

DNA Barcoding of Chironomid Larvae (Diptera: Chironomidae) from Large Rivers in South Korea to Facilitate Freshwater Biomonitoring and Public Health Surveillance

Abstract
:1. Introduction
2. Materials and Methods
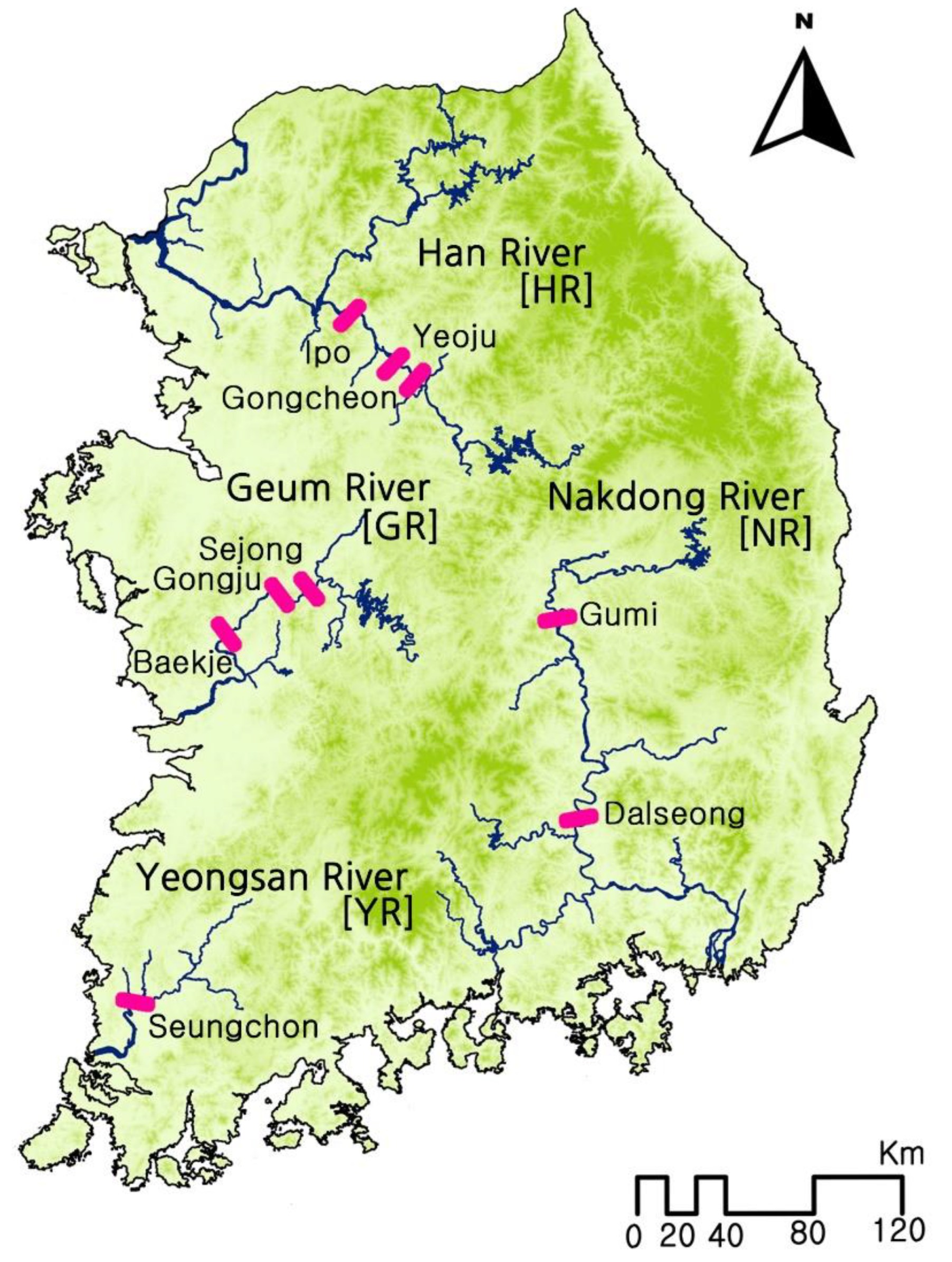
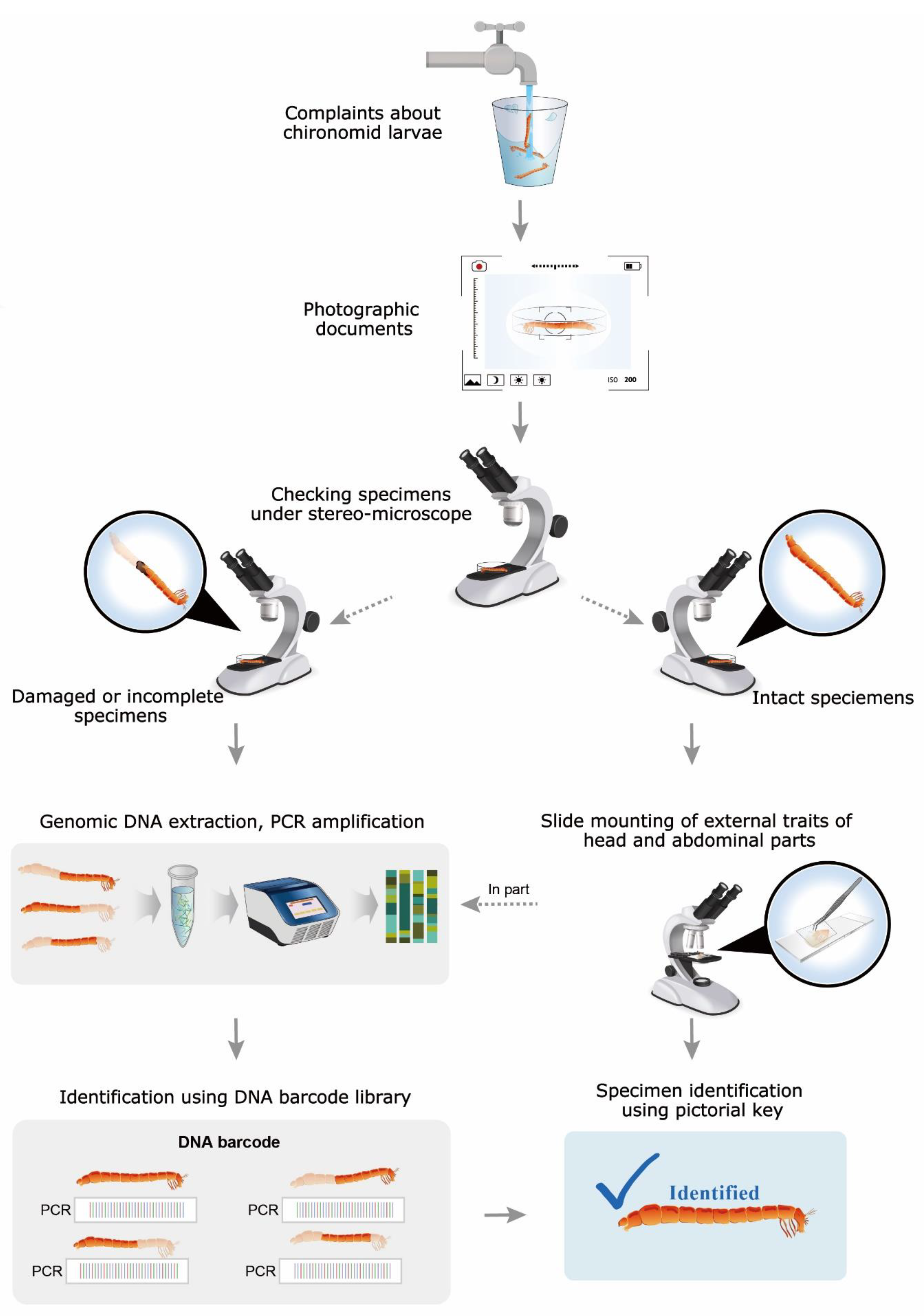
2.1. Larval Sampling and Morphological Identification
2.2. Molecular Analysis
3. Results
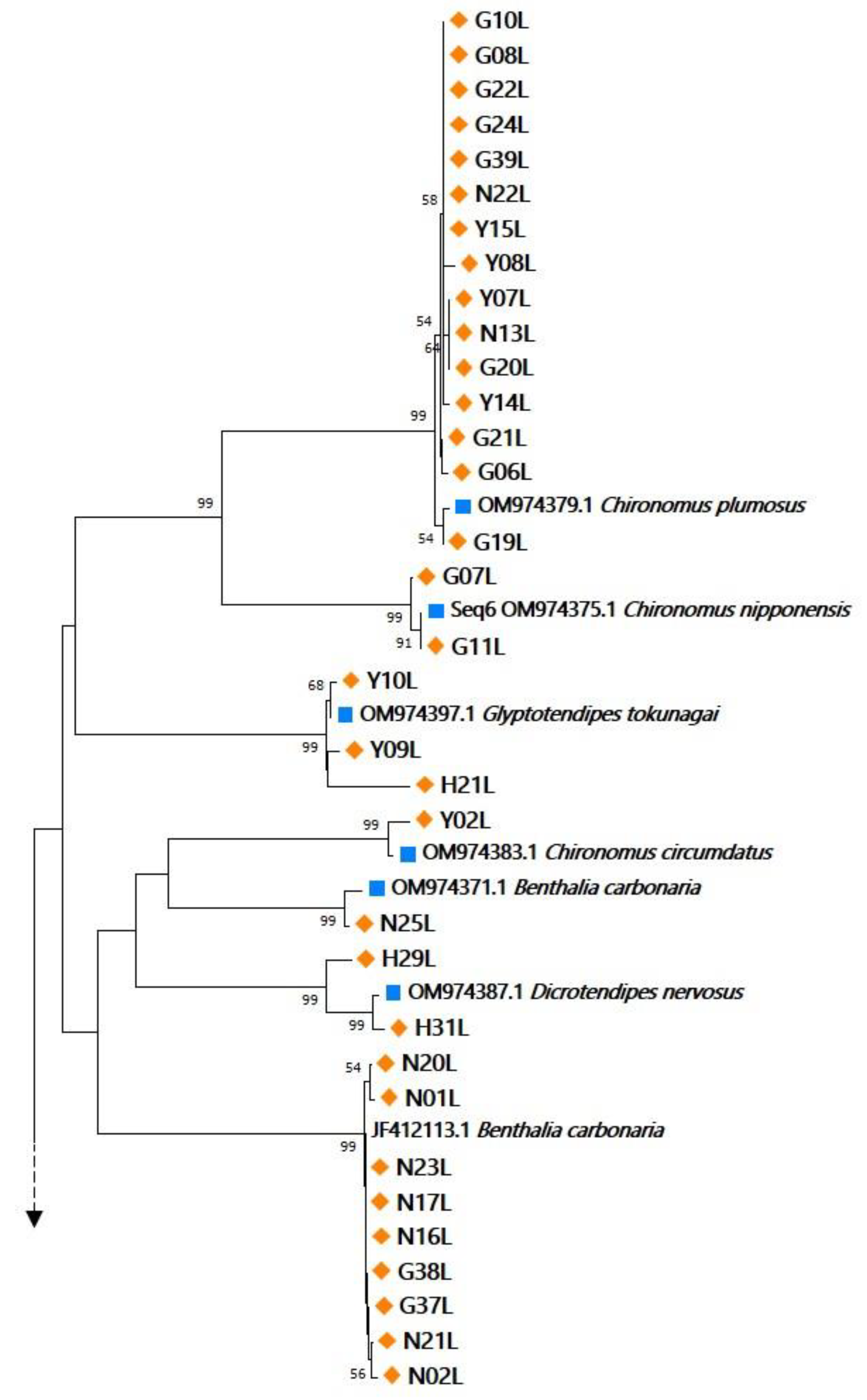
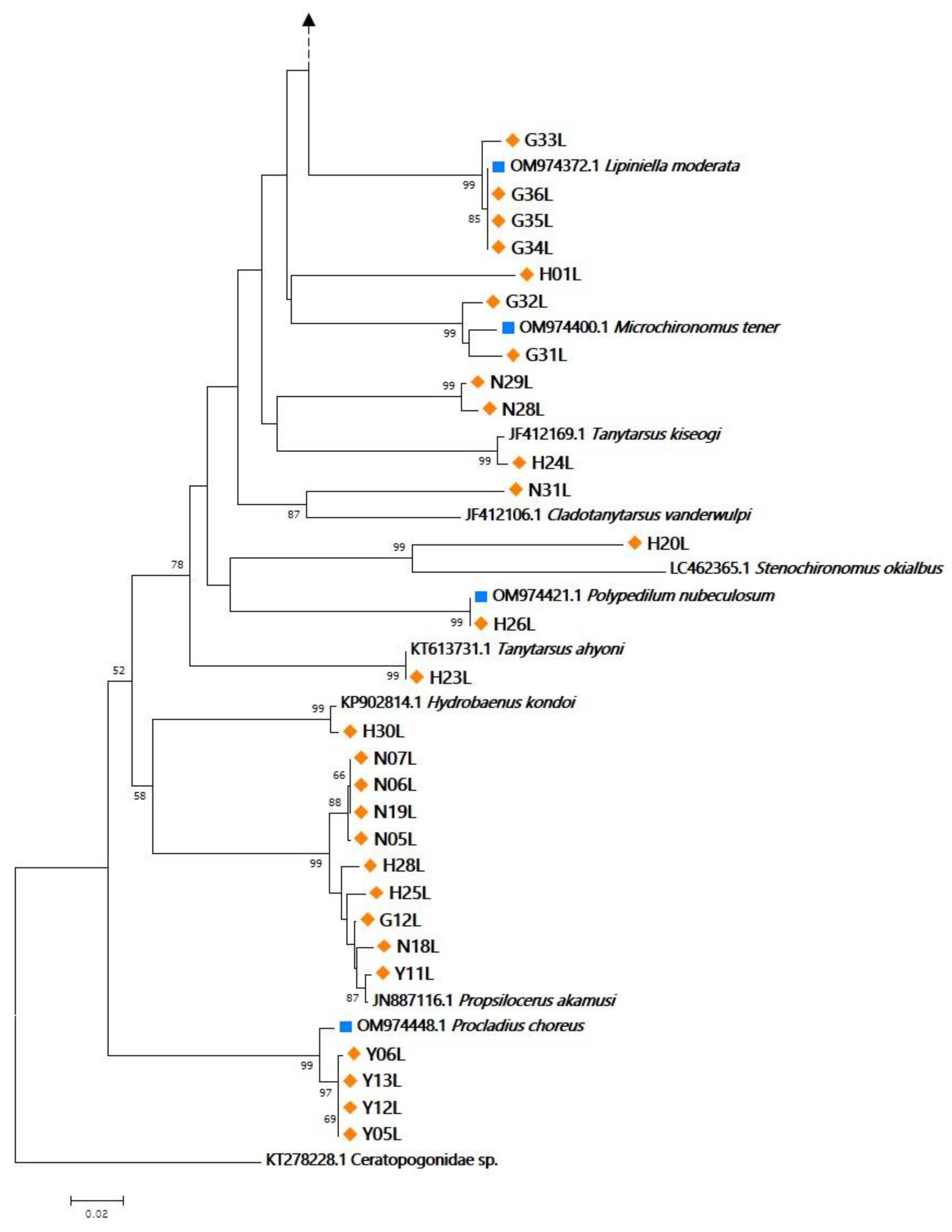
3.1. Specimen Identification
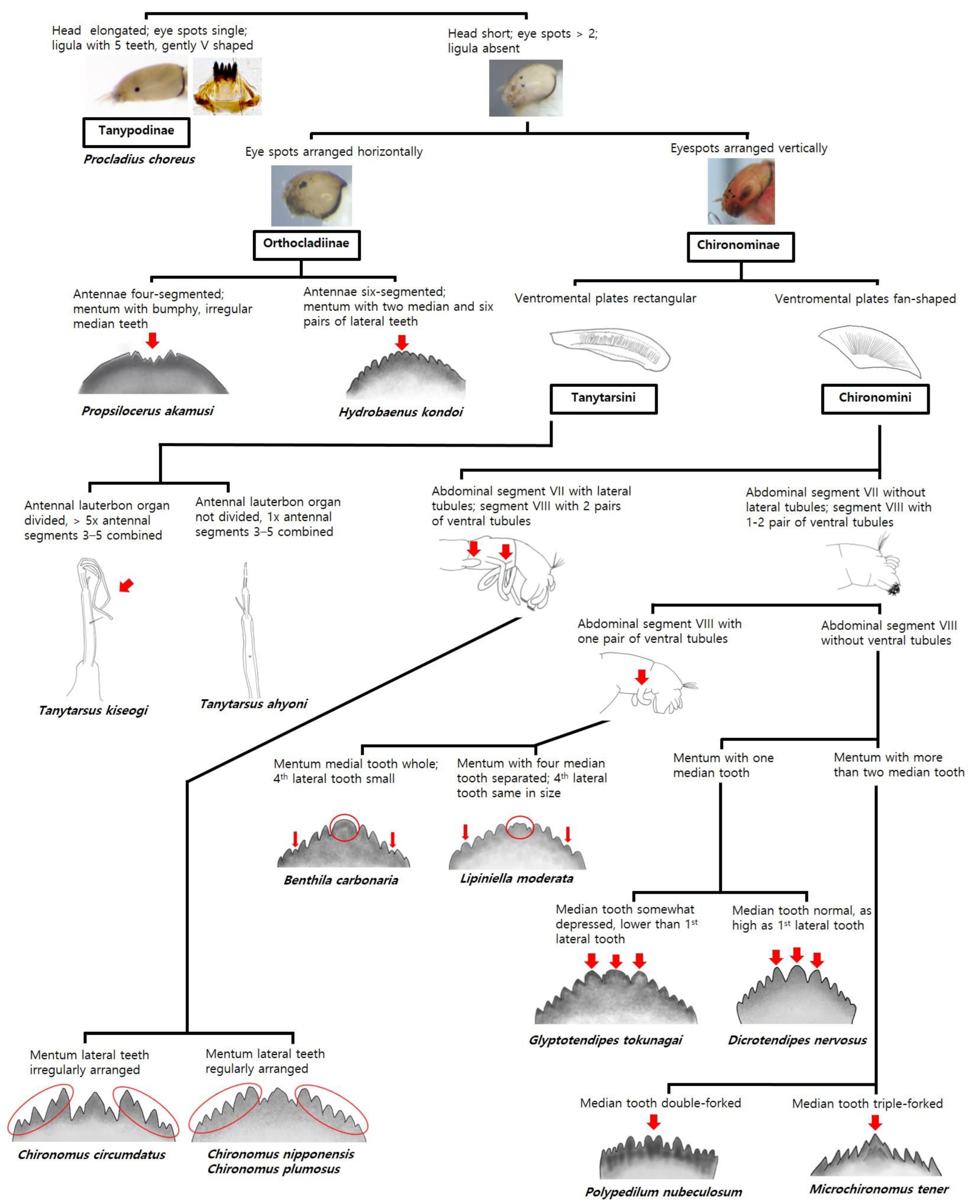
3.2. Morphological Key and Action Protocol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, H.; Choo, G.; Kim, H.; Oh, J.E. Evaluation of the current spill status of PFASs and OPFRs in South Korean tap water associated with its origin. Sci. Total Environ. 2018, 634, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.J.; Lee, B.H. Human impacts on stream ecosystems and freshwater arthropods in Korea. Korean J. Entomol. 2001, 31, 63–76. [Google Scholar]
- K-Water. Research of Benthic Macroinvertebrates in 4 Major Rivers and Major Inflow Rivers; K-Water: Deajeon, Korea, 2016. [Google Scholar]
- Kwak, I.; Lee, D.; Hong, C.; Park, Y. Distribution patterns of benthic macroinvertebrates in streams of Korea. Korean J. Ecol. Environ. 2018, 51, 60–70. [Google Scholar] [CrossRef]
- Min, J.; Kong, D. Distribution patterns of benthic macroinvertebrate communities based on multispatial-scale environmental variables in the river systems of Republic of Korea. J. Freshw. Ecol. 2020, 35, 323–347. [Google Scholar] [CrossRef]
- Kwak, I.; Park, J.; Kim, W.; Park, K. Morphological and genetic species identification in the Chironomus larvae (Diptera: Chironomidae) found in domestic tap water purification plants. Korean J. Ecol. Environ. 2020, 53, 286–294. [Google Scholar] [CrossRef]
- Kwak, I.; Park, J.; Kim, W.; Park, K. Morphological and genetic species identification in the Chironomidae larvae found in tap water purification plants in Jeju. Korean J. Ecol. Environ. 2021, 54, 240–246. [Google Scholar] [CrossRef]
- Rosenberg, D.M.; Resh, V.H. Introduction to freshwater biomonitoring and benthic macroinvertebrates. In Freshwater Biomonitoring and Benthic Macroinvertebrates; Rosenberg, D.M., Resh, V.H., Eds.; Chapman & Hall: New York, NY, USA, 1993; pp. 1–9. [Google Scholar]
- Rosenberg, D.M. Freshwater biomonitoring and Chironomidae. Neth. J. Aquat. Ecol. 1992, 26, 101–122. [Google Scholar] [CrossRef]
- Kawai, K.; Yamagishi, T.; Kubo, Y.; Konishi, K. Usefulness of chironomid larvae as indicators of water quality. Med. Entomol. Zool. 1989, 40, 269–283. [Google Scholar] [CrossRef]
- Vermeulen, A.C. Elaborating chironomid deformities as bioindicators of toxic sediment stress: The potential application of mixture toxicity concepts. Ann. Zool. Fenn. 1995, 32, 265–285. [Google Scholar]
- Baek, M.J.; Kang, H.J.; Yoon, T.J.; Bae, Y.J. Biological and genetic characteristics of Glyptotendipes tokunagai (Diptera: Chironomidae) on the basis of successive rearing of forty-two generations over seven years under laboratory conditions. Environ. Entomol. 2014, 43, 1406–1418. [Google Scholar] [CrossRef]
- Lin, C.T.; Chiu, M.C.; Kuo, M.H. Effects of anthropogenic activities on microplastics in deposit-feeders (Diptera: Chironomidae) in an urban river of Taiwan. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Ekrem, T.; Stur, E.; Hebert, P.D.N. Females do count: Documenting Chironomidae (Diptera) species diversity using DNA barcoding. Org. Divers. Evol. 2010, 10, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Maier, K.J.; Kosalwat, P.; Knight, A.W. Culture of Chironomus decorus (Diptera: Chironomidae) and the effect of temperature on its life history. Environ. Entomol. 1990, 19, 1681–1688. [Google Scholar] [CrossRef]
- Ristola, T.; Pellinen, J.; Ruokolainen, M.; Kostamo, A.; Kukkonen, J.V.K. Effect of sediment type, feeding level, and larval density on growth and development of a midge (Chironomus riparius). Environ. Toxicol. Chem. 1999, 18, 756–764. [Google Scholar] [CrossRef]
- Baek, M.J.; Yoon, Y.J.; Bae, Y.J. Development of Glyptotendipes tokunagai (Diptera: Chronomidae) under different temperature conditions. Environ. Entomol. 2012, 41, 950–958. [Google Scholar] [CrossRef]
- Kang, H.J.; Baek, M.J.; Kang, J.H.; Bae, Y.J. Diversity and DNA barcode analysis of chironomids (Diptera: Chironomidae) from large rivers in South Korea. Insects 2022, 13, 346. [Google Scholar] [CrossRef]
- Wiederholm, T. Chironomidae of the Holarctic Region. Keys and Diagnoses. Part 1: Larvae; Entomologica Scandinavica: Stockholm, Sweden, 1983; pp. 1–457. [Google Scholar]
- Epler, J.H. Identification Manual for the Larval Chironomidae (Diptera) of Florida; Bureau of Surface Water Management, Florida Department of Environmental Protection: Tallahassee, FL, USA, 1995.
- Madden, C.P. Key to genera of larvae of Australian Chironomidae (Diptera). Mus. Vic. Sci. Rep. 2010, 13, 1–31. [Google Scholar] [CrossRef]
- Cranston, P.S. Identification guide to genera of aquatic larval Chironomidae (Diptera) of Australia and New Zealand. Zootaxa 2019, 4706, 71–102. [Google Scholar] [CrossRef]
- Sasa, M.; Kikuchi, M. Chironomidae [Diptera] of Japan, 1st ed.; University of Tokyo Press: Tokyo, Japan, 1995; pp. 1–333. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Kim, S.; Song, K.H.; Ree, H.I.; Kim, W. A DNA barcode library for Korean Chironomidae (Insecta: Diptera) and indexes for defining barcode gap. Mol. Cells 2012, 33, 9–17. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, K.F.; Ball, S.L. DNA barcodes for biosecurity: Invasive species identification. Philosoph. Trans. Roy. Soc. B Biol. Sci. 2005, 360, 1813–1823. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.H.; Xu, M.; Li, C.P. Rapid DNA barcoding analysis of large datasets using the composition vector method. BMC. Bioinform. 2009, 10, S8. [Google Scholar] [CrossRef]
- Ashfaq, M.; Hebert, P.D.N.; Mirza, J.H.; Khan, A.M.; Zafar, Y.; Mirza, M.S. Analyzing mosquito (Diptera: Culicidae) diversity in Pakistan by DNA barcoding. PLoS ONE 2014, 9, e97268. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Suh, K.I.; Bae, Y.J. DNA barcode library and its efficacy for identifying food-associated insect pests in Korea. Entomol. Res. 2013, 43, 253–261. [Google Scholar] [CrossRef]
- Meiklejohn, K.A.; Damaso, N.; Robertson, J.M. Assessment of BOLD and GenBank-Their accuracy and reliability for the identification of biological materials. PLoS ONE 2019, 14, e0217084. [Google Scholar] [CrossRef]
- Meyer, C.P.; Paulay, G. DNA Barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef]
- Kim, W.; Park, J.; Hong, C.; Choi, B.; Kim, H.; Park, Y.; Park, J.; Song, H.; Kwak, I. Changes in community structure of Chironomidae caused by variability of environmental factors among weir sections in Korean rivers. Korean J. Ecol. Environ. 2020, 53, 46–54. [Google Scholar] [CrossRef]
- Nagell, B.; Landahl, C.C. Resistance to anoxia of Chironomus plumosus and Chironomus anthracinus (Diptera) larvae. In Holarctic Ecology; Vik, R., Ed.; Munksgaard: Copenhagen, Danmark, 1978; Volume 1, pp. 333–336. [Google Scholar] [CrossRef]
- Zou, W.; Cai, Y.; Tolonen, K.T.; Zhu, G.; Qin, B.; Peng, K.; Gong, Z. The adaptations to tube-dwelling life of Propsilocerus akamusi (Diptera: Chironomidae) larvae and its eutrophication-tolerant mechanisms. Limnologica 2019, 77, 125684. [Google Scholar] [CrossRef]
- Walshe, B.M. The function of haemoglobin in Chironomus plumosus under natural conditions. J. Exp. Biol. 1950, 27, 73–95. [Google Scholar] [CrossRef]
- Krüger, V.F. Eine parthenogenetische Chironomide als Wasserleitungsschädling. In Die Naturwissenschaften; Süffert, F., Ed.; Springer: Berlin/Heidelberg, Germany, 1941; Volume 36, pp. 556–558. [Google Scholar] [CrossRef]
- Kelly, S.N. Infestation of the Norwich England water system. J. Am. Water Works Assoc. 1955, 47, 330–334. [Google Scholar] [CrossRef]
- Williams, D.A. An infestation by a parthenogenetic chironomid. Water Treat. Exam. 1974, 23, 215–229. [Google Scholar]
- Bay, E.C. Chironomid (Diptera: Chironomidae) larval occurrence and transport in a municipal water system. J. Am. Mosq. Control Assoc. 1993, 9, 275–284. [Google Scholar] [PubMed]
- Alexander, M.K.; Merrit, R.W.; Berg, M.B. New strategic for the control of the parthenogenetic chironomid (Paratanytarsus grimmii) (Diptera: Chironomidae) infesting water systems. J. Am. Mosq. Control Assoc. 1997, 13, 189–192. [Google Scholar] [PubMed]
- Morse, J.; Bae, Y.J.; Sangpradub, N.; Tanida, K.; Vshivkova, T.S.; Yang, L.; Yule, C. Freshwater biomonitoring with benthic macroinvertebrates in East Asia. Front. Ecol. Environ. 2007, 5, 33–42. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Species | Accession Number |
|---|---|---|
| Tanypodinae | Procladius choreus (Meigen, 1804) | OM974448 †, OP381670, OP381671, OP381677, OP381678 |
| Orthocladiinae | Hydrobaenus kondoi Saether, 1989 * | KP902814 †, OP381696 |
| Propsilocerus akamusi (Tokunaga, 1938) | JN887116 †, OP381672, OP381685, OP381686, OP381690, OP381691, OP381692, OP381698, OP381700, OP381718 | |
| Chironominae | Benthalia carbonaria (Meigen, 1804) | OM974371 †, JF412113 †, OP381680, OP381681, OP381682, OP381683, OP381684, OP381687, OP381688, OP381693, OP381694, OP381704, OP381706 |
| Chironomus circumdatus (Kieffer, 1916) | OM974383 †, OP381679 | |
| Chironomus nipponensis Tokunaga, 1940 | OM974375 †, OP381719, OP381722 | |
| Chironomus plumosus (Linnaeus, 1758) | OM974379 †, OP381668, OP381669, OP381675, OP381676, OP381689, OP381705, OP381713, OP381714, OP381715, OP381716, OP381717, OP381720, OP381721, OP381723 | |
| Dicrotendipes nervosus (Staeger, 1839) | OM974387 †, OP381695, OP381697 | |
| Glyptotendipes tokunagai Sasa, 1979 | OM974397 †, OP381673, OP381674, OP381703, | |
| Microchironomus tener (Kieffer, 1918) | OM974400 †, OP381711, OP381712 | |
| Lipiniella moderata Kalugina, 1970 | OM974372 †, OP381707, OP381708, OP381709, OP381710 | |
| Polypedilum nubeculosum (Meigen, 1804) | OM974421 †, OP381699 | |
| Stenochironomus sp. | OP381666 | |
| Tanytarsus ahyoni Ree & Jeong, 2010 | KT613731 †, OP381702 | |
| Tanytarsus kiseogi Ree & Jeong, 2010 | JF412169 †, OP381701 | |
| Chironominae sp. 1 | OP381663 | |
| Chironominae sp. 2 | OP381664, OP381665 | |
| Chironominae sp. 3 | OP381667 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.J.; Baek, M.J.; Kang, J.H.; Bae, Y.J. DNA Barcoding of Chironomid Larvae (Diptera: Chironomidae) from Large Rivers in South Korea to Facilitate Freshwater Biomonitoring and Public Health Surveillance. Int. J. Environ. Res. Public Health 2022, 19, 12035. https://doi.org/10.3390/ijerph191912035
Kang HJ, Baek MJ, Kang JH, Bae YJ. DNA Barcoding of Chironomid Larvae (Diptera: Chironomidae) from Large Rivers in South Korea to Facilitate Freshwater Biomonitoring and Public Health Surveillance. International Journal of Environmental Research and Public Health. 2022; 19(19):12035. https://doi.org/10.3390/ijerph191912035
Chicago/Turabian StyleKang, Hyo Jeong, Min Jeong Baek, Ji Hyoun Kang, and Yeon Jae Bae. 2022. "DNA Barcoding of Chironomid Larvae (Diptera: Chironomidae) from Large Rivers in South Korea to Facilitate Freshwater Biomonitoring and Public Health Surveillance" International Journal of Environmental Research and Public Health 19, no. 19: 12035. https://doi.org/10.3390/ijerph191912035