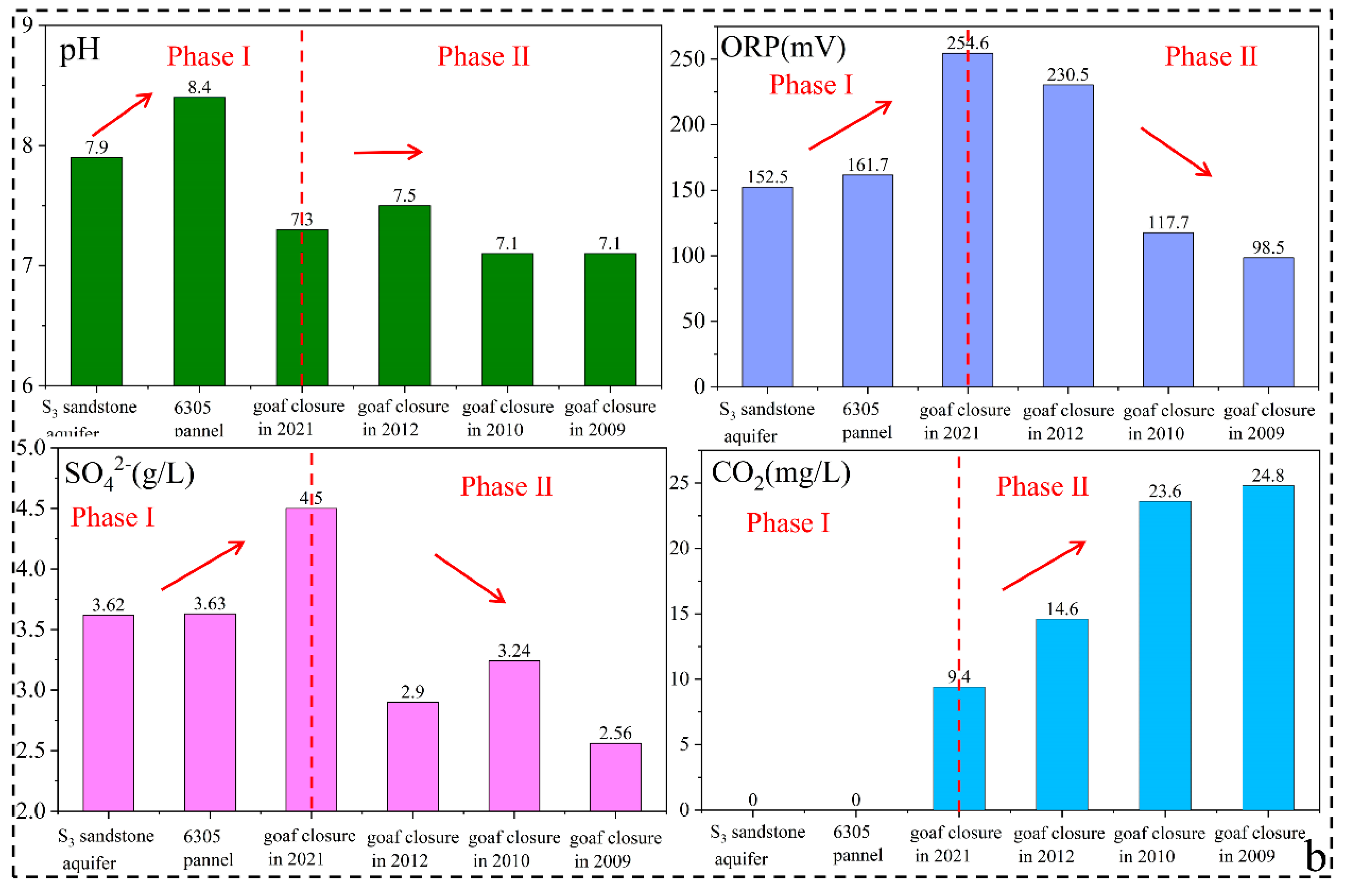
3.4.3. Variation Characteristics of Microbial Communities in Goafs with the Time since Cessation of Mining
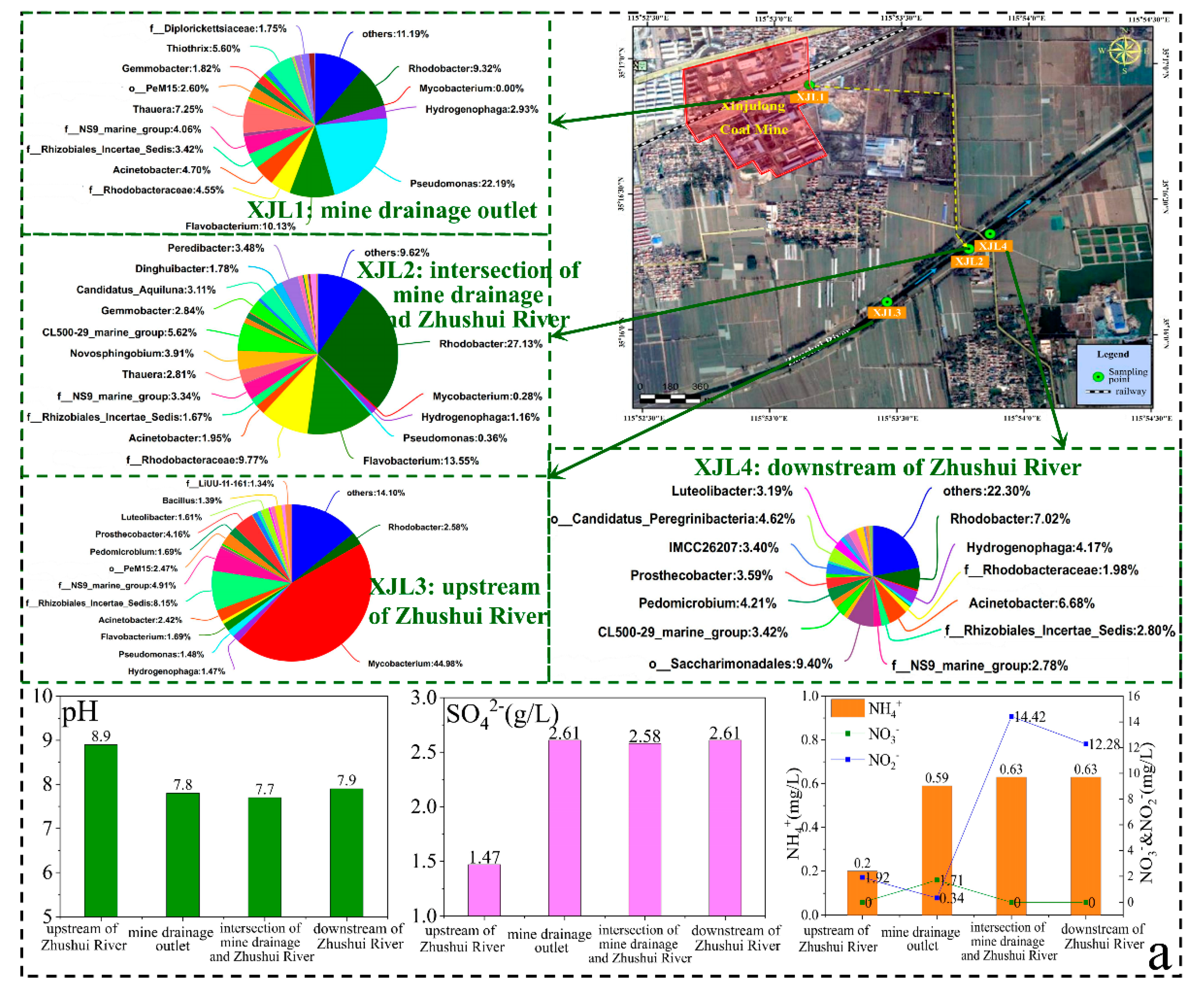
3.4.4. Effect of Mine Drainage on Microbial Community of the River
4. Conclusions
- (1)
- The hydrochemical types of mine waters inherited that of their recharge water source (SO4-Na type), but their hydrochemical compositions showed obvious zone-specific patterns. The microbial community richness of the six zones was ranked as follows: coal roadways > water sumps ≈ surface waters > rock roadways > goafs > groundwater aquifers, while the microbial community diversity of surface waters was the highest.
- (2)
- The microbial communities in the different zones were eminently different. Bacteria related to sulfur oxidation and hydrocarbon degradation (i.e., Thioclava and Dietzia) were more abundant in the coal roadways. Anyway, bacteria related to the SO42− reduction, hydrocarbon degradation, and nitrification (i.e., c_Thermodesulfovibrionia, Candidatus_Nitrotoga, and Nitrospira) accounted for higher proportions in goafs and rock roadways. Sphingobium and Candidatus desulforudis were almost exclusively distributed in groundwater aquifers, while aerobic Mycobacterium was the dominant species only in surface waters.
- (3)
- The RDA and heatmap analysis demonstrated that the microbial communities corresponded to the levels of redox sensitive indices (i.e., COD, NH4+, NO2−, ORP, SO42−, Fe3+, and Fe2+). Co-occurrence networks, with 88.71% positive correlations, confirmed that these microbial genera, related to the S, N, C, and Fe cycles, had significant correlation with each other and might drive the formation and evolution of hydrochemical characteristics.
- (4)
- Coal roadways and goafs were the critical zones of groundwater pollution prevention and control. During tunneling in the panel, pyrite could be oxidized by sulfur-oxidizing bacteria, leading to SO42− increase. With the closure of the panel and the formation of the goaf, the SO42− concentration increased rapidly for a short period. However, with the time since goaf closure, the proportion of SRB increased significantly, leading to SO42− concentration’s decrease by 42% over 12 years, which indicated that long-term closed goafs had a certain self-purification ability.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, H.B.; Zhou, J.W.; Chai, B.; Zhou, A.G.; Li, J.Z.; Zhu, H.H.; Chen, H.N.; Su, D.H. Groundwater environmental risk assessment of abandoned coal mine in each phase of the mine life cycle: A case study of Hongshan coal mine, North China. Environ. Sci. Pollut. Res. 2020, 27, 42001–42021. [Google Scholar] [CrossRef] [PubMed]
- Li, J.G.; Li, Z.X.; Brandis, K.J.; Bu, J.W.; Sun, Z.Y.; Yu, Q.; Ramp, D. Tracing geochemical pollutants in stream water and soil from mining activity in an alpine catchment. Chemosphere 2020, 242, 125167. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.Z.; Li, J.F.; Cao, Z.G.; Wu, B.Y.; Jiang, B.B.; Yang, Y.; Yang, J.; Chen, Y.P. Technology and engineering development strategy of water protection and utilization of coal mine in China. J. China Coal Soc. 2021, 46, 1–12. [Google Scholar]
- Ren, K.; Zeng, J.; Liang, J.P.; Yuan, D.X.; Jiao, Y.J.; Peng, C.; Pan, X.D. Impacts of acid mine drainage on karst aquifers: Evidence from hydrogeochemistry, stable sulfur and oxygen isotopes. Sci. Total Environ. 2021, 761, 143223. [Google Scholar] [CrossRef]
- Li, S.; Yu, L.F.; Jiang, W.J.; Yu, H.X.; Wang, X.M. The recent progress china has made in green mine construction, Part I: Mining groundwater pollution and sustainable mining. Int. J. Environ. Res. Public Health 2022, 19, 5673. [Google Scholar] [CrossRef]
- Luis, A.T.; Cordoba, F.; Antunes, C.; Loayza-Muro, R.; Grande, J.A.; Silva, B.; Diaz-Curiel, J.; Ferreira da Silva, E. Extremely acidic eukaryotic (micro) organisms: Life in acid mine drainage polluted environments-mini-review. Int. J. Environ. Res. Public Health 2022, 19, 376. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Ji, Y.; Li, C.R.; Luo, P.P.; Wang, W.K.; Zhang, Y.; Nover, D. Effects of Phytoremediation Treatment on Bacterial Community Structure and Diversity in Different Petroleum-Contaminated Soils. Int. J. Environ. Res. Public Health 2018, 15, 2168. [Google Scholar] [CrossRef] [Green Version]
- Meheust, R.; Castelle, C.J.; Carnevali, P.B.M.; Farag, I.F.; He, C.; Chen, L.X.; Amano, Y.; Hug, L.A.; Banfield, J.F. Groundwater Elusimicrobia are metabolically diverse compared to gut microbiome Elusimicrobia and some have a novel nitrogenase paralog. ISME J. 2020, 14, 2907–2922. [Google Scholar] [CrossRef]
- Zhang, H.S.; Wang, M.Y.; Cai, W.T.; Bian, C.; Liu, J.W. Characteristics of microbial community composition and environmental response in deep fluorinated groundwater. Earth Sci. 2021, 1, 1–12. [Google Scholar] [CrossRef]
- Brantner, J.S.; Senko, J.M. Response of soil-associated microbial communities to intrusion of coal mine-derived acid mine drainage. Environ. Sci. Technol. 2014, 48, 8556–8563. [Google Scholar] [CrossRef]
- de Quadros, P.D.; Zhalnina, K.; Davis-Richardson, A.G.; Drew, J.C.; Menezes, F.B.; Camargo, F.A.O.; Triplett, E.W. Coal mining practices reduce the microbial biomass, richness and diversity of soil. Appl. Soil Ecol. 2016, 98, 195–203. [Google Scholar] [CrossRef]
- Wang, H.; Guo, C.L.; Yang, C.F.; Lu, G.N.; Chen, M.Q.; Dang, Z. Distribution and diversity of bacterial communities and sulphate-reducing bacteria in a paddy soil irrigated with acid mine drainage. J. Appl. Microbiol. 2016, 121, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Gao, P.; Sun, X.X.; Xiao, E.Z.; Xu, Z.X.; Li, B.Q.; Sun, W.M. Characterization of iron-metabolizing communities in soils contaminated by acid mine drainage from an abandoned coal mine in Southwest China. Environ. Sci. Pollut. Res. 2019, 26, 9585–9598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Tang, S.; Wang, M.; Sun, W.M.; Xie, Y.W.; Peng, H.; Zhong, A.M.; Liu, H.L.; Zhang, X.W.; Yu, H.X.; et al. Acid mine drainage affects the diversity and metal resistance gene profile of sediment bacterial community along a river. Chemosphere 2019, 217, 790–799. [Google Scholar] [CrossRef]
- Jin, L.R.; Gerson, J.R.; Rocca, J.D.; Bernhardt, E.S.; Simonin, M. Alkaline mine drainage drives stream sediment microbial community structure and function. Sci. Total Environ. 2022, 805, 150189. [Google Scholar] [CrossRef]
- Bomberg, M.; Arnold, M.; Kinnunen, P. Characterization of the bacterial and sulphate reducing community in the alkaline and constantly cold water of the closed kotalahti mine. Minerals 2015, 5, 452–472. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.P.; Guo, C.L.; Wang, H.; Lu, G.N.; Yang, C.F.; Chen, M.Q.; Dang, Z. Fe- and S-metabolizing microbial communities dominate an AMD-contaminated river ecosystem and play important roles in Fe and S cycling. Geomicrobiol. J. 2017, 34, 695–705. [Google Scholar] [CrossRef]
- Kefeni, K.K.; Msagati, T.A.M.; Mamba, B.B. Acid mine drainage: Prevention, treatment options, and resource recovery: A review. J. Clean Prod. 2017, 151, 475–493. [Google Scholar] [CrossRef]
- Raudsepp, M.J.; Gagen, E.J.; Evans, P.; Tyson, G.W.; Golding, S.D.; Southam, G. The influence of hydrogeological disturbance and mining on coal seam microbial communities. Geobiology 2016, 14, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Ben Maamar, S.; Aquilina, L.; Quaiser, A.; Pauwels, H.; Michon-Coudouel, S.; Vergnaud-Ayraud, V.; Labasque, T.; Roques, C.; WAbbott, B.; Dufresne, A. Groundwater isolation governs chemistry and microbial community structure along hydrologic flowpaths. Front. Microbiol. 2015, 6, 1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B. Study on occurrence and biodegradation mechanism of PAHs in closed coal mine. PhD Thesis, China University of Mining and Technology, Xuzhou, China, 2019. [Google Scholar]
- Zhang, Y.T. Experimental study on the degradation of polycyclic aromatic hydrocarbons in groundwater by obligate mixed Bacteria. Master’s Thesis, China University of Mining and Technology, Xuzhou, China, 2019. [Google Scholar]
- Modi, E.; Ding, T.; Zhan, S.C.; Yao, F.W.; Wan, H.K. Effect of chlorine concentration on determining COD by acid and alkaline potassium permanganate. In Proceedings of the International Conference on Advances in Energy and Environmental Science (ICAEES), Guangzhou, China, 12–14 August 2013; Volume 807–809, pp. 232–235. [Google Scholar]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the Third International Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Sun, W.M.; Xiao, E.Z.; Krumins, V.; Dong, Y.R.; Li, B.Q.; Deng, J.; Wang, Q.; Xiao, T.F.; Liu, J. Comparative analyses of the microbial communities inhabiting coal mining waste dump and an adjacent acid mine drainage creek. Microb. Ecol. 2019, 78, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- An, X.L.; Baker, P.; Li, H.; Su, J.Q.; Yu, C.P.; Cai, C. The patterns of bacterial community and relationships between sulfate-reducing bacteria and hydrochemistry in sulfate-polluted groundwater of Baogang rare earth tailings. Environ. Sci. Pollut. Res. 2016, 23, 21766–21779. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.T.; Liu, T.; Chen, N.; Feng, C.P. Changes in microbial community diversity, composition, and functions upon nitrate and Cr(VI) contaminated groundwater. Chemosphere 2022, 288, 132476. [Google Scholar] [CrossRef]
- Jiang, B.H.; Zhang, B.; Li, L.; Zhao, Y.; Shi, Y.; Jiang, Q.; Jia, L.P. Analysis of microbial community structure and diversity in surrounding rock soil of different waste dump sites in fushun western opencast mine. Chemosphere 2021, 269, 128777. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Chen, Y.L.; Zhang, D.D.; Zhao, M.M.; Li, H.L.; Guo, P. Microbiome analysis reveals microecological balance in the emerging rice-crayfish integrated breeding mode. Front. Microbiol. 2021, 12, 1447. [Google Scholar] [CrossRef]
- Taubert, M.; Overholt, W.A.; Heinze, B.M.; Matanfack, G.A.; Houhou, R.; Jehmlich, N.; Von Bergen, M.; Roesch, P.; Popp, J.; Kuesel, K. Bolstering fitness via CO2 fixation and organic carbon uptake: Mixotrophs in modern groundwater. ISME J. 2022, 16, 1153–1162. [Google Scholar] [CrossRef]
- Jin, L.; Jin, C.Z.; Lee, H.G.; Lee, C.S. Genomic insights into denitrifying methane-oxidizing bacteria gemmobacter fulva sp. nov., isolated from an anabaena culture. Microorganisms 2021, 9, 2423. [Google Scholar] [CrossRef] [PubMed]
- Safonov, A.V.; Boguslavsky, A.E.; Gaskova, O.L.; Boldyrev, K.A.; Shvartseva, O.S.; Khvashchevskaya, A.A.; Popova, N.M. Biogeochemical modelling of uranium immobilization and aquifer remediation strategies near NCCP sludge storage facilities. Appl. Sci. 2021, 11, 2875. [Google Scholar] [CrossRef]
- Custodio, M.; Peñaloza, R.; Espinoza, C.; Espinoza, W.; Mezarina, J. Treatment of dairy industry wastewater using bacterial biomass isolated from eutrophic lake sediments for the production of agricultural water. Bioresour. Technol. Rep. 2022, 17, 100891. [Google Scholar] [CrossRef]
- Braun, B.; Richert, I.; Szewzyk, U. Detection of iron-depositing pedomicrobium species in native biofilms from the Odertal National Park by a new, specific FISH probe. J. Microbiol. Methods 2009, 79, 37–43. [Google Scholar] [CrossRef]
- Hu, J.W.; Zhang, J.S.; Li, Z.; Tao, W.Q. Enhanced methane yield through sludge two-phase anaerobic digestion process with the addition of calcium hypochlorite. Bioresour. Technol. 2022, 347, 126693. [Google Scholar] [CrossRef]
- Donovan, J.J.; Leavitt, B.R.; Werner, E. Long-term changes in water chemistry as a result of mine flooding in closed mines of the Pittsburgh Coal Basin, USA. In Proceedings of the 6th International Conference on Acid Rock Drainage, Cairns, Australia, 14–17 July 2003; pp. 869–875. [Google Scholar]
- Qian, H.; Ma, Z.Y. Hydrogeochemistry, 1st ed.; Geology Press: Beijing, China, 2005; p. 134. [Google Scholar]
- Sharp, C.E.; Brady, A.L.; Sharp, G.H.; Grasby, S.E.; Stott, M.B.; Dunfield, P.F. Humboldt’s spa: Microbial diversity is controlled by temperature in geothermal environments. ISME J. 2014, 8, 1166–1174. [Google Scholar] [CrossRef] [Green Version]
- Duan, P.P.; Song, Y.F.; Li, S.S.; Xiong, Z.Q. Responses of N2O production pathways and related functional microbes to temperature across greenhouse vegetable field soils. Geoderma 2019, 355, 113904. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Xu, Z.; Sun, Y.; Gao, Y.; Zhu, L. Coal Mining Activities Driving the Changes in Microbial Community and Hydrochemical Characteristics of Underground Mine Water. Int. J. Environ. Res. Public Health 2022, 19, 13359. https://doi.org/10.3390/ijerph192013359
Zhang L, Xu Z, Sun Y, Gao Y, Zhu L. Coal Mining Activities Driving the Changes in Microbial Community and Hydrochemical Characteristics of Underground Mine Water. International Journal of Environmental Research and Public Health. 2022; 19(20):13359. https://doi.org/10.3390/ijerph192013359
Chicago/Turabian StyleZhang, Li, Zhimin Xu, Yajun Sun, Yating Gao, and Lulu Zhu. 2022. "Coal Mining Activities Driving the Changes in Microbial Community and Hydrochemical Characteristics of Underground Mine Water" International Journal of Environmental Research and Public Health 19, no. 20: 13359. https://doi.org/10.3390/ijerph192013359