

A Preliminary Study of Bacterioplankton Community Structure in the Taiyangshan Wetland in Ningxia and Its Driving Factors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Area and Distribution of Sampling Sites
2.2. Sample Collection and Measurement of Physical and Chemical Indicators
2.3. Extraction, High-Throughput Sequencing, and Bioinformatics Analysis of DNA
2.4. Statistical Analysis
3. Results
3.1. Characteristics of Water Environment in the Taiyangshan Wetland
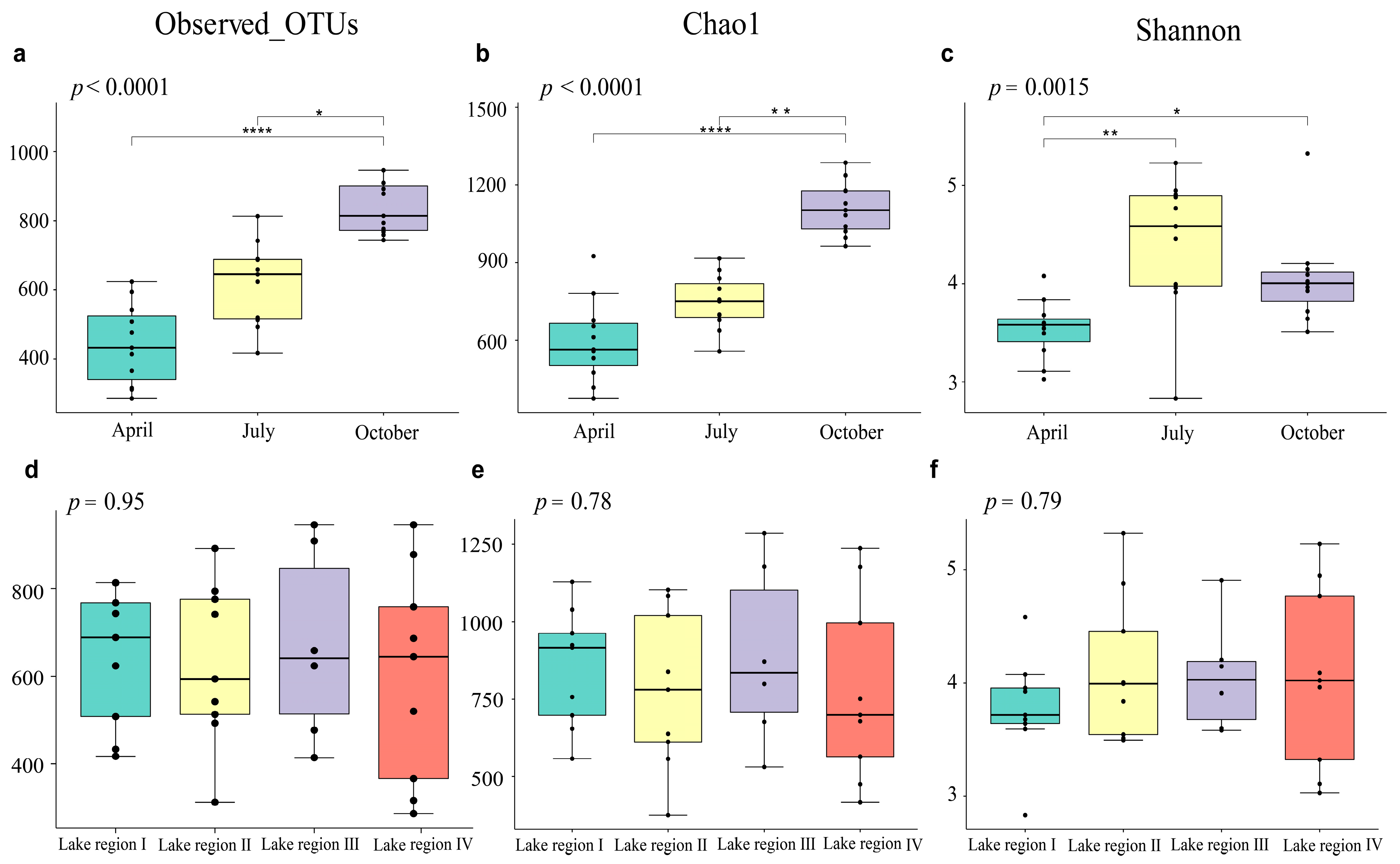
3.2. Variations in Bacterioplankton Diversity in Different Sampling Periods and Lake Regions
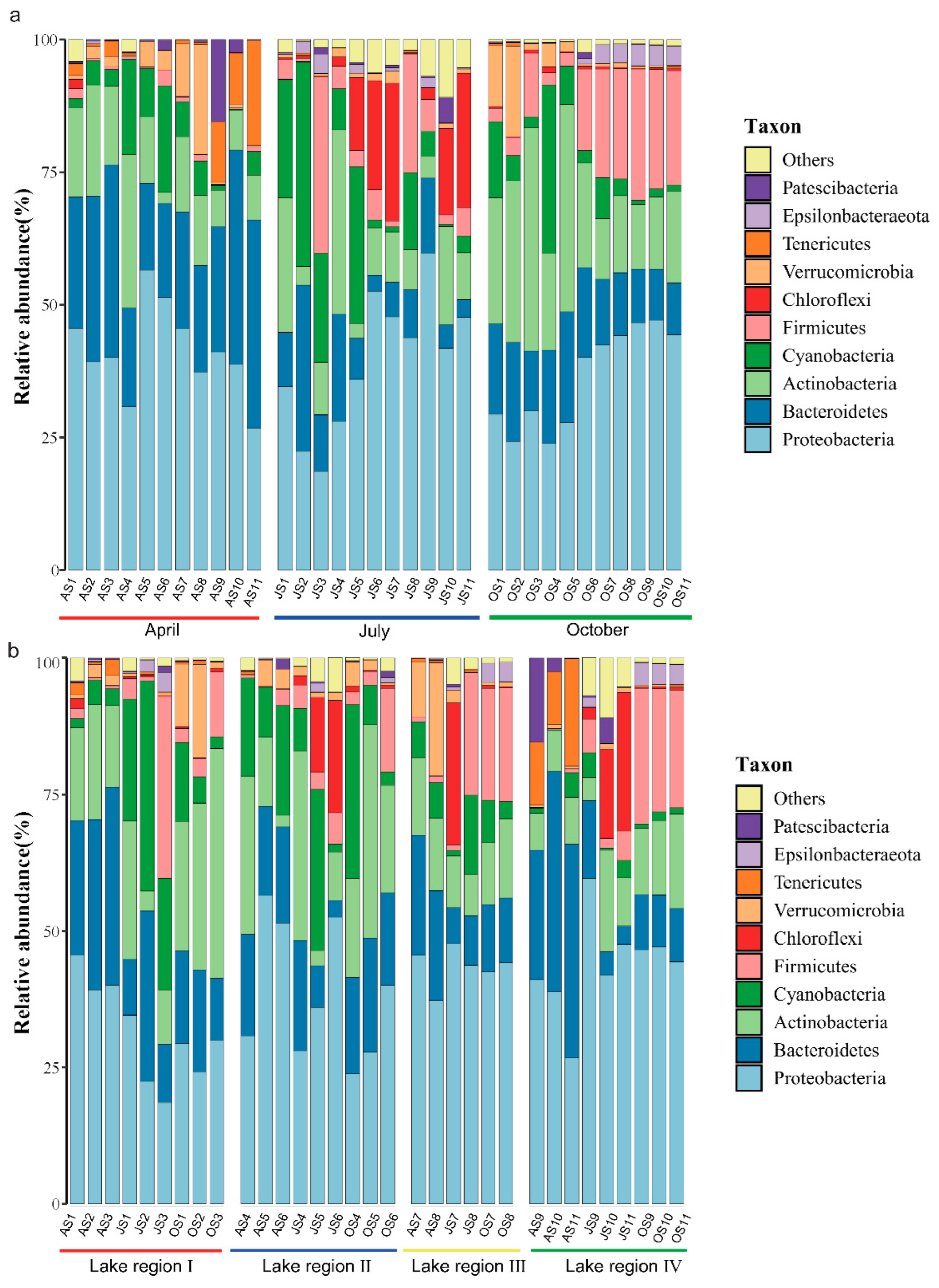
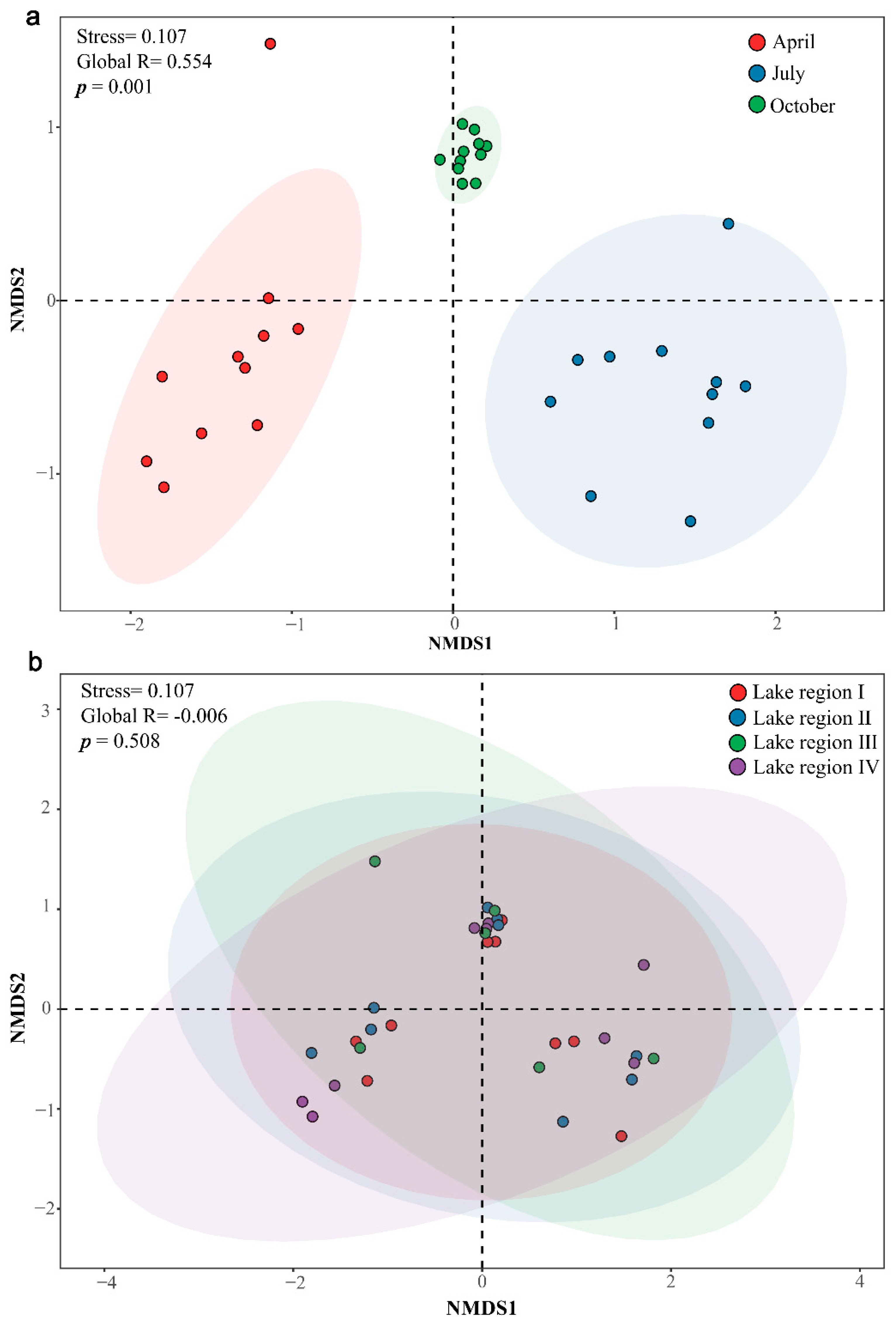
3.3. Community Structures of Bacterioplankton in Different Sampling Periods and Lake Regions
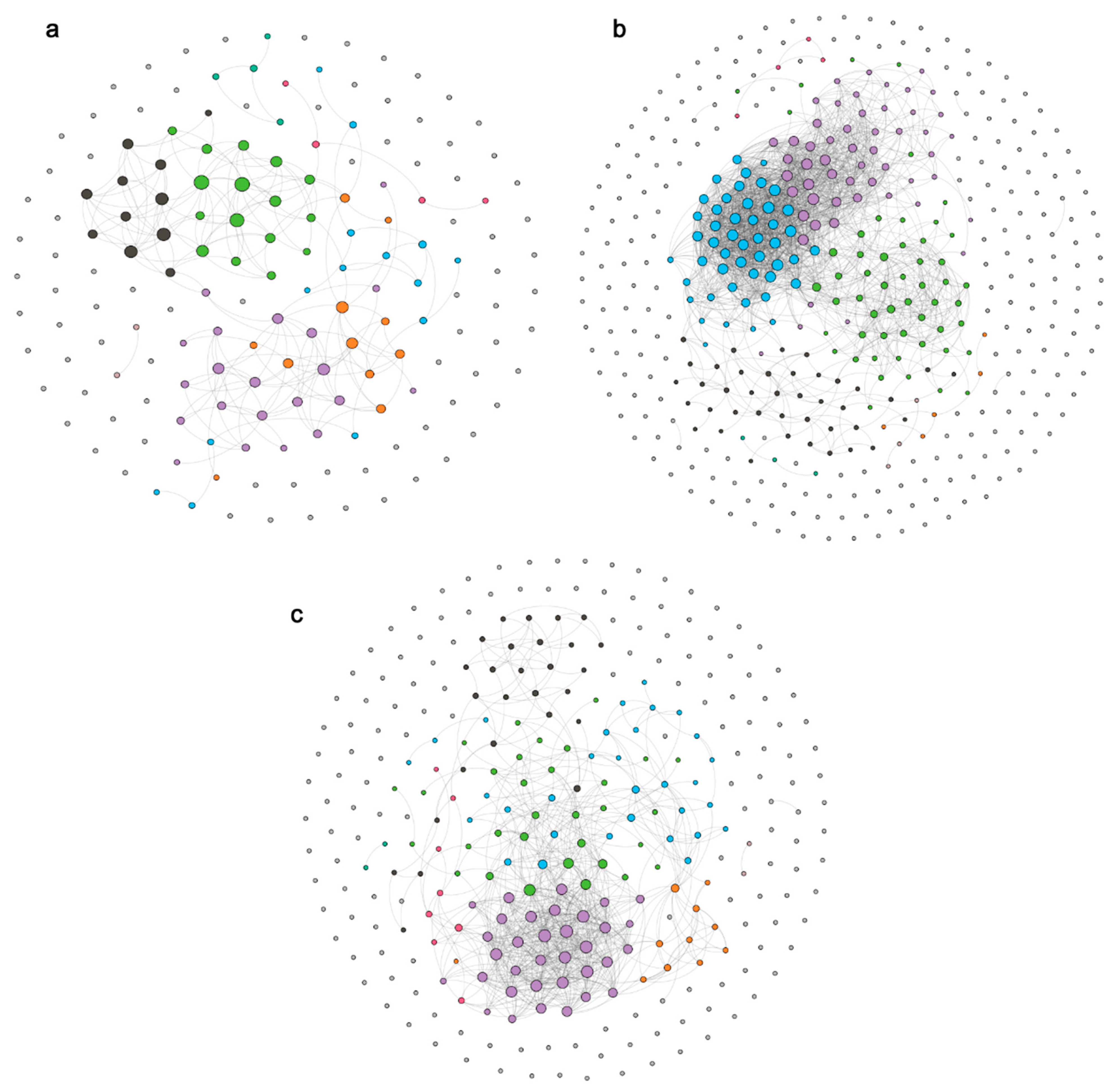
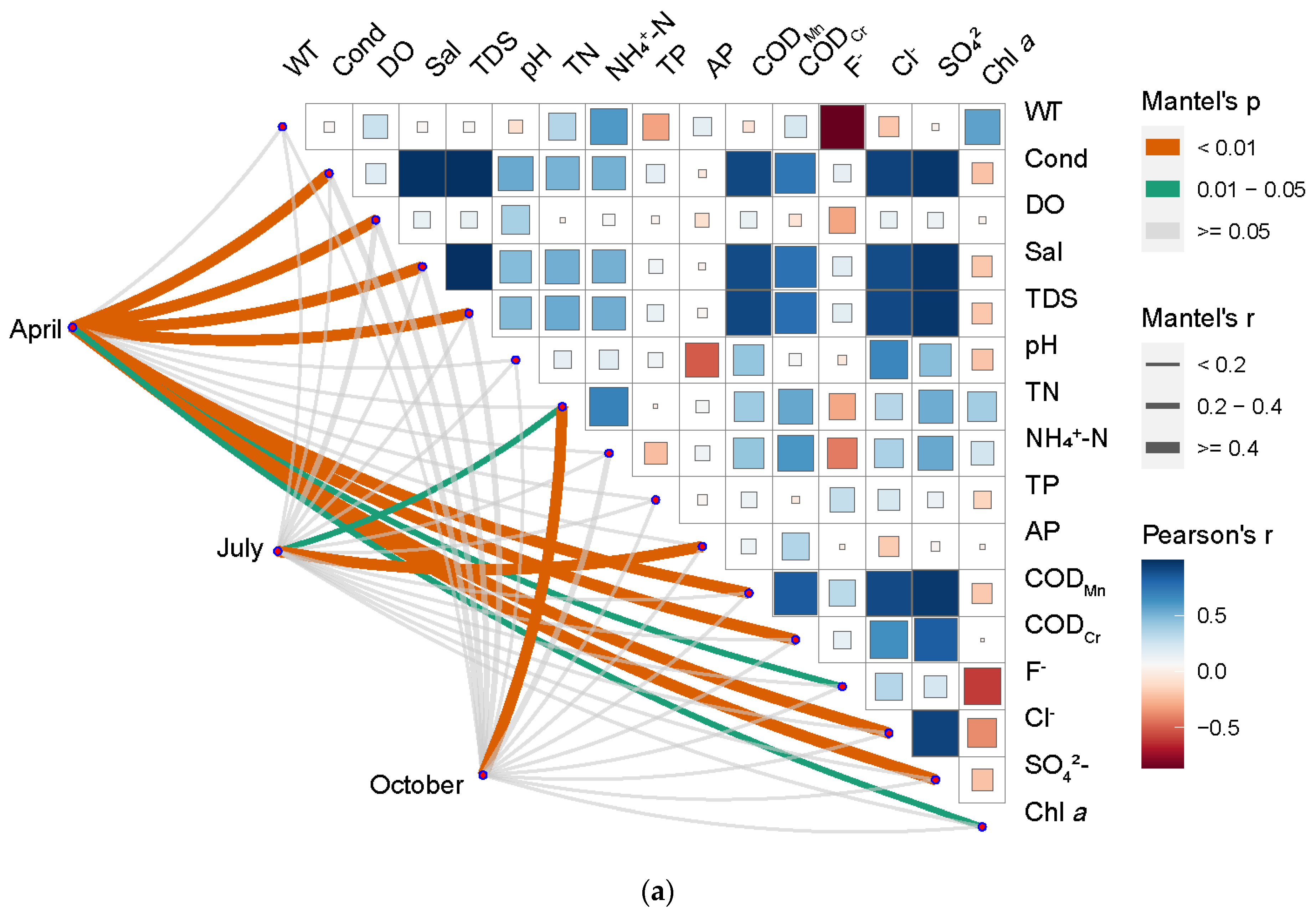
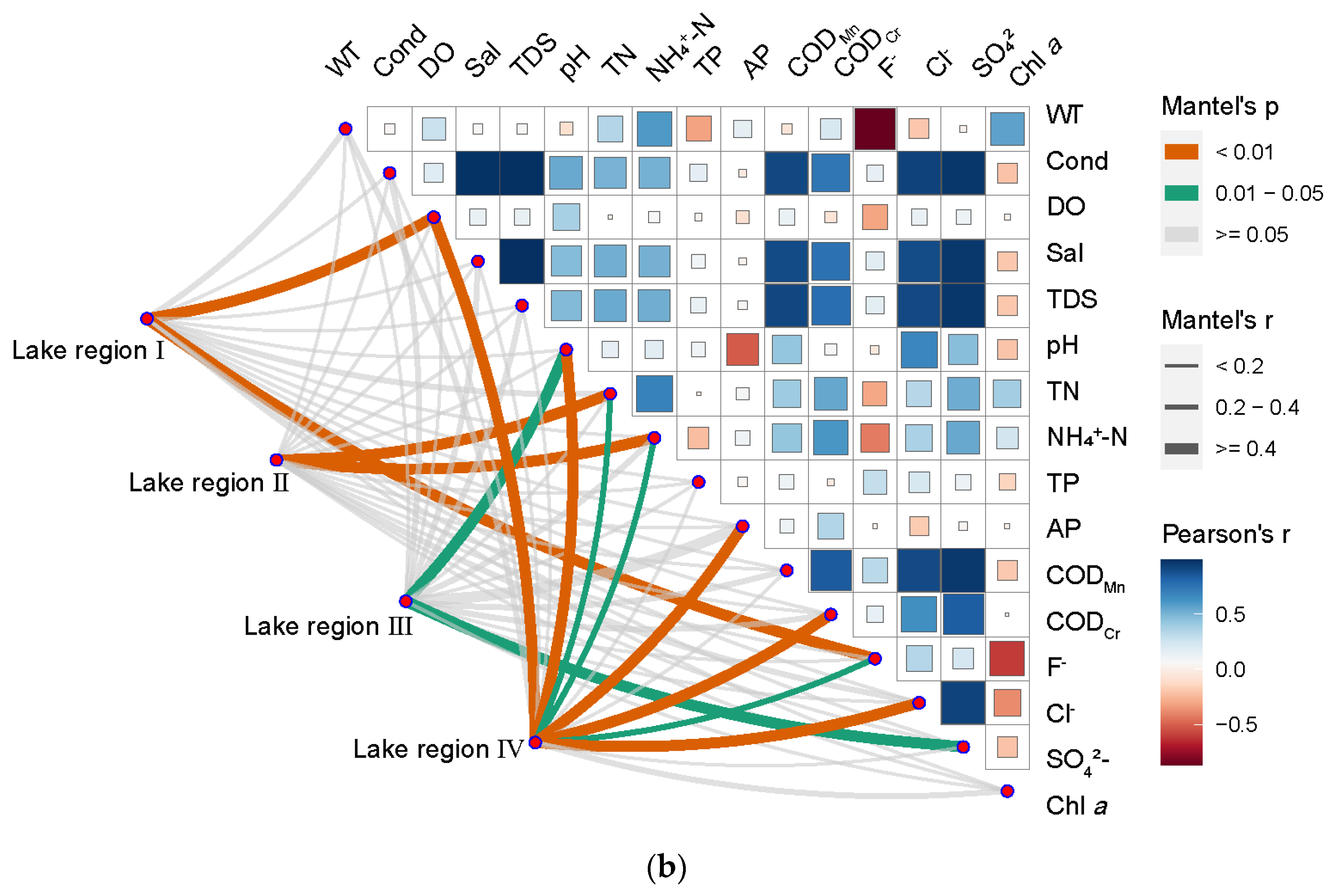
3.4. Correlation Analysis between Bacterioplankton Community Structure and Water Environment Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, R.C.; Finlayson, C. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People 2018. Available online: https://researchoutput.csu.edu.au/en/publications/global-wetland-outlook-state-of-the-worlds-wetlands-and-their-ser (accessed on 20 December 2021).
- Zedler, J.B.; Kercher, S. WETLAND RESOURCES: Status, Trends, Ecosystem Services, and Restorability. Annu. Rev. Environ. Resour. 2005, 30, 39–74. [Google Scholar] [CrossRef]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Findlay, S. Stream microbial ecology. J. N. Am. Benthol. Soc. 2010, 29, 170–181. [Google Scholar] [CrossRef]
- Madsen, E.L. Microorganisms and their roles in fundamental biogeochemical cycles. Curr. Opin. Biotechnol. 2011, 22, 456–464. [Google Scholar] [CrossRef]
- Sims, A.; Zhang, Y.; Gajaraj, S.; Brown, P.B.; Hu, Z. Toward the development of microbial indicators for wetland assessment. Water Res. 2013, 47, 1711–1725. [Google Scholar] [CrossRef]
- Pinto, I.; Calisto, R.; Serra, C.R.; Lage, O.M.; Antunes, S.C. Bacterioplankton Community as a Biological Element for Reservoirs Water Quality Assessment. Water 2021, 13, 2836. [Google Scholar] [CrossRef]
- Escalas, A.; Hale, L.; Voordeckers, J.W.; Yang, Y.; Firestone, M.K.; Alvarez-Cohen, L.; Zhou, J. Microbial functional diversity: From concepts to applications. Ecol. Evol. 2019, 9, 12000–12016. [Google Scholar] [CrossRef]
- Rappé, M.S.; Connon, S.A.; Vergin, K.; Giovannoni, S.J. Cultivation of the ubiquitous SAR11 marine bacterioplankton clade. Nature 2002, 418, 630–633. [Google Scholar] [CrossRef]
- Gich, F.; Schubert, K.; Bruns, A.; Hoffelner, H.; Overmann, J. Specific Detection, Isolation, and Characterization of Selected, Previously Uncultured Members of the Freshwater Bacterioplankton Community. Appl. Environ. Microbiol. 2005, 71, 5908–5919. [Google Scholar] [CrossRef]
- Raes, J.; Bork, P. Molecular eco-systems biology: Towards an understanding of community function. Nat. Rev. Genet. 2008, 6, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Staley, C.; Unno, T.; Gould, T.J.; Jarvis, B.; Phillips, J.; Cotner, J.B.; Sadowsky, M.J. Application of Illumina next-generation sequencing to characterize the bacterial community of the Upper Mississippi River. J. Appl. Microbiol. 2013, 115, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Eiler, A.; Heinrich, F.; Bertilsson, S. Coherent dynamics and association networks among lake bacterioplankton taxa. ISME J. 2011, 6, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.; Liu, Y.; Xie, S.; Liu, Y. Bacterioplankton communities in a high-altitude freshwater wetland. Ann. Microbiol. 2013, 64, 1405–1411. [Google Scholar] [CrossRef]
- Adamovich, B.; Medvinsky, A.; Nikitina, L.; Radchikova, N.; Mikheyeva, T.; Kovalevskaya, R.; Veres, Y.; Chakraborty, A.; Rusakov, A.; Nurieva, N.; et al. Relations between variations in the lake bacterioplankton abundance and the lake trophic state: Evidence from the 20-year monitoring. Ecol. Indic. 2018, 97, 120–129. [Google Scholar] [CrossRef]
- Huws, S.A.; Edwards, J.E.; Kim, E.J.; Scollan, N.D. Specificity and sensitivity of eubacterial primers utilized for molecular profiling of bacteria within complex microbial ecosystems. J. Microbiol. Methods 2007, 70, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Barbier, B.A.; Bottos, E.M.; McDonald, I.; Cary, S. The Inter-Valley Soil Comparative Survey: The ecology of Dry Valley edaphic microbial communities. ISME J. 2011, 6, 1046–1057. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Yang, N.; Wen, Z.; Sun, X.; Chai, Y.; Ma, Z. Wheat microbiome bacteria can reduce virulence of a plant pathogenic fungus by altering histone acetylation. Nat. Commun. 2018, 9, 3429. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, J.; Liu, J.; Du, D. Responses of soil N-fixing bacteria communities to invasive species over a gradient of simulated nitrogen deposition. Ecol. Eng. 2017, 98, 32–39. [Google Scholar] [CrossRef]
- Cui, Y.; Bing, H.; Fang, L.; Wu, Y.; Yu, J.; Shen, G.; Jiang, M.; Wang, X.; Zhang, X. Diversity patterns of the rhizosphere and bulk soil microbial communities along an altitudinal gradient in an alpine ecosystem of the eastern Tibetan Plateau. Geoderma 2018, 338, 118–127. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. 2021. R Package Version 0.7.0. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 5 December 2021).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 14 December 2020).
- Harell, F.E., Jr. Hmisc: Harrell Miscellaneous. 2021. R Package Version 4.6-0. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 5 December 2021).
- Zhang, T.; Xu, S.; Yan, R.; Wang, R.; Gao, Y.; Kong, M.; Yi, Q.; Zhang, Y. Similar geographic patterns but distinct assembly processes of abundant and rare bacterioplankton communities in river networks of the Taihu Basin. Water Res. 2022, 211, 118057. [Google Scholar] [CrossRef]
- Huang, H. linkET: Everything is Linkable. R Package Version 0.0.4. 2021. Available online: https://github.com/Hy4m/linkET (accessed on 20 October 2021).
- Cheng, X.; Xing, W.; Liu, J. Forest Litter Chemical Traits and Microbial Stoichiometry Regulate the Formation and Stabilization of Soil Organic Matter. SSRN Electron. J. 2022. [Google Scholar] [CrossRef]
- Tilman, D.; Kilham, S.S.; Kilham, P. Phytoplankton Community Ecology: The Role of Limiting Nutrients. Annu. Rev. Ecol. Syst. 1982, 13, 349–372. [Google Scholar] [CrossRef]
- Thomas, M.K.; Aranguren-Gassis, M.; Kremer, C.T.; Gould, M.R.; Anderson, K.; Klausmeier, C.A.; Litchman, E. Temperature–nutrient interactions exacerbate sensitivity to warming in phytoplankton. Glob. Chang. Biol. 2017, 23, 3269–3280. [Google Scholar] [CrossRef] [PubMed]
- Lindström, E.S. Bacterioplankton Community Composition in Five Lakes Differing in Trophic Status and Humic Content. Microb. Ecol. 2000, 40, 104–113. [Google Scholar] [CrossRef]
- Haukka, K.; Kolmonen, E.; Hyder, R.; Hietala, J.; Vakkilainen, K.; Kairesalo, T.; Haario, H.; Sivonen, K. Effect of Nutrient Loading on Bacterioplankton Community Composition in Lake Mesocosms. Microb. Ecol. 2006, 51, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, T.; Wang, Q.; Li, L.; Shen, T.; Gao, G. Bacterial community composition in aquatic and sediment samples with spatiotemporal dynamics in large, shallow, eutrophic Lake Chaohu, China. J. Freshw. Ecol. 2019, 34, 575–589. [Google Scholar] [CrossRef]
- Mary, I.; Cummings, D.; Biegala, I.; Burkill, P.; Archer, S.; Zubkov, M. Seasonal dynamics of bacterioplankton community structure at a coastal station in the western English Channel. Aquat. Microb. Ecol. 2006, 42, 119–126. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, J.; Nawaz, M.Z.; Mahboob, S.; Al-Ghanim, K.A.; Khan, I.A.; Lu, Z.; Chen, T. Seasonal succession and spatial distribution of bacterial community structure in a eutrophic freshwater Lake, Lake Taihu. Sci. Total Environ. 2019, 669, 29–40. [Google Scholar] [CrossRef]
- Hu, H.; He, J.; Yan, H.; Hou, D.; Zhang, D.; Liu, L.; Wang, K. Seasonality in Spatial Turnover of Bacterioplankton Along an Ecological Gradient in the East China Sea: Biogeographic Patterns, Processes and Drivers. Microorganisms 2020, 8, 1484. [Google Scholar] [CrossRef]
- Kosek, K.; Luczkiewicz, A.; Kozioł, K.; Jankowska, K.; Ruman, M.; Polkowska, Ż. Environmental characteristics of a tundra river system in Svalbard. Part 1: Bacterial abundance, community structure and nutrient levels. Sci. Total Environ. 2018, 653, 1571–1584. [Google Scholar] [CrossRef]
- Salazar, R.N.; Aguirre, C.; Soto, J.; Salinas, P.; Salinas, C.; Prieto, H.; Paneque, M. Physicochemical Parameters Affecting the Distribution and Diversity of the Water Column Microbial Community in the High-Altitude Andean Lake System of La Brava and La Punta. Microorganisms 2020, 8, 1181. [Google Scholar] [CrossRef]
- Hamilton, W.; Lenton, T. Spora and Gaia: How microbes fly with their clouds. Ethol. Ecol. Evol. 1998, 10, 1–16. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J.; Michot, T.C. Invertebrate Eggs Can Fly: Evidence of Waterfowl-Mediated Gene Flow in Aquatic Invertebrates. Am. Nat. 2005, 165, 274–280. [Google Scholar] [CrossRef]
- Tang, X.; Xie, G.; Shao, K.; Hu, Y.; Cai, J.; Bai, C.; Gong, Y.; Gao, G. Contrast diversity patterns and processes of microbial community assembly in a river-lake continuum across a catchment scale in northwestern China. Environ. Microbiome 2020, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, L.; Fu, R.; Cheng, L.; Yan, S.; Mahtab, N.; Song, Y. Effects of foraging site distances on the intestinal bacterial community compositions of the sympatric wintering Hooded Crane (Grus monacha) and Domestic Duck (Anas platyrhynchos domesticus). Avian Res. 2021, 12, 20. [Google Scholar] [CrossRef]
- Shang, Y.; Wu, X.; Wang, X.; Wei, Q.; Ma, S.; Sun, G.; Zhang, H.; Wang, L.; Dou, H.; Zhang, H. Factors affecting seasonal variation of microbial community structure in Hulun Lake, China. Sci. Total Environ. 2021, 805, 150294. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Genet. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef]
- Czárán, T.L.; Hoekstra, R.F.; Pagie, L. Chemical warfare between microbes promotes biodiversity. Proc. Natl. Acad. Sci. USA 2002, 99, 786–790. [Google Scholar] [CrossRef]
- Logue, J.B.; Lindström, E.S. Biogeography of Bacterioplankton in Inland Waters. Freshw. Rev. 2008, 1, 99–114. [Google Scholar] [CrossRef]
- Lindström, E.S.; Forslund, M.; Algesten, G.; Bergström, A.-K. External control of bacterial community structure in lakes. Limnol. Oceanogr. 2006, 51, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Schiaffino, M.R.; Unrein, F.; Gasol, J.M.; Massana, R.; Balagué, V.; Izaguirre, I. Bacterial community structure in a latitudinal gradient of lakes: The roles of spatial versus environmental factors: Factors shaping bacterioplankton community structure. Freshw. Biol. 2011, 56, 1973–1991. [Google Scholar] [CrossRef]
- Campbell, B.J.; Kirchman, D.L. Bacterial diversity, community structure and potential growth rates along an estuarine salinity gradient. ISME J. 2012, 7, 210–220. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Knight, R. Global patterns in bacterial diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Ocean Plankton. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef]
- Spietz, R.; Williams, C.M.; Rocap, G.; Horner-Devine, C. A Dissolved Oxygen Threshold for Shifts in Bacterial Community Structure in a Seasonally Hypoxic Estuary. PLoS ONE 2015, 10, e0135731. [Google Scholar] [CrossRef]
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef]
- Newton, R.J.; McMahon, K.D. Seasonal differences in bacterial community composition following nutrient additions in a eutrophic lake: Seasonal variation in nutrient-amended microcosms. Environ. Microbiol. 2010, 13, 887–899. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, Z.; Pei, Y. Evaluation of water quality and nitrogen removal bacteria community in Fuhe River. Procedia Environ. Sci. 2012, 13, 1809–1819. [Google Scholar] [CrossRef]
- Song, S.; Yang, F.; Ma, L.-A.; Gao, X.-F. Effects of Sewage Treatment on Microbial Community Structure of Surface Water in Xiantao Wetland. OALib 2018, 5, 1–12. [Google Scholar] [CrossRef]
- Yannarell, A.C.; Triplett, E.W. Geographic and Environmental Sources of Variation in Lake Bacterial Community Composition. Appl. Environ. Microbiol. 2005, 71, 227–239. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Parameters | April | July | October | Statistical Significance |
|---|---|---|---|---|
| WT (°C) | 19.78 ± 1.54 | 25.25 ± 2.63 | 11.15 ± 1.49 | * |
| Cond (μS/cm) | 15,372 ± 12,827 | 18,946 ± 16,102 | 17,406 ± 15,074 | ns |
| DO (mg/L) | 5.34 ± 1.11 | 6.51 ± 1.63 | 5.52 ± 0.97 | ns |
| Sal (ppt) | 10.93 ± 10.5 | 11.67 ± 10.36 | 11.13 ± 10.77 | ns |
| TDS (mg/L) | 11,584 ± 10,165 | 12,185 ± 10,320 | 11,404 ± 9946 | ns |
| pH | 7.68 ± 0.17 | 8.65 ± 0.70 | 8.56 ± 0.57 | * |
| TN (mg/L) | 2.94 ± 1.89 | 2.77 ± 1.82 | 1.65 ± 1.66 | ns |
| NH4+-N (mg/L) | 1.34 ± 0.91 | 1.68 ± 1.59 | 0.25 ± 0.1 | * |
| TP (mg/L) | 0.04 ± 0.02 | 0.04 ± 0.01 | 0.08 ± 0.07 | ns |
| AP (mg/L) | 0.019 ± 0.012 | 0.003 ± 0.002 | 0.002 ± 0.001 | * |
| CODMn (mg/L) | 6.58 ± 3.87 | 5.11 ± 3.03 | 6.25 ± 4.33 | ns |
| CODCr (mg/L) | 160.35 ± 208.24 | 35.48 ± 27.38 | 28.04 ± 20.3 | * |
| F− (mg/L) | 3.99 ± 0.39 | 2.98 ± 0.27 | 4.95 ± 0.4 | * |
| Cl− (mg/L) | 8388 ± 8144 | 12,236 ± 12,783 | 16,707 ± 15,374 | ns |
| SO42− (mg/L) | 6135 ± 5695 | 5624.14 ± 5239 | 5835 ± 5626 | ns |
| Chl a (mg/L) | 27.46 ± 17.64 | 25.1 ± 10.19 | 11.19 ± 8.04 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.-Z.; Zhang, W.-J.; Zhang, W.; Zhao, Z.-F.; Qiu, X.-C. A Preliminary Study of Bacterioplankton Community Structure in the Taiyangshan Wetland in Ningxia and Its Driving Factors. Int. J. Environ. Res. Public Health 2022, 19, 12224. https://doi.org/10.3390/ijerph191912224
Zhao R-Z, Zhang W-J, Zhang W, Zhao Z-F, Qiu X-C. A Preliminary Study of Bacterioplankton Community Structure in the Taiyangshan Wetland in Ningxia and Its Driving Factors. International Journal of Environmental Research and Public Health. 2022; 19(19):12224. https://doi.org/10.3390/ijerph191912224
Chicago/Turabian StyleZhao, Rui-Zhi, Wei-Jiang Zhang, Wen Zhang, Zeng-Feng Zhao, and Xiao-Cong Qiu. 2022. "A Preliminary Study of Bacterioplankton Community Structure in the Taiyangshan Wetland in Ningxia and Its Driving Factors" International Journal of Environmental Research and Public Health 19, no. 19: 12224. https://doi.org/10.3390/ijerph191912224