

Spatio-Temporal Variations of Zooplankton and Correlations with Environmental Parameters around Tiaowei Island, Fujian, China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
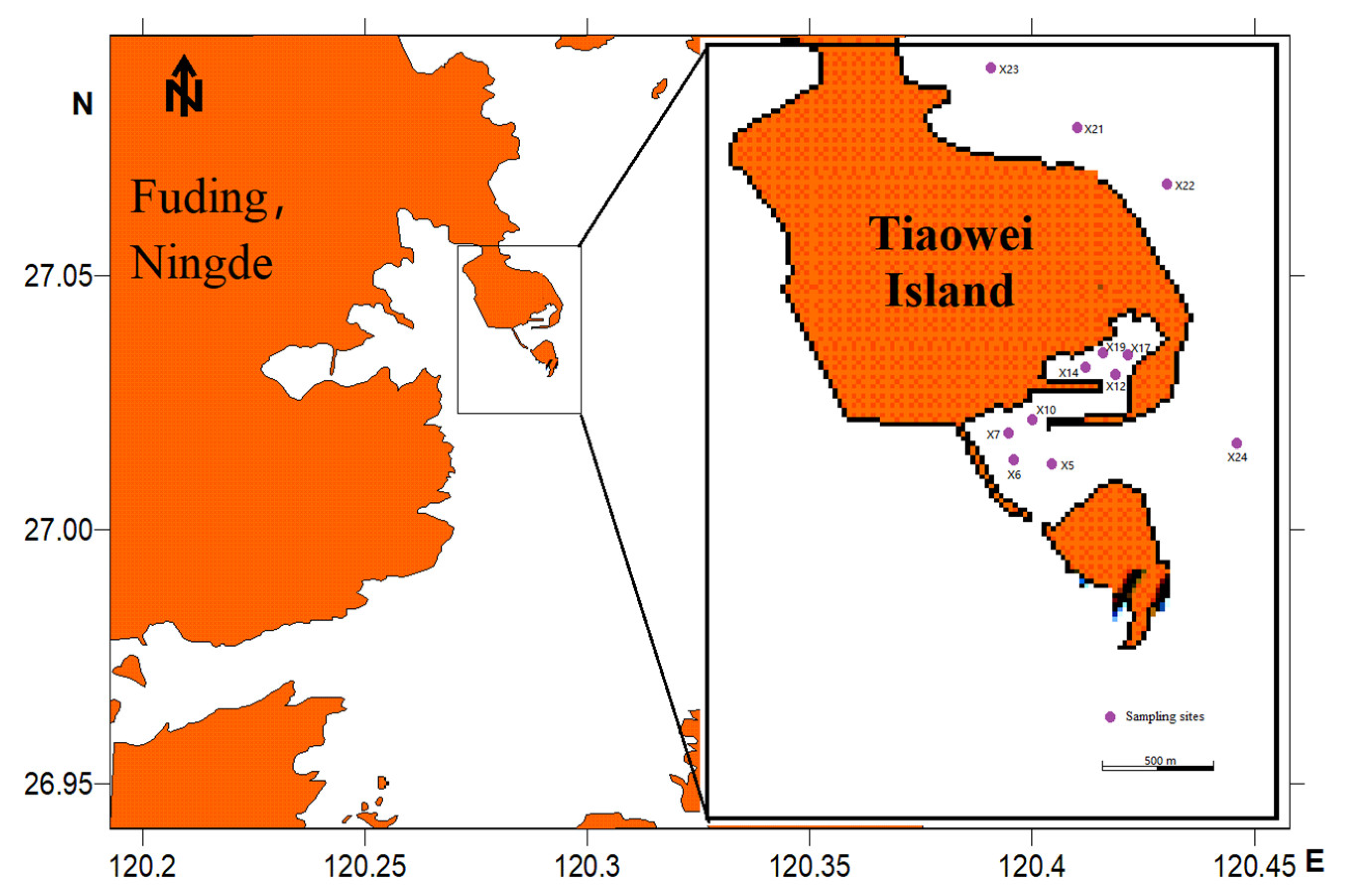
2.1. Sampling, Identification and Classification
2.2. Measurements of Environmental Variables
2.3. Statistical Analyses
3. Results
3.1. Environmental Factors
3.2. Species Composition and Diversity Index
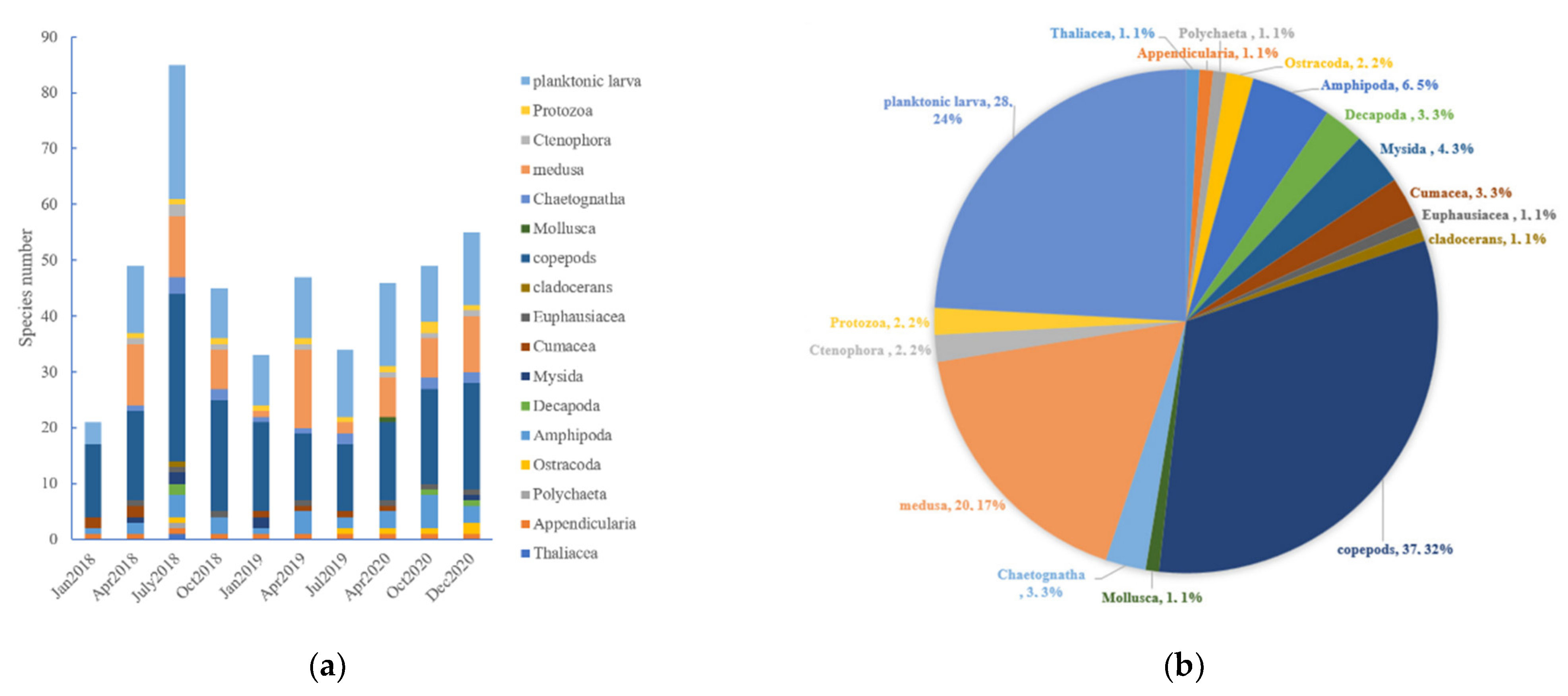
3.2.1. Zooplankton Composition
3.2.2. Variations in Abundance and Diversity Index
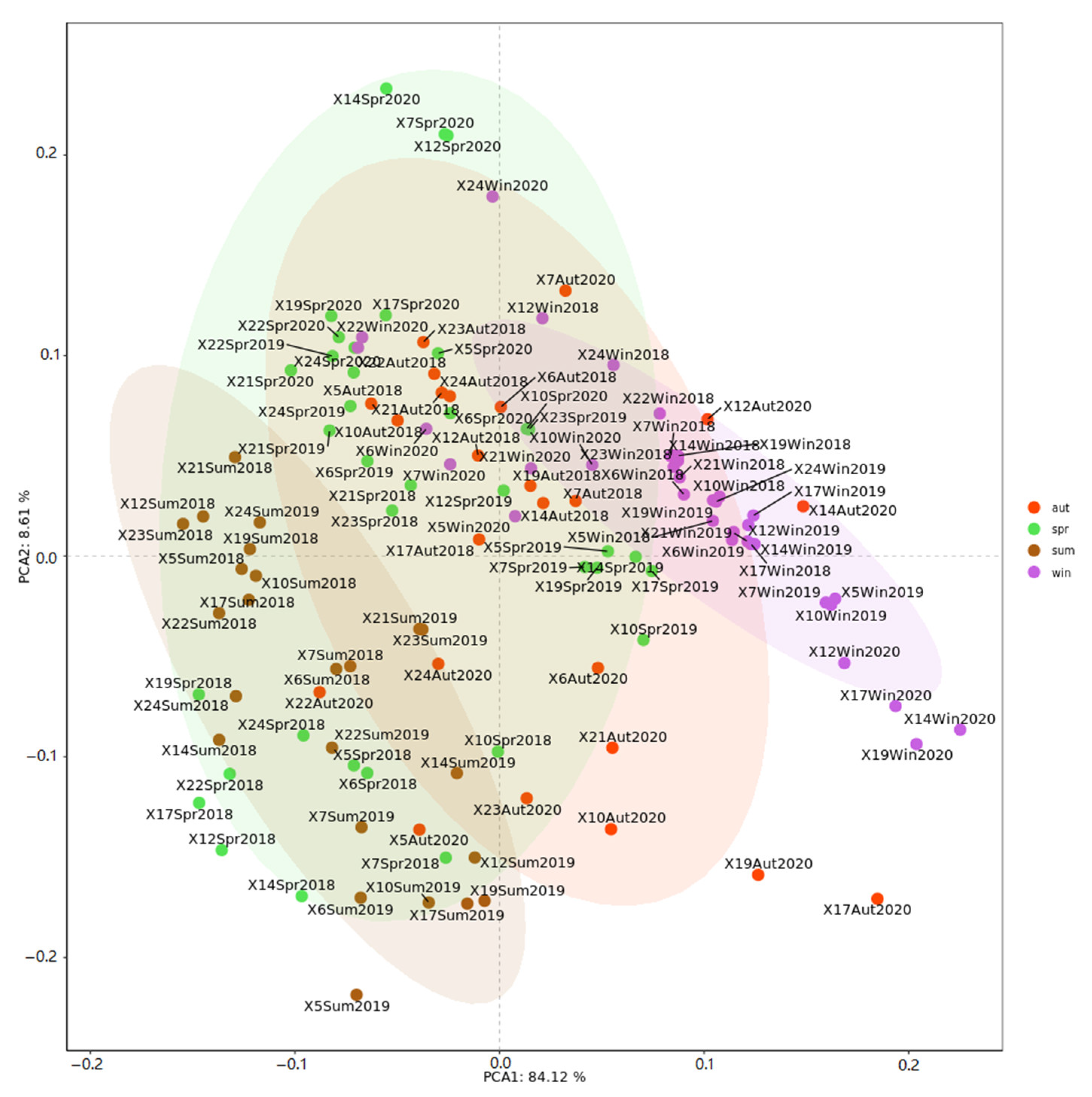
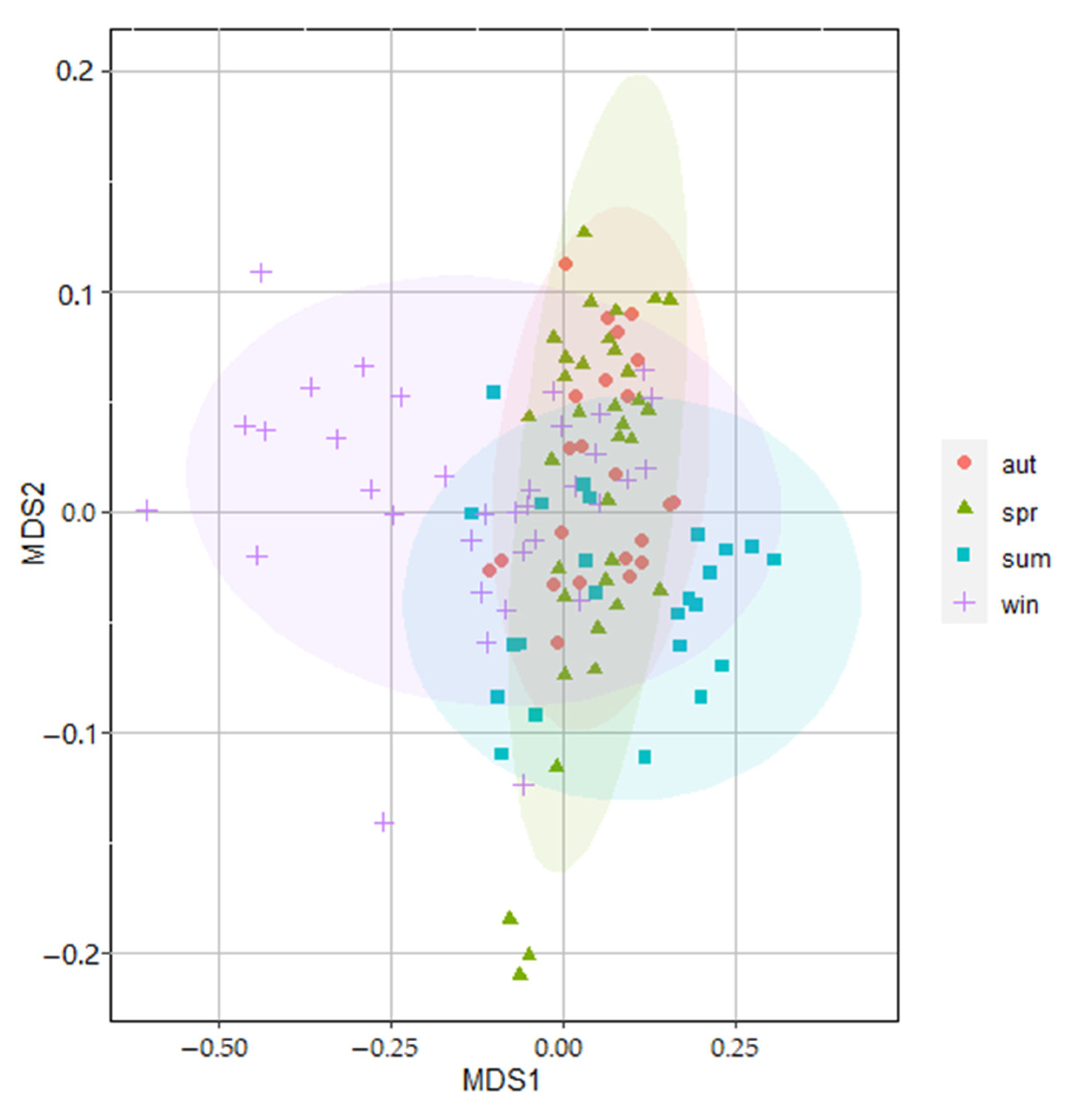
3.3. Spatial and Temporal Variations in Zooplankton Communities
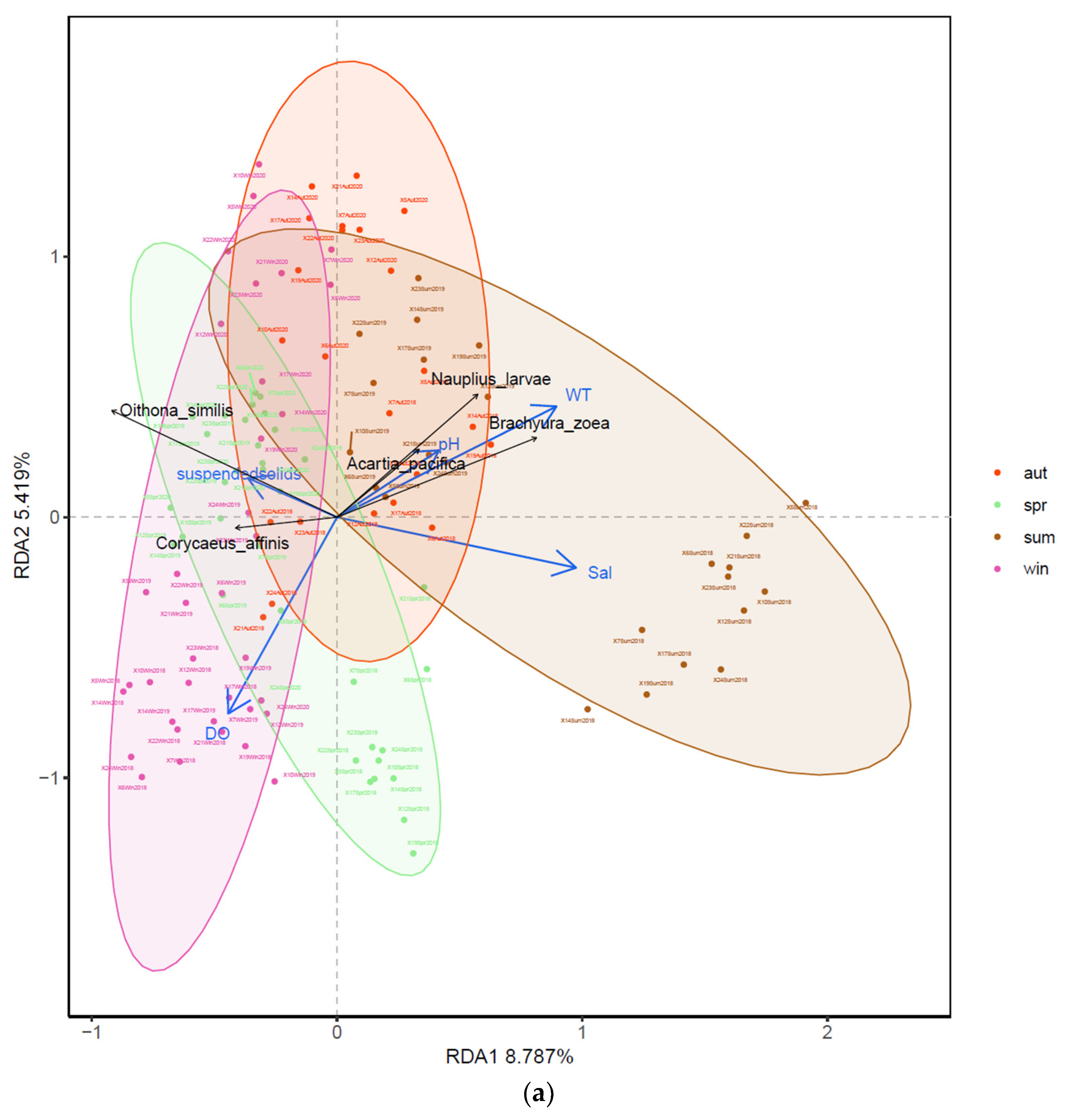
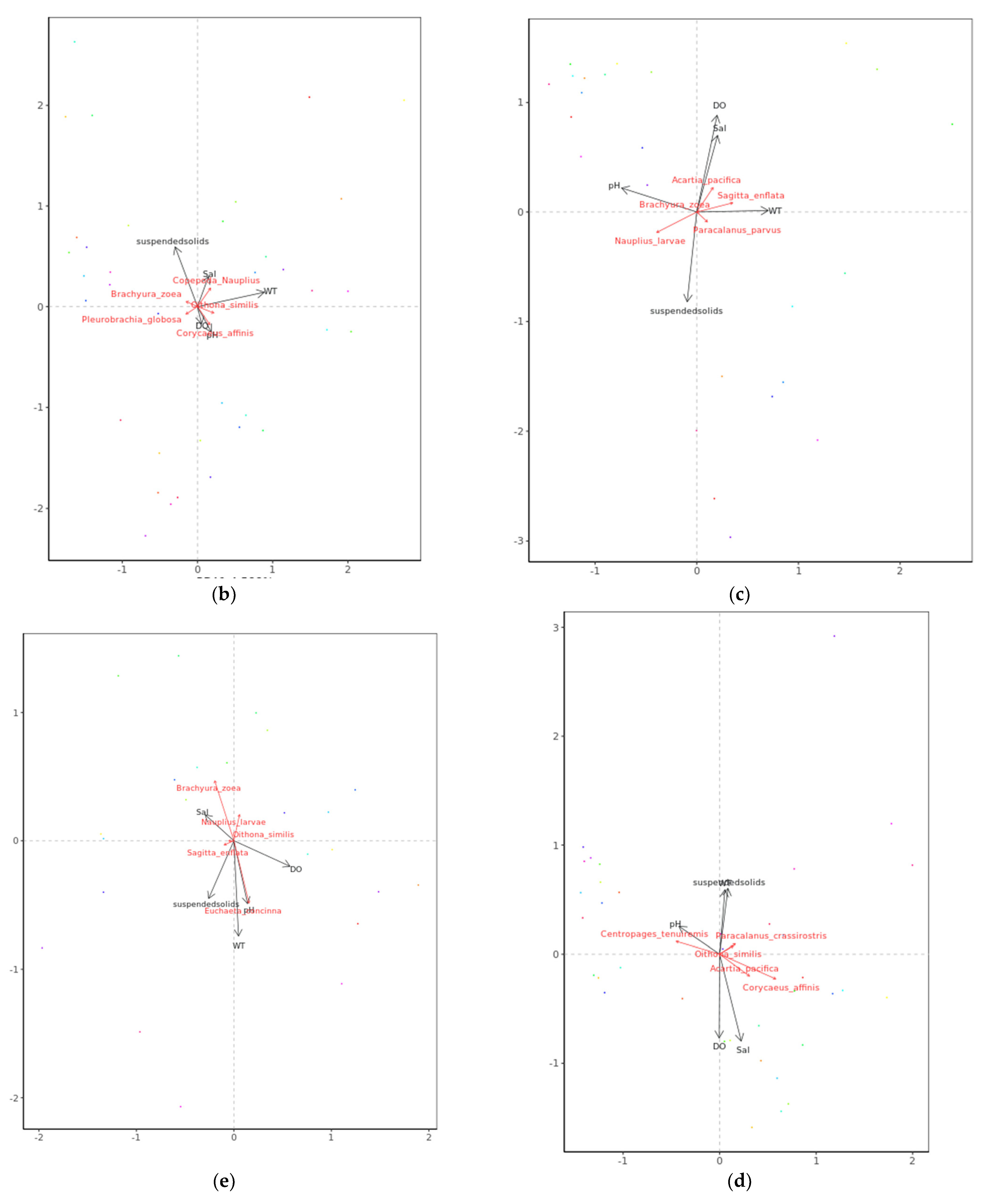
3.4. Relationships between Zooplankton Assemblage and Environmental Factors
4. Discussion
4.1. Species Composition
4.2. Variation in Diversity Indices
4.3. Spatial-Temporal Dynamics of Community Structure
4.4. Relationship between Environmental Parameters and Zooplankton
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, S.; Huo, Y.; Yang, B. Zooplankton functional groups on the continental shelf of the yellow sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1006–1016. [Google Scholar] [CrossRef]
- Bruggeman, J.; Kooijman, S.A. A biodiversity-inspired approach to aquatic ecosystem modeling. Limnol. Oceanogr. 2007, 52, 1533–1544. [Google Scholar] [CrossRef]
- Djurhuus, A.; Pitz, K.; Sawaya, N.A.; Rojas-Márquez, J.; Michaud, B.; Montes, E.; Muller-Karger, F.; Breitbart, M. Evaluation of marine zooplankton community structure through environmental DNA metabarcoding. Limnol. Oceanogr. Methods 2018, 16, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardyna, M.; Gosselin, M.; Michel, C.; Poulin, M.; Tremblay, J.É. Environmental forcing of phytoplankton community structure and function in the Canadian High Arctic: Contrasting oligotrophic and eutrophic regions. Mar. Ecol. Prog. Ser. 2011, 442, 37–57. [Google Scholar] [CrossRef] [Green Version]
- Nong, X.; Shao, D.; Shan, Y.; Liang, J. Analysis of spatio-temporal variation in phytoplankton and its relationship with water quality parameters in the South-to-North Water Diversion Project of China. Environ. Monit. Assess. 2021, 193, 593. [Google Scholar] [CrossRef]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Salvador, B.; Bersano, J.G.F. Zooplankton variability in the subtropical estuarine system of Paranaguá Bay, Brazil, in 2012 and 2013. Estuar. Coast. Shelf Sci. 2017, 199, 1–13. [Google Scholar] [CrossRef]
- Yebra, L.; Espejo, E.; Putzeys, S.; Giráldez, A.; Gómez-Jakobsen, F.; León, P.; Salles, S.; Torres, P.; Mercado, J.M. Zooplankton biomass depletion event reveals the importance of small pelagic fish top-down control in the Western Mediterranean coastal waters. Front. Mar. Sci. 2020, 7, 608690. [Google Scholar] [CrossRef]
- Griffith, G.P.; Hop, H.; Vihtakari, M.; Wold, A.; Kalhagen, K.; Gabrilsen, G.W. Ecological resilience of Arctic marine food webs to climate change. Nat. Clim. Change 2019, 9, 868–872. [Google Scholar] [CrossRef]
- Shemi, A.; Alcolombri, U.; Schatz, D.; Farstey, V.; Vincent, F.; Rotkopf, S.; Ben-Dor, S.; Frada, M.J.; Tawfik, D.S.; Vardi, S. Dimethyl sulfide mediates microbial predator–prey interactions between zooplankton and algae in the ocean. Nat. Microbiol. 2021, 6, 1357–1366. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Papathanassiou, E.; Lepretre, A.; Frontier, S. Zooplankton Assemblages and Influence of EnvironmentalParameters on Them in a Mediterranean Coastal Area. J. Plankton Res. 1998, 20, 847–870. [Google Scholar] [CrossRef] [Green Version]
- Marcus, N. An Overview of the Impacts of Eutrophication and Chemical Pollutants on Copepods of the Coastal Zone. Zool Stud 2004, 43, 211–217. [Google Scholar]
- Zhang, K.; Jiang, F.; Chen, H.; Dibar, D.T.; Wu, Q.; Zhou, Z. Temporal and spatial variations in zooplankton communities in relation to environmental factors in four floodplain lakes located in the middle reach of the Yangtze River, China. Environ. Pollut. 2019, 251, 277–284. [Google Scholar] [CrossRef]
- Annabi-Trabelsi, N.; Guermazi, W.; Leignel, V.; Al-Enezi, Y.; Karam, Q.; Ali, M.; Ayadi, H.; Belmonte, G. Effects of Eutrophication on Plankton Abundance and Composition in the Gulf of Gabès (Mediterranean Sea, Tunisia). Water 2022, 14, 2230. [Google Scholar] [CrossRef]
- Rose, T.H.; Tweedley, J.R.; Warwick, R.M.; Potter, I.C. Influences of microtidal regime and eutrophication on estuarine zooplankton. Estuar. Coast. Shelf Sci. 2020, 238, 106689. [Google Scholar] [CrossRef]
- Havas, M.; Rosseland, B.O. Response of zooplankton, benthos, and fish to acidification: An overview. Water Air Soil Pollut. 1995, 85, 51–62. [Google Scholar] [CrossRef]
- Fischer, J.M.; Frost, T.M.; Ives, A.R. Compensatory dynamics in zooplankton community responses to acidification: Measurement and mechanisms. Ecol. Appl. 2001, 11, 1060–1072. [Google Scholar] [CrossRef]
- Williamson, C.E.; Grad, G.; De Lange, H.J.; Gilroy, S.; Karapelou, D.M. Temperature-dependent ultraviolet responses in zooplankton: Implications of climate change. Limnol. Oceanogr. 2002, 47, 1844–1848. [Google Scholar] [CrossRef]
- Brodeur, R.D.; Auth, T.D.; Phillips, A.J. Major shifts in pelagic micronekton and macrozooplankton community structure in an upwelling ecosystem related to an unprecedented marine heatwave. Front. Mar. Sci. 2019, 6, 212. [Google Scholar] [CrossRef] [Green Version]
- Hylander, S.; Hansson, L.A. Vertical migration mitigates UV effects on zooplankton community composition. J. Plankton Res. 2010, 32, 971–980. [Google Scholar] [CrossRef]
- Liu, W.; Yu, Z.; Zhang, Z.; Lin, J.; Cheng, H.; Fan, L.; Wang, F.; Mu, J. Fish assemblage structure and spatio-temporal variation in Qingchuan bay, Ningde, Fujian. Mar. Environ. Sci. 2022, 41, 738–744. (In Chinese) [Google Scholar]
- Tang, Y.; Wang, J.; Cheng, H.; Zheng, B.; Ma, Z. Eco-environment quality assessment of macrobenthic community in the East Ningde sea waters. Mar. Environ. Sci. 2019, 38, 278–285+302. (In Chinese) [Google Scholar] [CrossRef]
- Calbet, A.; Landry, M.R. Mesozooplankton influences on the microbial food web: Direct and indirect trophic interactions in the oligotrophic open ocean. Limnol. Oceanogr. 1999, 44, 1370–1380. [Google Scholar] [CrossRef]
- Deng, B.; Liu, H.; Wang, H.; Qin, Y.; Xia, L.; Li, Y.; Zhang, H.; Jiang, X.; Yang, Y.; Wang, Y.; et al. Analysis on the community characteristics and potential ecological risk of jellyfish in the Qingchuan Bay of Ningde, Fujian Province. Acta Oceanol. Sin. 2020, 42, 128–136. (In Chinese) [Google Scholar] [CrossRef]
- Sun, S.; Li, C.; Cheng, F.; Jin, X.; Yang, B. Atlas of Common Zooplankton of the Chinese Coastal Seas; Ocean Press: Beijing, China, 2015. [Google Scholar]
- Rizzo, L.; Pusceddu, A.; Bianchelli, S.; Fraschetti, S. Potentially combined effect of the invasive seaweed Caulerpa cylindracea (Sonder) and sediment deposition rates on organic matter and meiofaunal assemblages. Mar. Environ. Res. 2020, 159, 104966. [Google Scholar] [CrossRef]
- Sartory, D.P.; Grobbelaar, J.U. Extraction of chlorophyll a from freshwater phytoplankton for spectrophotometric analysis. Hydrobiologia 1984, 114, 177–187. [Google Scholar] [CrossRef]
- Sun, D.; Li, Y.; Le, C.; Shi, K.; Huang, C.; Gong, S.; Yin, B. A semi-analytical approach for detecting suspended particulate composition in complex turbid inland waters (China). Remote Sens. Environ. 2013, 134, 92–99. [Google Scholar] [CrossRef]
- Da Silva, A.M.V.; da Silva, R.J.B.; Camoes, M.F.G. Optimization of the determination of chemical oxygen demand in wastewaters. Anal. Chim. Acta 2011, 699, 161–169. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Oksanen, M.J.; Solymos, P.; Wagner, H.; Szoecs, E.; et al. Package ‘vegan’. Community ecology package, version. 2013, pp. 1–295. Available online: https://cran.ism.ac.jp/web/packages/vegan/vegan.pdf (accessed on 17 April 2022).
- Anderson, M.J.; Walsh, D.C. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Chen, C.S.; Chiu, T.S. Composition and abundance of copepods and ichthyoplankton in Taiwan Strait (western North Pacific) are influenced by seasonal monsoons. Mar. Freshw. Res. 2005, 56, 153–161. [Google Scholar] [CrossRef]
- Hsiao, S.H.; Kâ, S.; Fang, T.H.; Hwang, J.S. Zooplankton assemblages as indicators of seasonal changes in water masses in the boundary waters between the East China Sea and the Taiwan Strait. Hydrobiologia 2011, 666, 317–330. [Google Scholar] [CrossRef]
- Lan, Y.C.; Lee, M.A.; Liao, C.H.; Lee, K.T. Copepod community structure of the winter frontal zone induced by the Kuroshio Branch Current and the China Coastal Current in the Taiwan Strait. J. Mar. Sci. Technol. 2009, 17, 1–6. [Google Scholar] [CrossRef]
- Tseng, L.C.; Souissi, S.; Dahms, H.U.; Chen, Q.C.; Hwang, J.S. Copepod communities related to water masses in the southwest East China Sea. Helgol. Mar. Res. 2008, 62, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Liu, T.; Zhu, M.; Liang, J.; Zhao, Y.; Zhang, B. Retention and characteristics of microplastics in natural zooplankton taxa from the East China Sea. Sci. Total Environ. 2018, 640, 232–242. [Google Scholar] [CrossRef]
- Xiang, C.; Ke, Z.; Li, K.; Liu, J.; Zhou, L.; Lian, X.; Tan, Y. Effects of terrestrial inputs and seawater intrusion on zooplankton community structure in Daya Bay, South China Sea. Mar. Pollut. Bull. 2021, 167, 112331. [Google Scholar] [CrossRef]
- Nandy, T.; Mandal, S. Unravelling the spatio-temporal variation of zooplankton community from the river Matla in the Sundarbans Estuarine System, India. Oceanologia 2020, 62, 326–346. [Google Scholar] [CrossRef]
- Basu, S.; Gogoi, P.; Bhattacharyya, S.; Das, S.K.; Das, B.K. Variability in the zooplankton assemblages in relation to environmental variables in the tidal creeks of Sundarbans estuarine system, India. Environ. Sci. Pollut. Res. 2022, 29, 45981–46002. [Google Scholar] [CrossRef]
- Omori, M.; Ishii, H.; Fujinaga, A. Life history strategy of Aurelia aurita (Cnidaria, Scyphomedusae) and its impact on the zooplankton community of Tokyo Bay. ICES J. Mar. Sci. 1995, 52, 597–603. [Google Scholar] [CrossRef]
- Souissi, S.; Yahia-Kefi, O.D.; Yahia, M.N.D. Spatial characterization of nutrient dynamics in the Bay of Tunis (south-western Mediterranean) using multivariate analyses: Consequences for phyto- and zooplankton distribution. J. Plankton Res. 2000, 22, 2039–2059. [Google Scholar] [CrossRef] [Green Version]
- Keister, J.E.; Peterson, W.T. Zonal and seasonal variations in zooplankton community structure off the central Oregon coast, 1998–2000. Prog. Oceanogr. 2003, 57, 341–361. [Google Scholar] [CrossRef]
- Vieira, L.; Azeiteiro, U.; Ré, P.; Pastorinho, R.; Marques, J.C.; Morgado, F. Zooplankton distribution in a temperate estuary (Mondego estuary southern arm: Western Portugal). Acta Oecol. -Int J. Ecol. 2003, 24, S163–S173. [Google Scholar] [CrossRef]
- Paffenhofer, G. On the ecology of marine cyclopoid copepods (Crustacea, Copepoda). J. Plankton Res. 1993, 15, 37–55. [Google Scholar] [CrossRef]
- Gallienne, C.P.; Robins, D.B. Is Oithona the most important copepod in the world’s oceans? J. Plankton Res. 2001, 23, 1421–1432. [Google Scholar] [CrossRef]
- Hamil, S.; Bouchelouche, D.; Arab, S.; Alili, M.; Baha, M.; Arab, A. The relationship between zooplankton community and environmental factors of Ghrib Dam in Algeria. Environ. Sci. Pollut. Res. 2021, 28, 46592–46602. [Google Scholar] [CrossRef]
- Hamner, W.M.; Dawson, M.N. A review and synthesis on the systematics and evolution of jellyfish blooms: Advantageous aggregations and adaptive assemblages. Hydrobiologia 2009, 616, 161–191. [Google Scholar] [CrossRef]
- Uye, S.I. Human forcing of the copepod-fish-jellyfish triangular trophic relationship. Hydrobiologia 2011, 666, 71–83. [Google Scholar] [CrossRef]
- Lee, C.S.; O’Bryen, P.J.; Marcus, N.H. Copepods in Aquaculture; John Wiley & Sons: New York, NY, USA, 2008. [Google Scholar]
- Yasuda, T.; Kitajima, S.; Hayashi, A.; Takahashi, M.; Fukuwaka, M.A. Cold offshore area provides a favorable feeding ground with lipid-rich foods for juvenile Japanese sardine. Fish. Oceanogr. 2021, 30, 455–470. [Google Scholar] [CrossRef]
- Coma, R.; Ribes, M.; Gili, J.M.; Zabala, M. Seasonality in coastal benthic ecosystems. Trends Ecol. Evol. 2000, 15, 448–453. [Google Scholar] [CrossRef]
- Duffy, M.A.; Hall, S.R.; Tessier, A.J.; Huebner, M. Selective predators and their parasitized prey: Are epidemics in zooplankton under top-down control? Limnol. Oceanogr. 2005, 50, 412–420. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, Y.; Fu, C.; Wang, B.; Li, J.; Ren, Y.; Wan, R. Variability of coastal cephalopods in overexploited China Seas under climate change with implications on fisheries management. Fish. Res. 2018, 208, 22–33. [Google Scholar] [CrossRef]
- Savchuk, O.P. Nutrient biogeochemical cycles in the Gulf of Riga: Scaling up field studies with a mathematical model. J. Mar. Syst. 2002, 32, 253–280. [Google Scholar] [CrossRef]
- Tett, P.; Hydes, D.; Sanders, R. Influence of nutrient biogeochemistry on the ecology of northwest European shelf seas. In Biogeochemistry of Marine Systems; Black, K.D., Shimmield, G.B., Eds.; Blackwell: Boca Raton, FL, USA, 2003; pp. 293–363. [Google Scholar]
- Abdul, W.O.; Adekoya, E.O.; Ademolu, K.O.; Omoniyi, I.T.; Odulate, D.O.; Akindokun, T.E.; Olajidi, A.E. The effects of environmental parameters on zooplankton assemblages in tropical coastal estuary, south-west, Nigeria. Egypt. J. Aquat. Res. 2016, 42, 281–287. [Google Scholar] [CrossRef]
- Liu, R.H. Study on the Status of Environment in Sea Reclamation of Ningde Nuclear Power Station. Master’s Thesis, Jimei University, Xiamen, China, 2013. [Google Scholar]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Madin, E.M.; Dill, L.M.; Ridlon, A.D.; Heithaus, M.R.; Warner, R.R. Human activities change marine ecosystems by altering predation risk. Glob. Chang Biol. 2016, 22, 44–60. [Google Scholar] [CrossRef]
- Borgwardt, F.; Robinson, L.; Trauner, D.; Teixeira, H.; Nogueira, A.J.; Lillebø, A.I.; Piet, G.; Kuemmerlen, M.; O’Higgins, T.; McDonald, H.; et al. Exploring variability in environmental impact risk from human activities across aquatic ecosystems. Sci. Total Environ. 2019, 652, 1396–1408. [Google Scholar] [CrossRef]
- Jakhar, P. Role of phytoplankton and zooplankton as health indicators of aquatic ecosystem: A review. Int. J. Innov. Res. Stud. 2013, 2, 489–500. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Jan-2018 | Apr-2018 | Jul-2018 | Oct-2018 | Jan-2019 | Apr-2019 | Jul-2019 | Apr-2020 | Oct-2020 | Dec-2020 | |
|---|---|---|---|---|---|---|---|---|---|---|
| WT (℃) | 13.08 ± 1.21 (12.2~15.8) | 20.12 ± 2.11 (17.9~24.8) | 31.40 ± 2.16 (29.8~37.6) | 20.88 ± 0.67 (20.1~21.8) | 12.76 ± 0.41 (12.4~13.6) | 17.32 ± 0.88 (16.4~19.5) | 30.36 ± 0.43 (29.8~31.5) | 18.33 ± 1.47 (17.3~22.6) | 24.68 ± 1.90 (23.6~28.8) | 19.87 ± 3.21 (16.0~24.7) |
| Sal (PSU) | 26.90 ± 0.15(26.65~27.25) | 29.46 ± 0.20(29.11~29.72) | 32.48 ± 0.22 (32.22~32.86) | 27.96 ± 0.32 (27.41~28.40) | 27.17 ± 0.20 (27.03~27.78) | 27.55 ± 0.16 (27.22~27.72) | 30.60 ± 0.16 (30.35~30.86) | 27.70 ± 0.14 (27.24~27.78) | 28.02 ± 0.37 (27.50~28.41) | 25.94 ± 0.04 (25.87~26.02) |
| pH | 8.04 ± 0.02 (8.00~8.06) | 8.13 ± 0.03 (8.08~8.17) | 8.15 ± 0.04 (8.10~8.22) | 7.99 ± 0.04 (7.94~8.06) | 8.10 ± 0.01 (8.08~8.11) | 8.07 ± 0.04 (8.03~8.14) | 8.13 ± 0.02 (8.10~8.16) | 8.11 ± 0.02 (8.07~8.13) | 8.12 ± 0.01 (8.10~8.13) | 8.11 ± 0.07 (7.97~8.20) |
| DO (μmol·L−1) | 9.09 ± 0.15 (8.81~9.30) | 8.09 ± 0.36 (7.50~8.60) | 7.52 ± 0.77 (6.36~8.65) | 7.17 ± 0.25 (6.73~7.46) | 8.82 ± 0.10 (8.64~8.94) | 7.88 ± 0.38 (7.40~8.58) | 5.81 ± 0.25 (5.53~6.37) | 7.86 ± 0.12 (7.73~8.14) | 7.23 ± 0.34 (6.38~7.84) | 6.84 ± 0.16 (6.54~7.17) |
| COD (mg·L−1) | 1.18 ± 0.26 (0.89~1.94) | 0.78 ± 0.11 (0.62~0.97) | 0.95 ± 0.24 (0.63~1.39) | 0.75 ± 0.25 (0.50~1.42) | 1.13 ± 0.26 (0.75~1.56) | 0.84 ± 0.32 (0.54~1.75) | 0.85 ± 0.20 (0.48~1.16) | 0.71 ± 0.10 (0.59~0.86) | 1.10 ± 0.32 (0.5~1.6) | 0.94 ± 0.47 (0.47~1.76) |
| PO4—P (nmol·L−1) | 0.026 ± 0.009(0.014~0.043) | 0.014 ± 0.004 (0.008~0.021) | 0.017 ± 0.004 (0.009~0.022) | 0.031 ± 0.011 (0.015~0.047) | 0.055 ± 0.010 (0.047~0.074) | 0.021 ± 0.008 (0.012~0.033) | 0.015 ± 0.002 (0.013~0.019) | 0.033 ± 0.001 (0.030~0.035) | 0.021 ± 0.003 (0.017~0.027) | 0.032 ± 0.006 (0.026~0.040) |
| NO2—N (nmol·L−1) | 0.007 ± 0.003 (0.003~0.017) | 0.007 ± 0.001 (0.005~0.009) | 0.010 ± 0.004 (0.001~0.017) | 0.008 ± 0.00 (0.005~0.01) | 0.005 ± 0.001 (0.004~0.006) | 0.008 ± 0.004 (0.003~0.014) | 0.017 ± 0.002 (0.015~0.020) | 0.016 ± 0.001 (0.014~0.018) | 0.010 ± 0.005 (0.006~0.018) | 0.008 ± 0.002 (0.006~0.013) |
| NO3—N (nmol·L−1) | 0.35 ± 0.11 (0.19~0.56) | 0.22 ± 0.06 (0.12~0.30) | 0.07 ± 0.02 (0.03~0.10) | 0.36 ± 0.14 (0.18~0.57) | 0.54 ± 0.02 (0.51~0.56) | 0.33 ± 0.1178 (0.18~0.48) | 0.10 ± 0.00 (0.09~0.12) | 0.52 ± 0.02 (0.48~0.55) | 0.23 ± 0.05 (0.15~0.31) | 0.61 ± 0.02 (0.59~0.64) |
| NH4—N (nmol·L−1) | 0.026 ± 0.014 (0.013~0.065) | 0.036 ± 0.035 (0.013~0.114) | 0.021 ± 0.006 (0.015~0.039) | 0.014 ± 0.004 (0.007~0.020) | 0.039 ± 0.016 (0.022~0.079) | 0.031 ± 0.013 (0.018~0.063) | 0.015 ± 0.005 (0.008~0.024) | 0.031 ± 0.009 (0.018~0.048) | 0.031 ± 0.017 (0.017~0.080) | 0.048 ± 0.016 (0.018~0.069) |
| DIN (nmol·L−1) | 0.39 ± 0.11 (0.21~0.60) | 0.27 ± 0.09 (0.16~0.27) | 0.10 ± 0.03 (0.05~0.15) | 0.38 ± 0.14 (0.19~0.60) | 0.59 ± 0.02 (0.56~0.65) | 0.37 ± 0.12 (0.21~0.52) | 0.13 ± 0.01 (0.11~0.16) | 0.57 ± 0.02 (0.52~0.59) | 0.27 ± 0.06 (0.19~0.39) | 0.67 ± 0.01 (0.63~0.69) |
| SiO4-Si (nmol·L−1) | 0.77 ± 0.25 (0.38~1.27) | 0.46 ± 0.11 (0.27~0.62) | 0.47 ± 0.09 (0.26~0.58) | 0.77 ± 0.34 (0.25~1.22) | 1.30 ± 0.03 (1.22~1.33) | 0.66 ± 0.025 (0.34~0.98) | 0.66 ± 0.06 (0.54~0.76) | NA | NA | NA |
| Chl-a (μg·L−1) | 0.45 ± 0.15 (0.24~0.68) | 5.82 ± 2.20 (1.36~9.00) | 3.03 ± 1.12 (1.17~5.67) | 0.81 ± 0.29 (0.44~1.56) | 0.61 ± 0.01 (0.46~0.77) | 1.37 ± 0.41 (0.56~2.24) | 5.69 ± 2.53 (1.8~10.33) | 0.93 ± 0.18 (0.68~1.15) | 5.02 ± 1.42 (2.73~6.62) | 1.03 ± 0.36 (0.65~1.56) |
| ss (mg·L−1) | 93.69 ± 20.39 (48.2~133.0) | 26.92 ± 12.89 (11.6~53.4) | 19.43 ± 6.03 (12.4~33.2) | 44.60 ± 12.65 (27.0~69.2) | 144.39 ± 38.42 (96.0~212.3) | 57.20 ± 27.779 (20.8~95.2) | 43.85 ± 12.81 (18.8~63.0) | 29.40 ± 9.21 (18.4~52.8) | 119.93 ± 100.3 (25.6~383.2) | 152.43 ± 173.1 (25.6~528.7) |
| (a) | Jan-2018 | Apr-2018 | Jul-2018 | Oct-2018 | Jan-2019 | Apr-2019 | Jul-2019 | Apr-2020 | Oct-2020 | Dec-2020 | ||
| S | 22 | 43 | 80 | 45 | 33 | 40 | 34 | 46 | 46 | 55 | ||
| D | 4.702 | 7.883 | 12.411 | 8.059 | 6.290 | 7.084 | 6.375 | 8.329 | 8.408 | 9.987 | ||
| J′ | 0.9039 | 0.9303 | 0.9156 | 0.9354 | 0.9165 | 0.9399 | 0.9173 | 0.9331 | 0.9350 | 0.9142 | ||
| H’ | 2.794 | 3.499 | 4.012 | 3.561 | 3.205 | 3.467 | 3.235 | 3.572 | 3.580 | 3.664 | ||
| (b) | X5 | X6 | X7 | X10 | X12 | X14 | X17 | X19 | X21 | X22 | X23 | X24 |
| S | 78 | 65 | 69 | 73 | 69 | 71 | 66 | 63 | 64 | 61 | 68 | 54 |
| D | 14.44 | 12.20 | 12.92 | 13.64 | 13.27 | 13.52 | 12.56 | 12.09 | 12.16 | 11.67 | 12.92 | 11.28 |
| J’ | 0.9489 | 0.9381 | 0.9456 | 0.9476 | 0.9481 | 0.9367 | 0.9370 | 0.9472 | 0.9426 | 0.9354 | 0.9324 | 0.9593 |
| H’ | 4.134 | 3.916 | 4.004 | 4.066 | 4.014 | 3.993 | 3.926 | 3.925 | 3.920 | 3.846 | 3.934 | 3.827 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Shi, Z.; Yu, Z.; Zhou, K.; Lin, J.; Wu, J.; Mu, J. Spatio-Temporal Variations of Zooplankton and Correlations with Environmental Parameters around Tiaowei Island, Fujian, China. Int. J. Environ. Res. Public Health 2022, 19, 12731. https://doi.org/10.3390/ijerph191912731
Zhang Z, Shi Z, Yu Z, Zhou K, Lin J, Wu J, Mu J. Spatio-Temporal Variations of Zooplankton and Correlations with Environmental Parameters around Tiaowei Island, Fujian, China. International Journal of Environmental Research and Public Health. 2022; 19(19):12731. https://doi.org/10.3390/ijerph191912731
Chicago/Turabian StyleZhang, Zhi, Zhizhou Shi, Zefeng Yu, Konglin Zhou, Jing Lin, Jiangyue Wu, and Jingli Mu. 2022. "Spatio-Temporal Variations of Zooplankton and Correlations with Environmental Parameters around Tiaowei Island, Fujian, China" International Journal of Environmental Research and Public Health 19, no. 19: 12731. https://doi.org/10.3390/ijerph191912731