Hymenolepis diminuta Infection Affects Apoptosis in the Small and Large Intestine

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Choosing a Research Model
- -
- Control group (n = 6)—not infected with H. diminuta tapeworm (0 dpi),
- -
- Group I (n = 6)—8 days after H. diminuta infection (8 dpi),
- -
- Group II (n = 6)—25 days after H. diminuta infection (25 dpi),
- -
- Group III (n = 6)—40 days after H. diminuta infection (40 dpi),
- -
- Group IV (n = 6)—60 days after H. diminuta infection (60 dpi).
2.2. An Assay Using MicroBCA Protein Assay Kit and Plate Reader
2.3. Analysis of Gene Expression in the Small and Large Intestine by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. Analysis of Protein Expression in the Small and Large Intestine by Western Blot- Measurement of Bcl-2, Bax, Cas-3, and Cas-9 Protein Expression
2.5. Immunohistochemical Analysis of Bax, BCl-2, Cas-3, and Cas-9 in the Small and Large Intestine
2.6. Statistical Analysis
3. Results
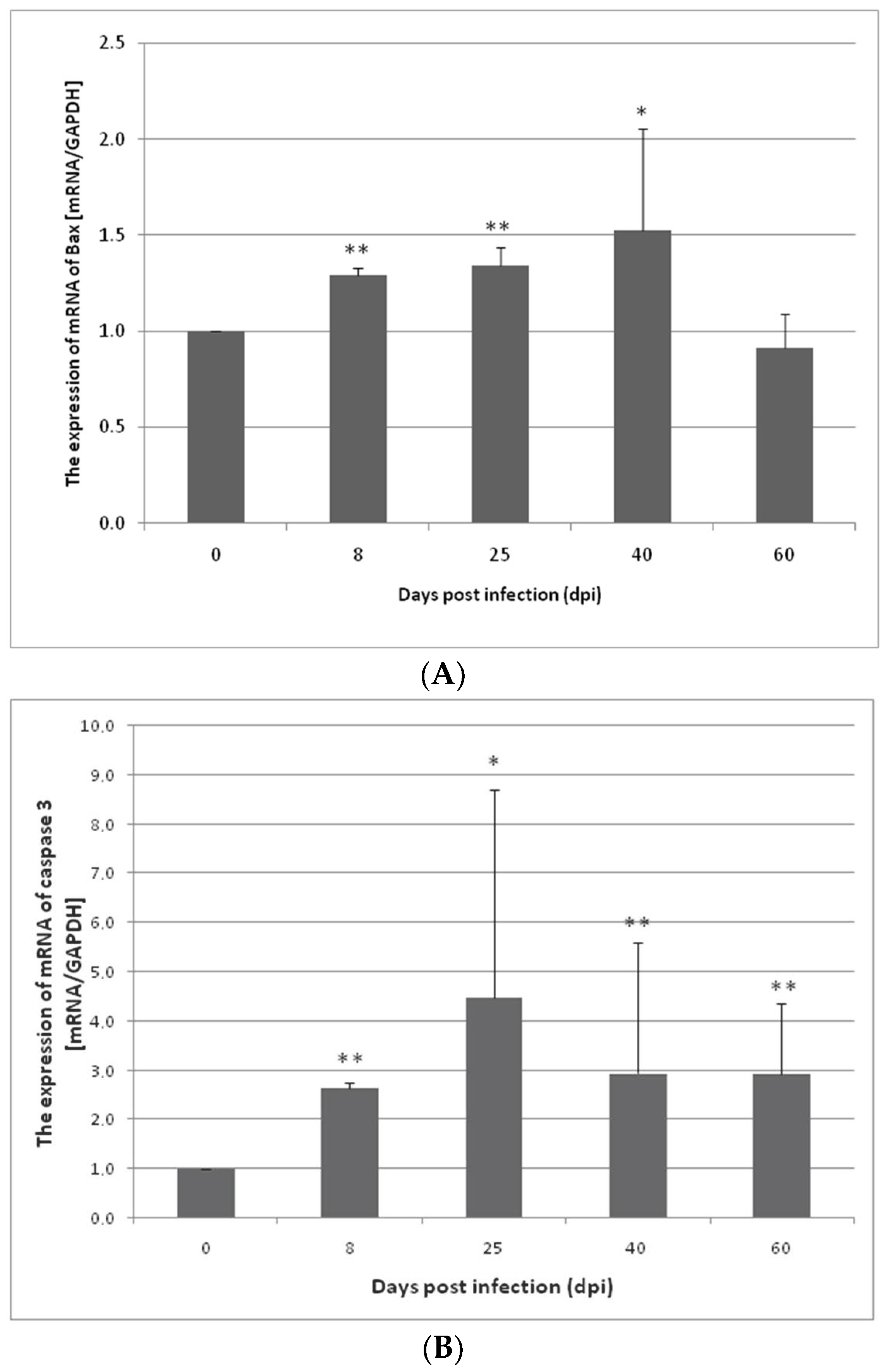
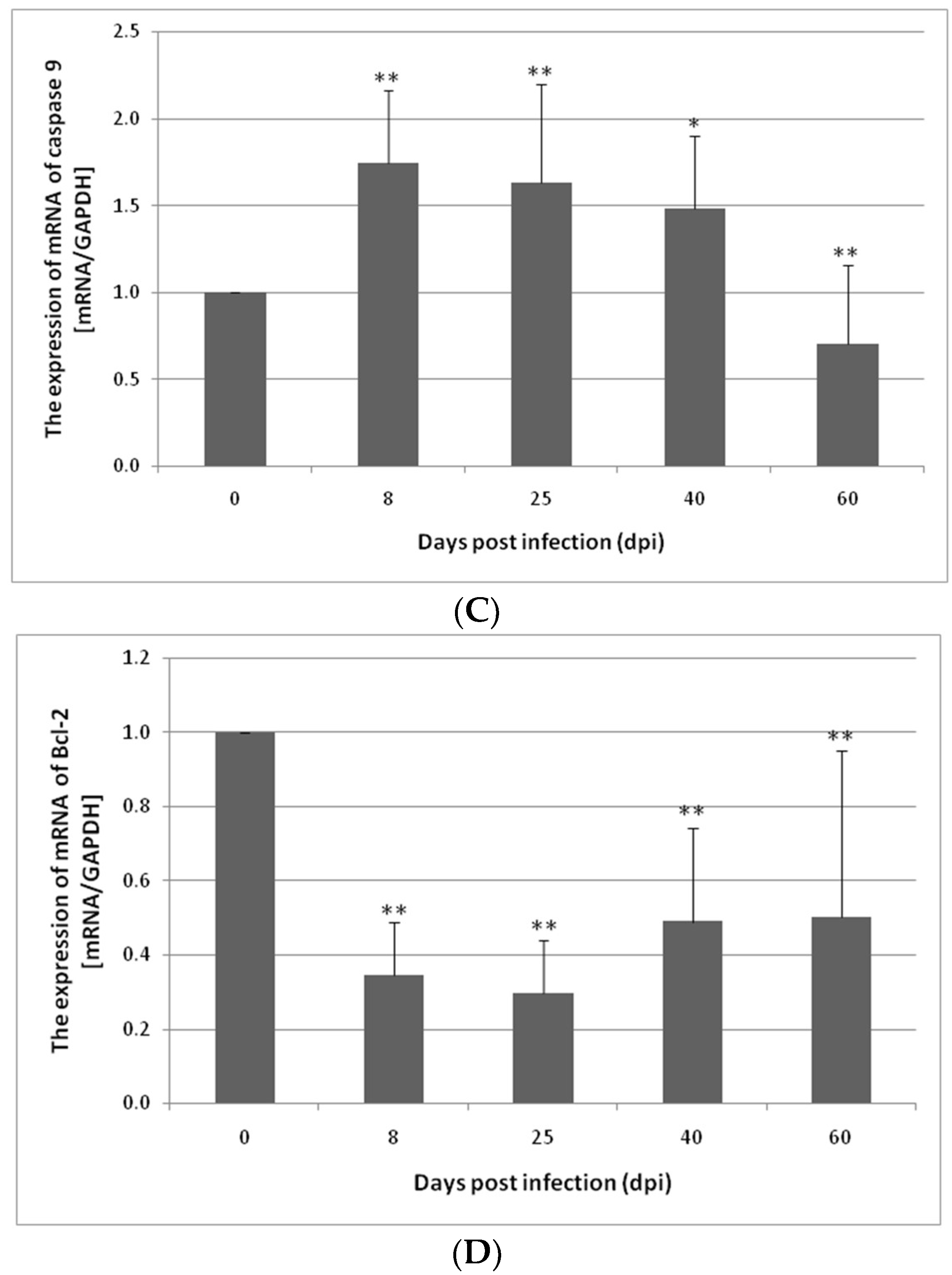
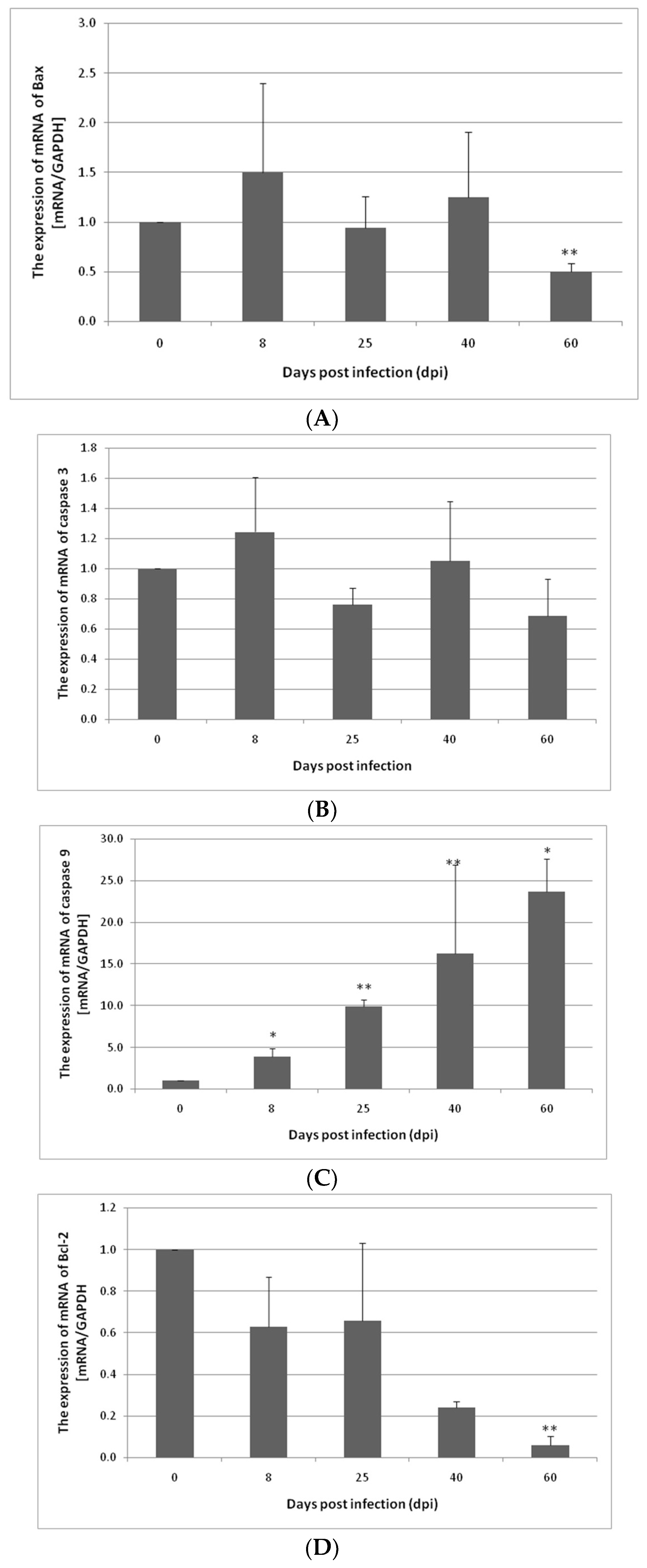
3.1. Changes in Apoptosis Gene Expression in the Small and Large Intestines of the Rat during H. diminuta Infection
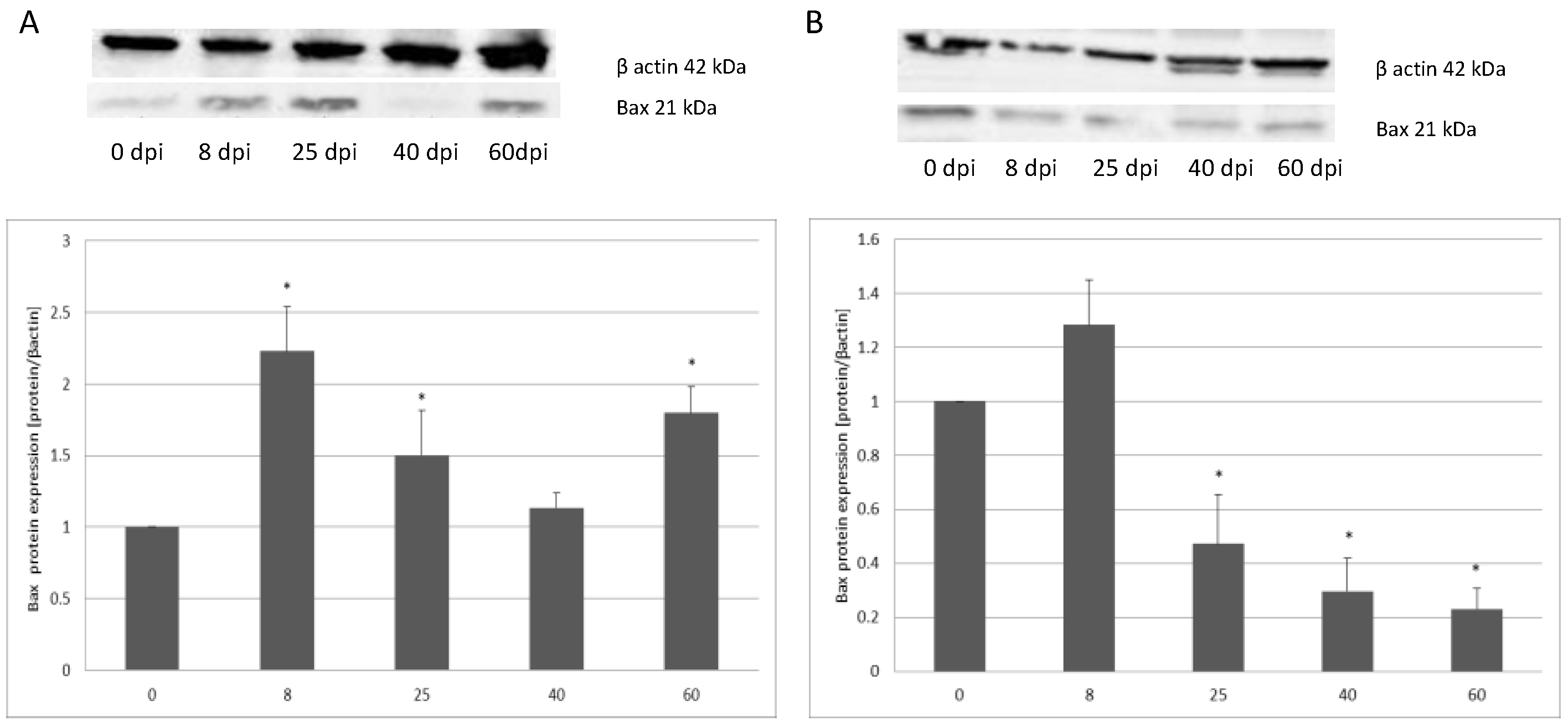
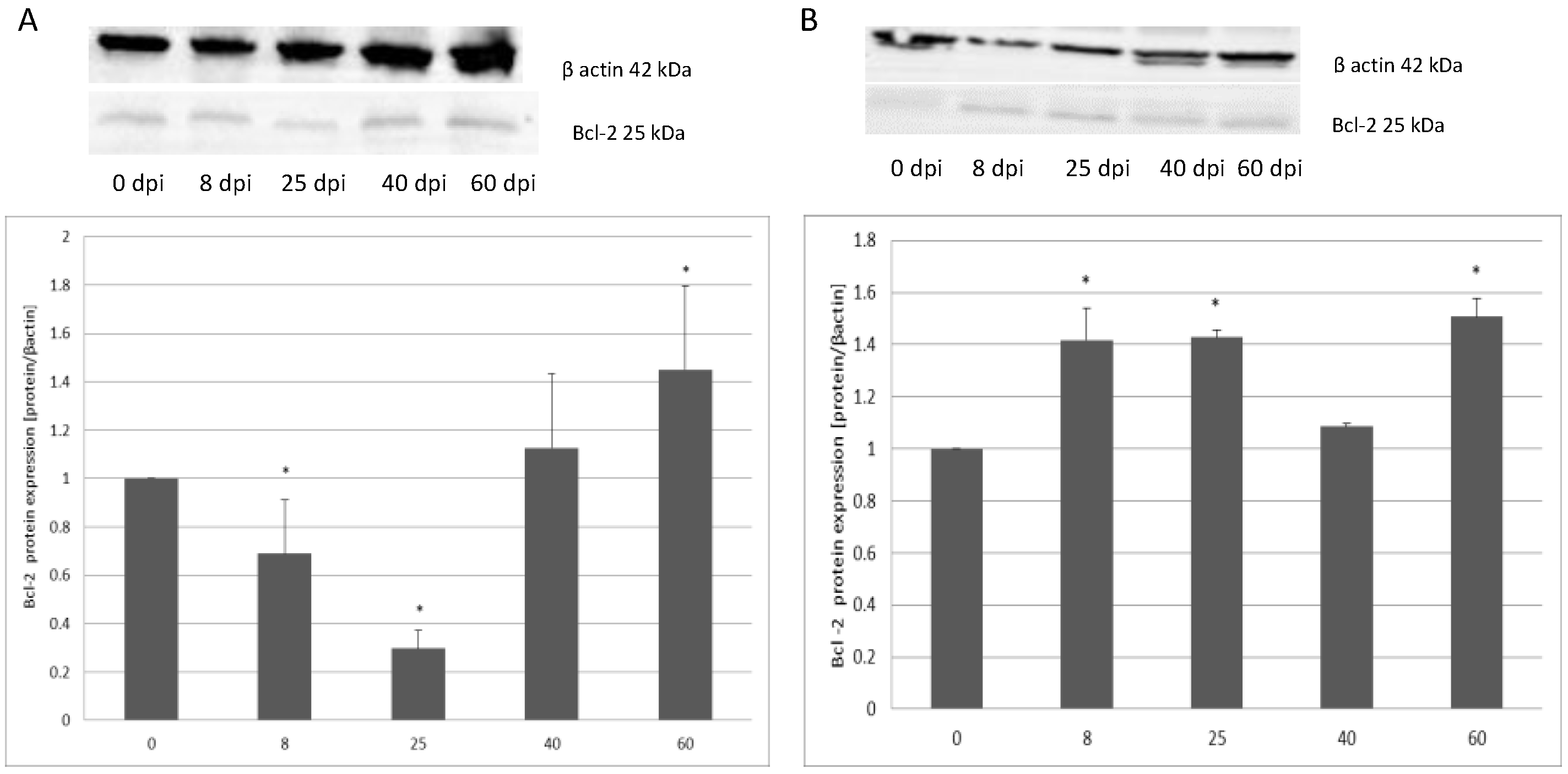
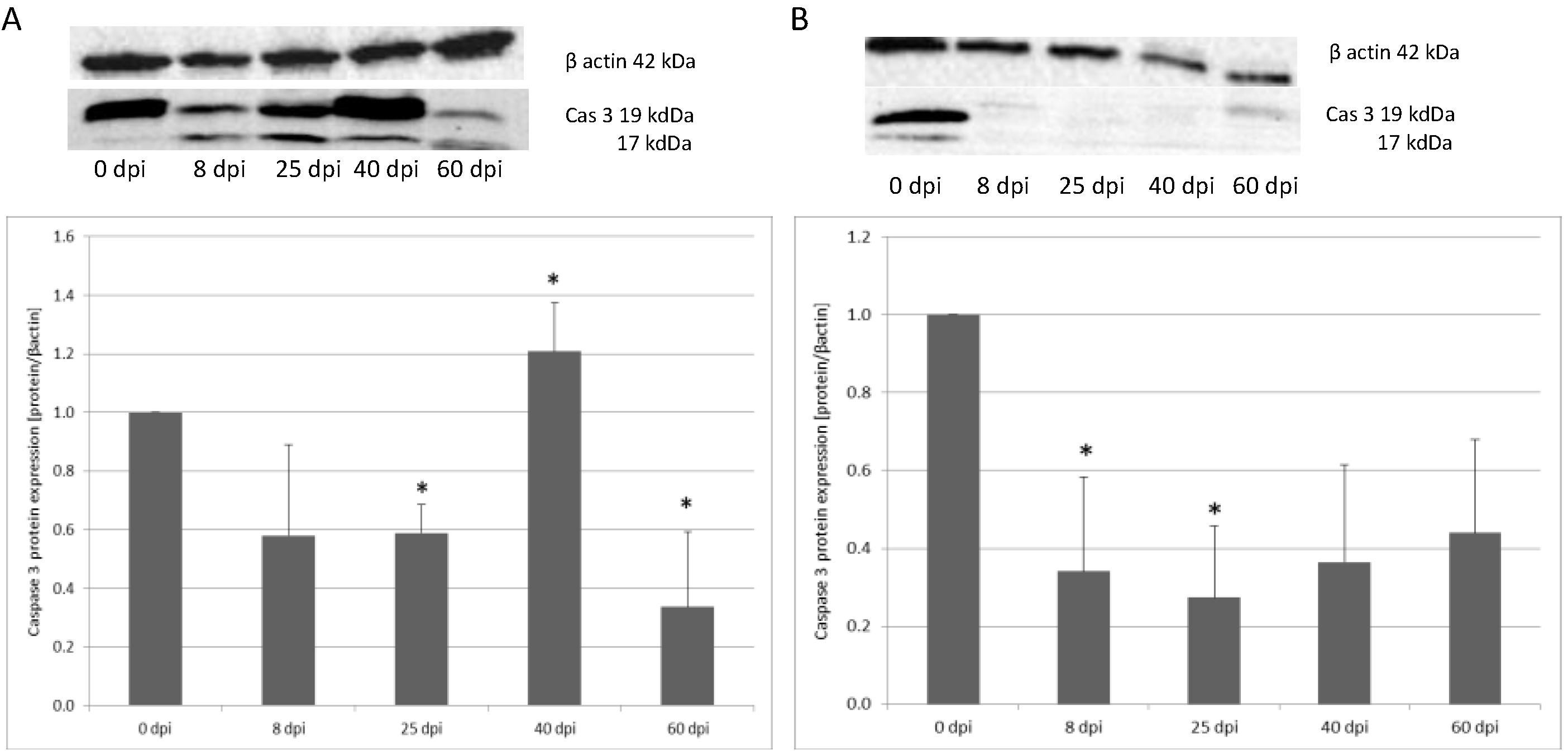
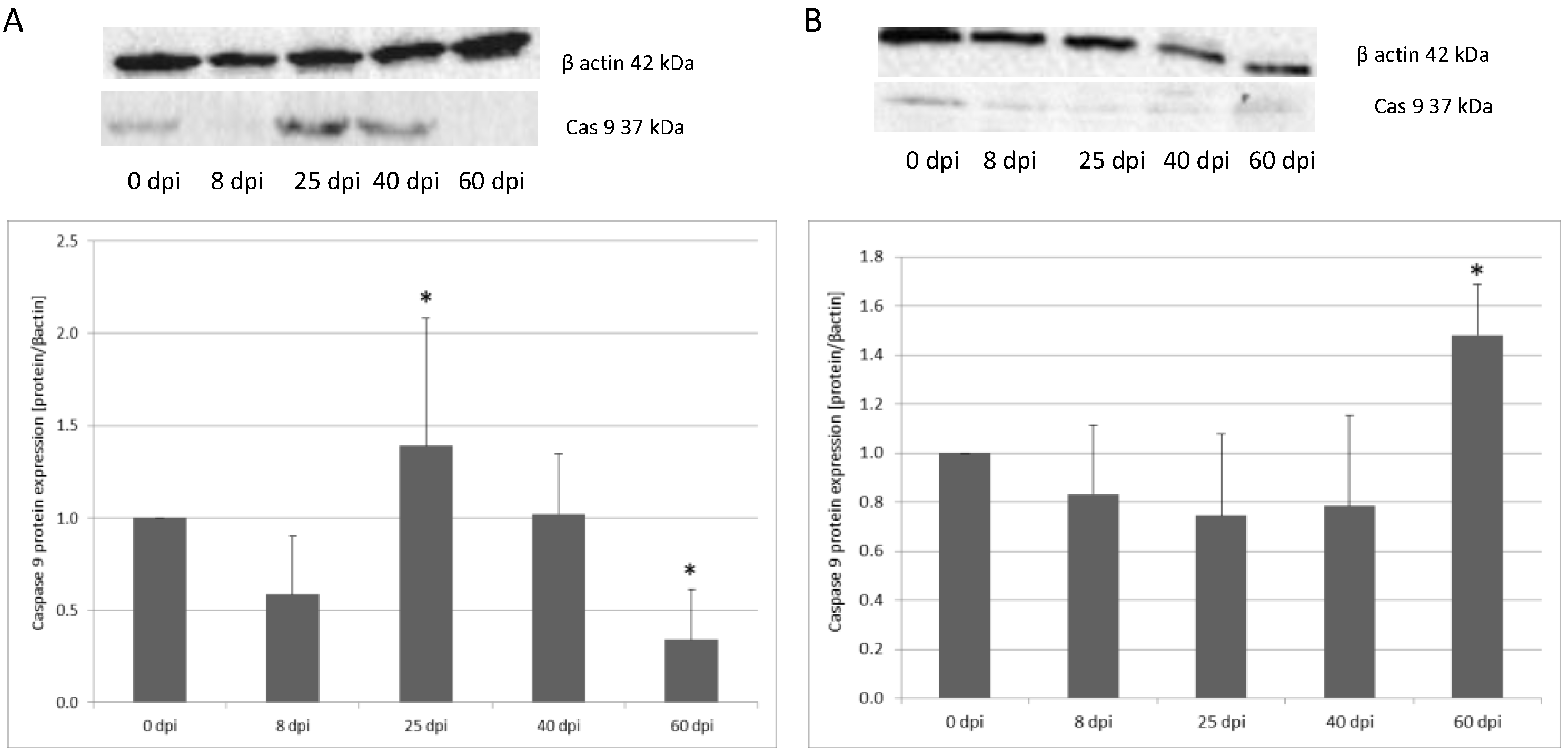
3.2. Changes in Apoptotic Protein Expression in the Small and Large Intestine of the Rat during H. diminuta Infection
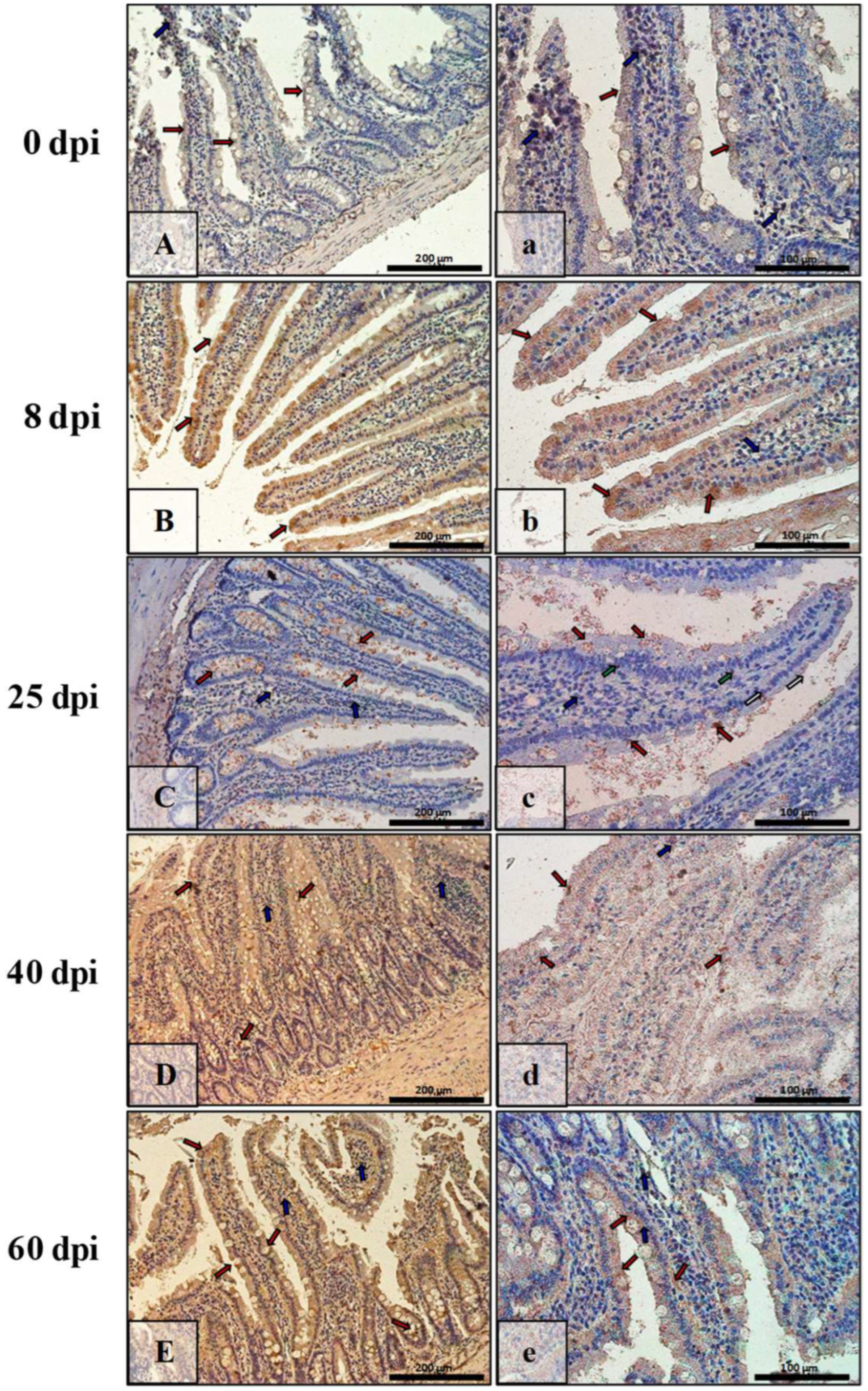
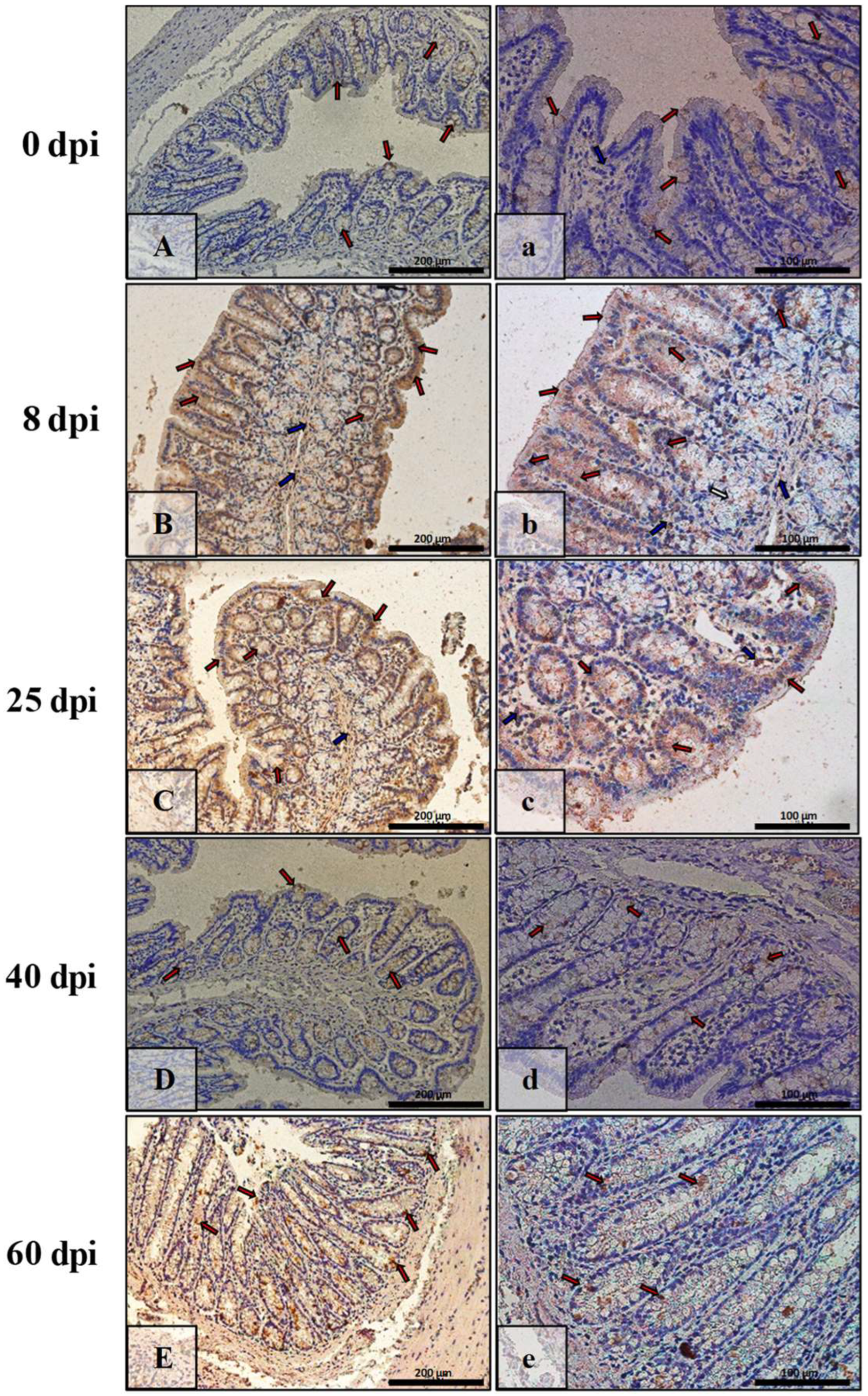
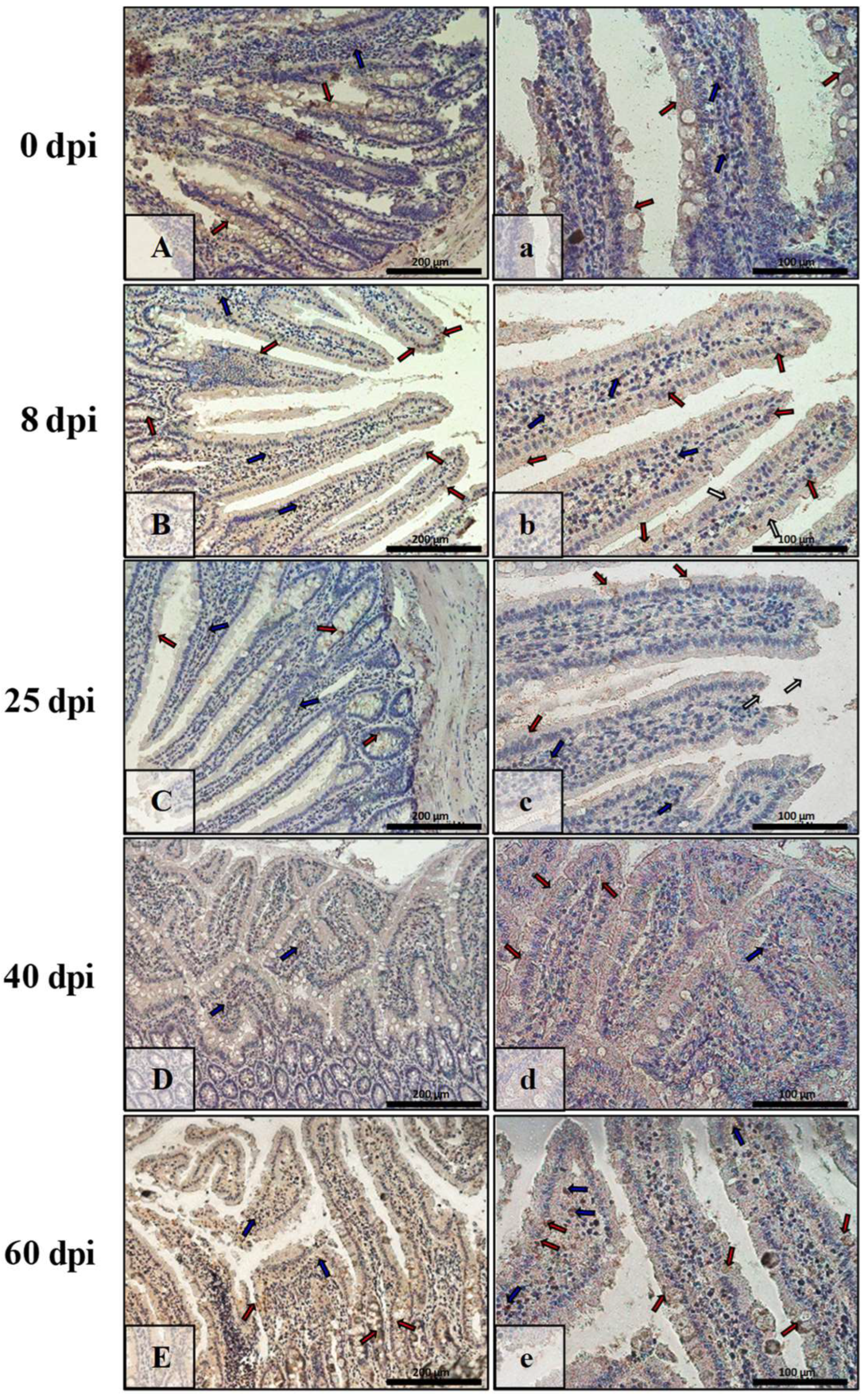
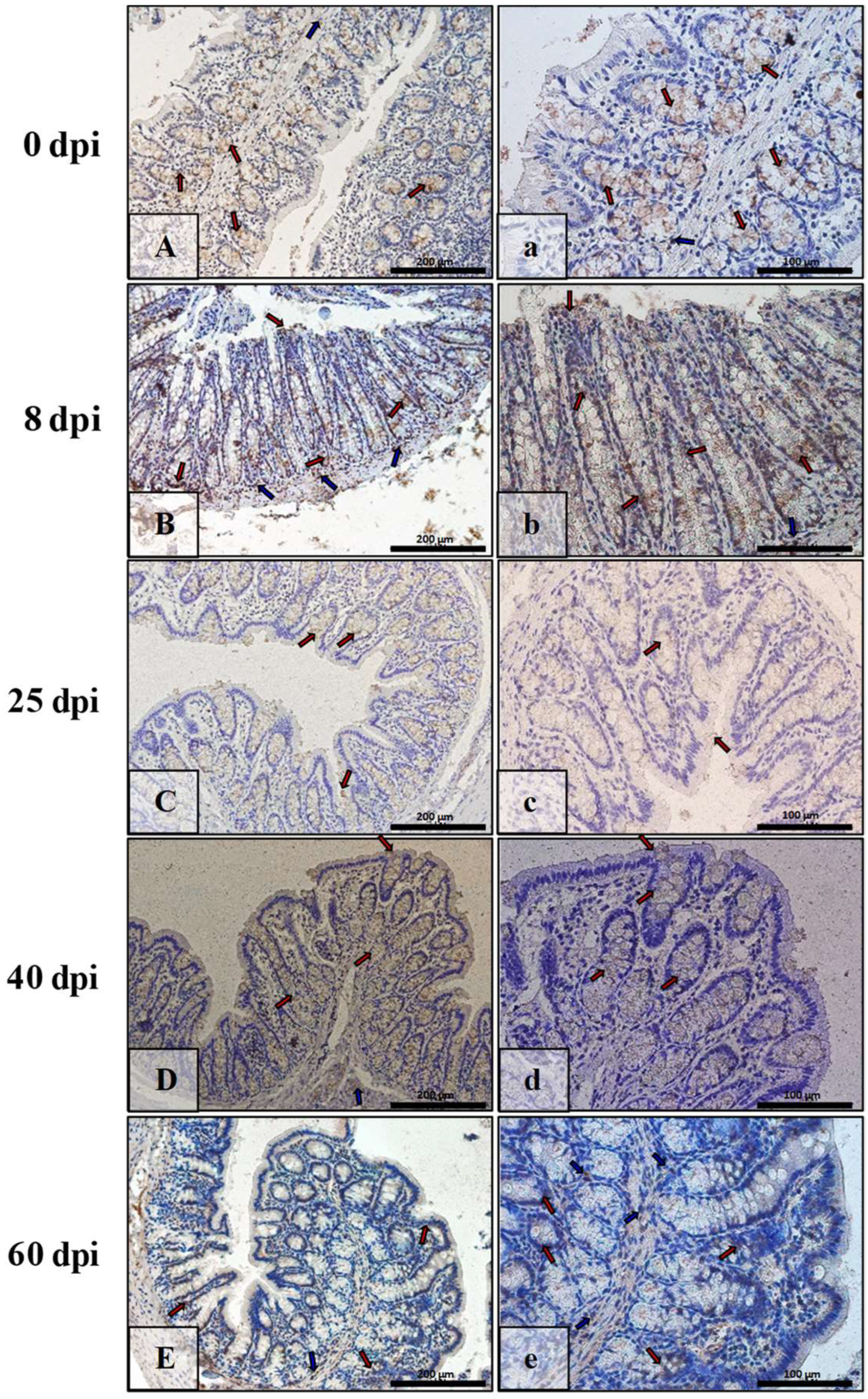
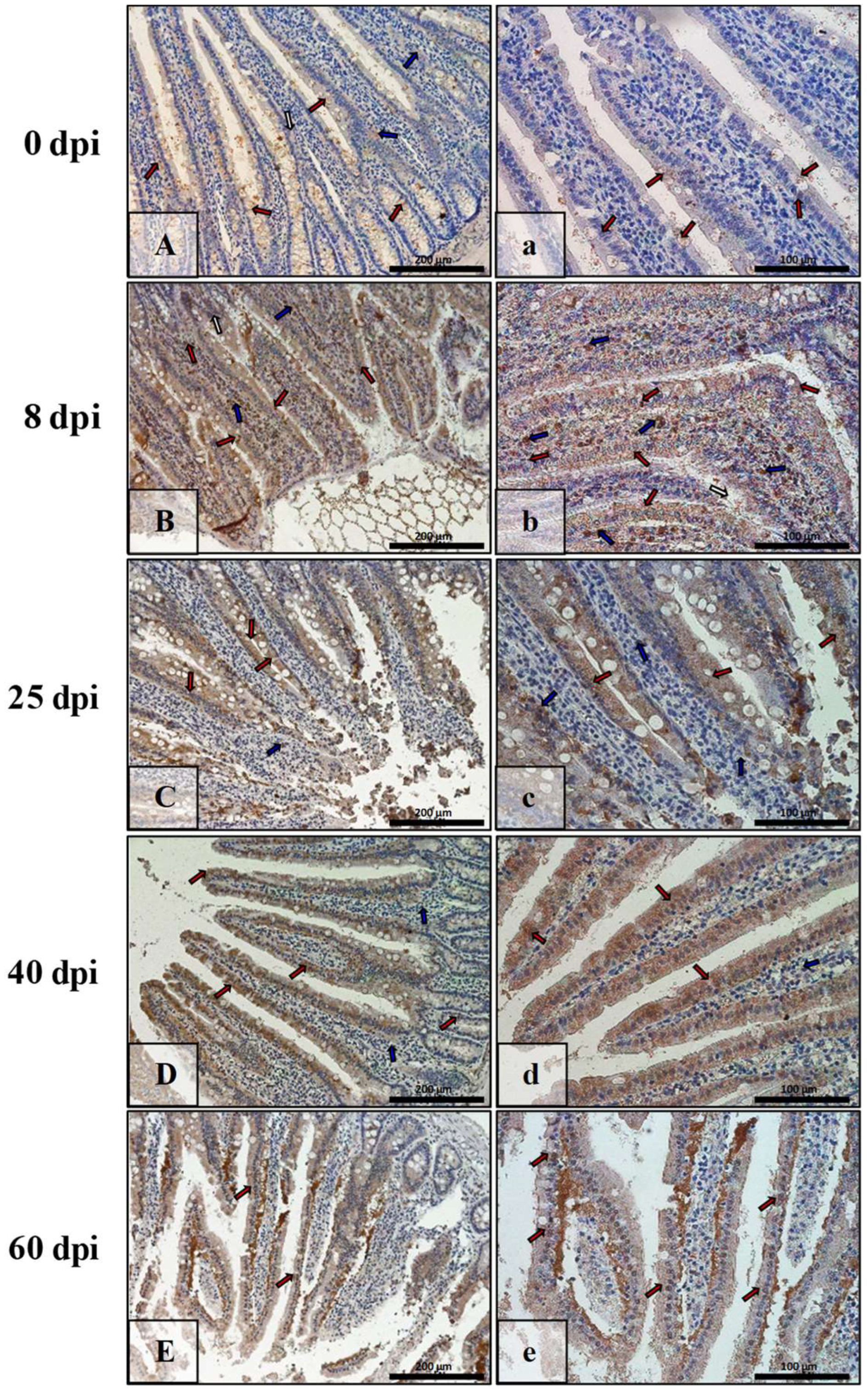
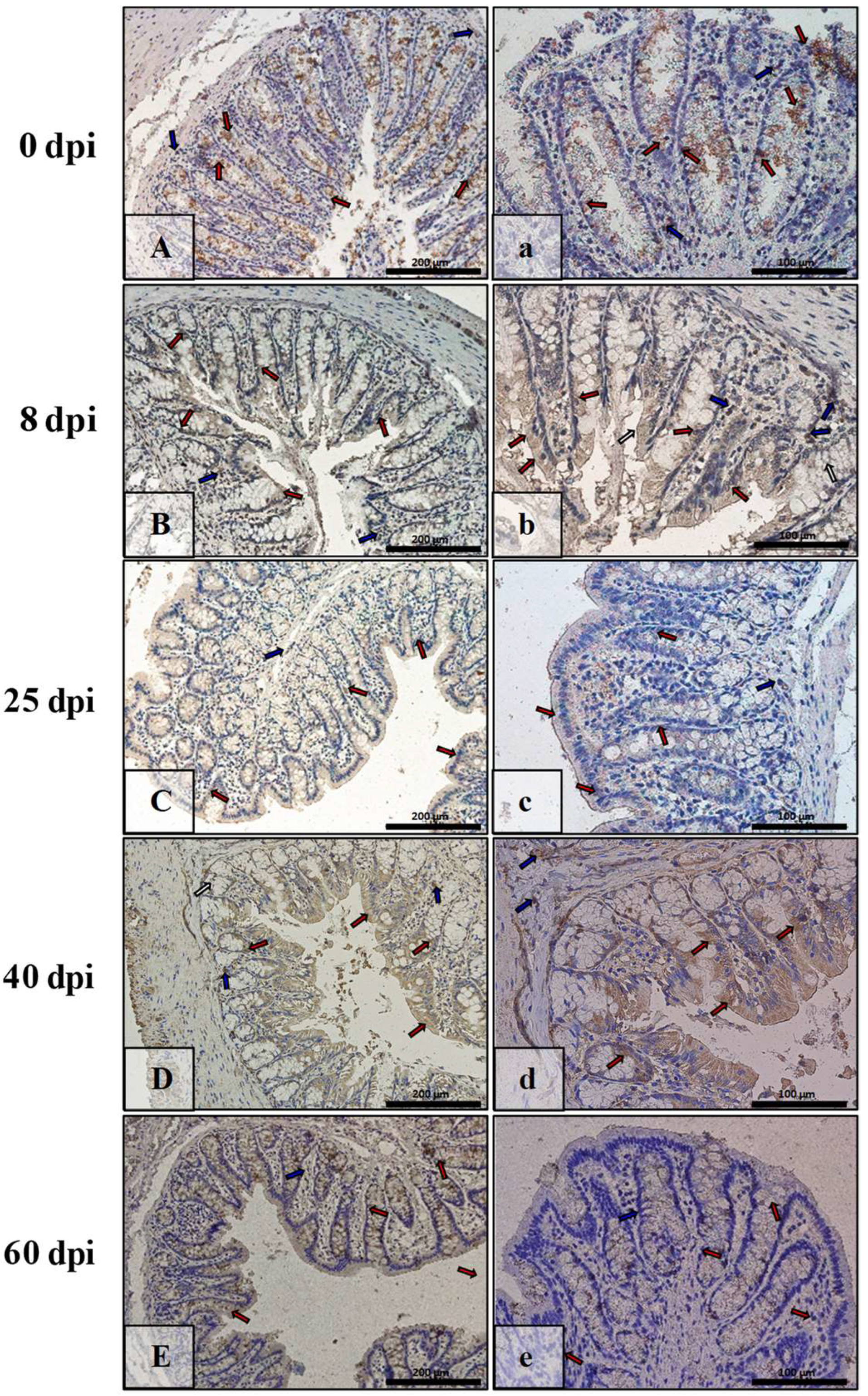
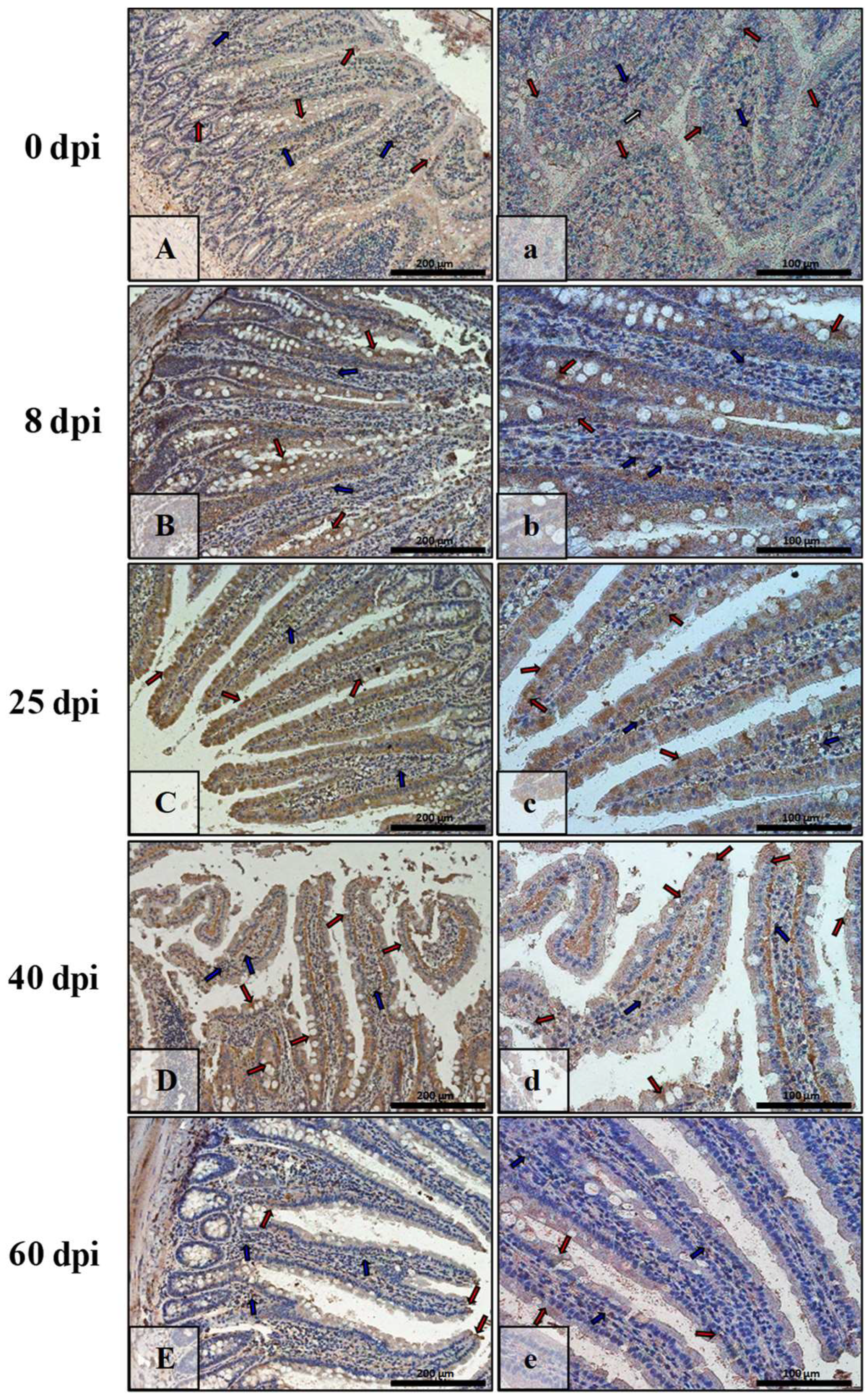
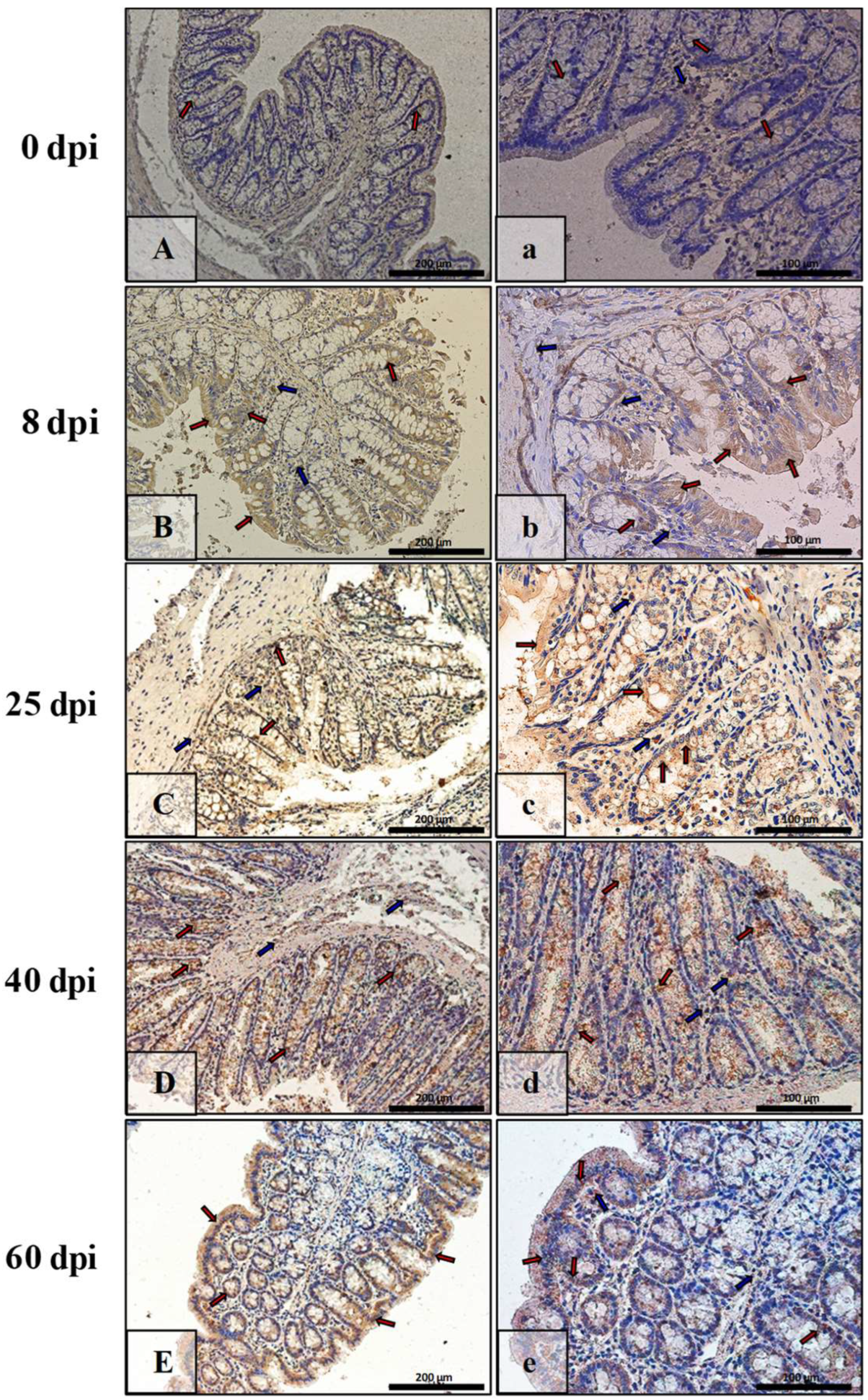
3.3. Immunohistochemical Analysis of Bax, Bcl-2, Cas-3, and Cas-9
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Panti-May, J.A.; Rodríguez-Vivas, R.I.; García-Prieto, L. Worldwide overview of human infections with Hymenolepis diminuta. Parasitol. Res. 2020, 119, 1997–2004. [Google Scholar] [CrossRef]
- Kapczuk, P.; Chlubek, D.; Baranowska-Bosiacka, I. Epidemiological and clinical characteristic of Hymenolepis diminuta infection—review of current literature. Pomeranian J. Life Sci. 2020, 66, 32–38. [Google Scholar] [CrossRef]
- Fal, W.; Czaplicka, H. Effect of experimental hymenolepiasis on various tissue reactions in rats. Wiad. Parazytol. 1991, 37, 331–342. [Google Scholar]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites: Masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef]
- Smallwood, T.B.; Giacomin, P.R.; Loukas, A.; Mulvenna, J.P.; Clark, R.J.; Miles, J.J. Helminth Immunomodulation in Autoimmune Disease. Front. Immunol. 2017, 8, 453. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wu, L.; Weng, R.; Zheng, W.; Wu, Z.; Lv, Z. Therapeutic potential of helminths in autoimmune diseases: Helminth-derived immune-regulators and immune balance. Parasitol. Res. 2017, 116, 2065–2074. [Google Scholar] [CrossRef]
- Goswami, R.; Singh, S.M.; Kataria, M.; Somvanshi, R. Clinicopathological studies on spontaneous Hymenolepis diminuta infection in wild and laboratory rats. Braz. J. Vet. Pathol. 2011, 4, 103–111. [Google Scholar]
- Kosik-Bogacka, D.I.; Kolasa, A. Histopathological changes in small and large intestines during hymenolepidosis in rats. Folia Biol. 2012, 60, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Kosik-Bogacka, D.; Baranowska-Bosiacka, I.; Salamatin, R. Hymenolepis diminuta: Effect of infection on ion transport in colon and blood picture of rats. Exp. Parasitol. 2010, 124, 285–294. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Kolasa, A.; Baranowska-Bosiacka, I.; Marchlewicz, M. Hymenolepis diminuta: The effects of infection on transepithelial ion transport and tight junctions in rat intestines. Exp. Parasitol. 2011, 127, 398–404. [Google Scholar] [CrossRef]
- Bikopoulos, G.J.; Hoque, T.; Webb, R.A. Infection with the cestode Hymenolepis diminuta induces changes in acetylcholine metabolism and muscarinic receptor mRNA expression in the rat jejunum. Parasitol. Res. 2006, 99, 231–237. [Google Scholar] [CrossRef]
- Shute, A.; Wang, A.; Jayme, T.S.; Strous, M.; McCoy, K.D.; Buret, A.G.; McKay, D.M. Worm expulsion is independent of alterations in composition of the colonic bacteria that occur during experimental Hymenolepis diminuta-infection in mice. Gut Microbes 2020, 11, 497–510. [Google Scholar] [CrossRef]
- McKenney, E.A.; Williamson, L.; Yoder, A.D.; Rawls, J.F.; Bilbo, S.D.; Parker, W. Alteration of the rat cecal microbiome during colonization with the helminth Hymenolepis diminuta. Gut Microbes 2015, 69, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Gill, N.; Shaikh, A.A.; Khan, M.M.; Memon, M.S. Influence on intestinal cestodes on the blood picture of brown rats (Rattus norvegicus) of Hyderabad Sindh, Pakistan. Pak. J. Biol. Sci. 2007, 10, 4479–4484. [Google Scholar] [CrossRef] [Green Version]
- Sommerfelt, I.E.; Santillan, G.; Mira, G.; Ribicich, M.; De Torres, R. Toxoca racanis infection in a pig model: Immunological, haematological and blood biochemistry responses. J. Helminthol. 2006, 80, 73–77. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Baranowska-Bosiacka, I.; Lanocha, N.; Wandurska-Nowak, E.; Gutowska, I.; Salamatin, R.; Jagodziński, P.P. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes and protein (TLR3 and TLR9) in the small and large intestines of rats. Exp. Parasitol. 2014, 145, 61–67. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Czernomysy-Furowicz, D.; Lanocha, N.; Wandurska-Nowak, E.; Salamatin, R.; Jagodziński, P.P. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes (TLR2 and TLR4) in the small and large intestines of rats. Part II. Exp. Parasitol. 2013, 135, 437–445. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Salamatin, R.; Jagodziński, P.P.; Wandurska-Nowak, E. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes (TLR2 and TLR4) in the small and large intestines of rats. Exp. Parasitol. 2012, 130, 261–266. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.I.; Baranowska-Bosiacka, I.; Kolasa-Wołosiuk, A.; Lanocha-Arendarczyk, N.; Gutowska, I.; Korbecki, J.; Namięta, H.; Rotter, I. The inflammatory effect of infection with Hymenolepis diminuta via the increased expression and activity of COX-1 and COX-2 in the rat jejunum and colon. Exp. Parasitol. 2016, 169, 69–76. [Google Scholar] [CrossRef]
- Ahmad, A.K.; Abdel-Hafeez, E.H.; Kamal, A.M. Some studies on spontaneous Hymenolepis diminuta infection in laboratory rats. J. Egypt Soc. Parasitol. 2015, 45, 115–124. [Google Scholar]
- Młocicki, D.; Sulima, A.; Bień, J.; Näreaho, A.; Zawistowska-Deniziak, A.; Basałaj, K.; Sałamatin, R.; Conn, D.B.; Savijoki, K. Immunoproteomics and Surfaceomics of the Adult Tapeworm Hymenolepis diminuta. Front. Immunol. 2018, 12, 2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bień, J.; Sałamatin, R.; Sulima, A.; Savijoki, K.; Conn, D.B.; Nareaho, A.; Młocicki, D. Mass spectrometry analysis of the excretory-secretory (E-S) products of the model cestode Hymenolepis diminuta reveals their immunogenic properties and the presence of New E-S proteins in cestodes. Acta Parasitol. 2016, 61, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Baś, M.; Cywińska, A.; Sokołowska, J.; Krzyżowska, M. Apoptosis–programmedcelldeath. Part III. The role of apoptosis in physiology and pathology. Zycie Weter. 2004, 79, 671–675. [Google Scholar]
- Stępień, A.; Izdebska, M.; Grzanka, A. The types of cell death. Postepy Hig. Med. Dosw. 2007, 61, 420–428. [Google Scholar]
- Hordyjewska, A.; Pasternak, K. Apoptotic Death of the Cell. Adv. Clin. Exp. Med. 2005, 14, 545–554. [Google Scholar]
- Nijhawan, D.; Honarpour, N.; Wang, X. Apoptosis in neural development and disease. Annu. Rev. Neurosci. 2000, 23, 73–87. [Google Scholar] [CrossRef]
- Oppenheim, R.W.; Flavell, R.A.; Vinsant, S.; Prevette, D.; Kuan, C.Y.; Rakic, P. Programmed cell death of developing mammalian neurons after genetic deletion of caspases. J. Neurosci. 2001, 21, 4752–4760. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lenardo, M.J. Roles of caspases in apoptosis, development, and cytokine maturation revealed by homozygous gene deficiencies. J Cell Sci. 2000, 113, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Epple, H.J.; Schneider, T.; Troeger, H.; Kunkel, D.; Allers, K.; Moos, V.; Amasheh, M.; Loddenkemper, C.; Fromm, M.; Zeitz, M.; et al. Impairment of the intestinal barrier is evident in untreated but absent in suppressively treated HIV-infected patients. Gut 2009, 58, 220–227. [Google Scholar] [CrossRef]
- Matysiak-Budnik, T.; Coffin, B.; Lavergne-Slove, A.; Sabate, J.M.; Mégraud, F.; Heyman, M. Helicobacter pylori increases the epithelial permeability to a food antigen in human gastric biopsies. Am. J. Gastroenterol. 2004, 99, 225–232. [Google Scholar] [CrossRef]
- Kapczuk, P.; Kosik-Bogacka, D.; Kupnicka, P.; Metryka, E.; Simińska, D.; Rogulska, K.; Skórka, M.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. The influence of selected gastrointestinal parasites on apoptosis in intestinal epithelial cells. Biomolecules 2020, 10, 674. [Google Scholar] [CrossRef] [PubMed]
- Buret, A.A.; Mitchell, K.; Muench, D.G.; Scott, K.G. Giardia lamblia disrupts tight junctional ZO-1 and increases permeability in nontransformed human small intestinal epithelilal monolayers: Effects of epidermal growth factor. Parasitology 2002, 125, 11–19. [Google Scholar] [CrossRef]
- Shimizu, S.; Tsujimoto, Y. Proapoptotic BH3-only Bcl-2 family members induce cytochrome c release, but not mitochondrial membrane potential loss, and do not directly modulate voltage-dependent anion channel activity. Proc. Natl. Acad. Sci. USA 2000, 97, 577–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, H.; Henzel, W.J.; Liu, X.; Lutschg, A.; Wang, X. Apaf-1, a Human Protein Homologous to C. elegans CED-4, Participates in Cytochrome c–Dependent Activation of Caspase-3. Cell 1997, 90, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Panaro, M.A.; Cianciulli, A.; Mitolo, V.; Mitolo, C.I.; Acquafredda, A.; Brandonisio, O.; Cavallo, P. Caspase-dependent apoptosis of the HCT-epithelial cell line induced by the parasite Giardia intestinalis. FEMS Immunol. Med. Microbiol. 2007, 51, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Kapczuk, P.; Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Gutowska, I.; Kupnicka, P.; Chlubek, D.; Baranowska-Bosiacka, I. Selected Molecular Mechanisms Involved in the Parasite–Host System Hymenolepis diminuta–Rattus norvegicus. Int. J. Mol. Sci. 2018, 19, 2435. [Google Scholar] [CrossRef] [Green Version]
- Su, C.C.; Chen, J.Y.F.; Din, Z.H.; Su, J.H.; Yang, Z.Y.; Chen, Y.J.; Wang, R.Y.; Wu, Y.J. 13-acetoxysarcocrassolide induces apoptosis on human gastric carcinoma cells through mitochondria-related apoptotic pathways: P38/JNK activation and PI3K/AKT suppression. Mar. Drugs 2014, 12, 5295–5315. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Yang, Y.; Ma, L.; Ma, B.; Ren, L.; Guo, L.; Wang, W.; Zhang, Y.; Zhao, Z.; Cui, M. In vivo and in vitro induction of the apoptotic effects of oxysophoridine on colorectal cancer cells via the Bcl-2/Bax/caspase-3 signaling pathway. Oncol. Lett. 2017, 14, 8000–8006. [Google Scholar] [CrossRef]
- Zhao, Y.; Jing, Z.; Lv, J.; Zhang, Z.; Lin, J.; Cao, X.; Zhao, Z.; Liu, P.; Mao, W. Berberine activates caspase-9/cytochrome c-mediated apoptosis to suppress triple-negative breast cancer cells in vitro and in vivo. Biomed. Pharmacother. 2017, 95, 18–24. [Google Scholar] [CrossRef]
- Chin, A.C.; Teoh, D.A.; Scott, K.G.; Meddings, J.B.; Macnaughton, W.K.; Buret, A.G. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infect. Immun. 2002, 70, 3673–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roxstrom-Lindquist, K.; Ringqvist, E.; Palm, D.; Svärd, S. Giardia lamblia-indeced changes in gene expression in differentiated Caco-2 human intestinal epithelial cells. Infect. Immun. 2005, 73, 8204–8208. [Google Scholar] [CrossRef] [Green Version]
- Troeger, H.; Epple, H.J.; Schneider, T.; Wahnschaffe, U.; Ullrich, R.; Burchard, G.D.; Jelinek, T.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Effect of chronic Giardia lamblia infection on epithelial transport and barrier function in human duodenum. Gut 2007, 56, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Hayes, K.S.; Cliffe, L.J.; Bancroft, A.J.; Forman, S.P.; Thompson, S.; Booth, C.; Grencis, R.K. Chronic Trichurismuris infection causes neoplastic change in the intestine and exacerbates tumour formation in APC min/+ mice. PLoS Negl. Trop. Dis. 2017, 11, e0005708. [Google Scholar] [CrossRef] [Green Version]
- Boettner, D.R.; Petri, W. Entamoeba histolytica activates host cell caspases during contact-dependent cell killing. Curr. Top. Microbiol. Immunol. 2005, 289, 175–184. [Google Scholar]
- El-Ashram, S.; Al Nasr, I.; Mehmood, R.; Çağdaş, D.; Hu, M.; He, L.; Suo, X. Cell death offers exceptional cellular and molecular windows for pharmacological interventions in protozoan parasites. Integr. Mol. Med. 2017, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Hyoh, Y.; Nishida, M.; Tegoshi, T.; Yamada, M.; Uchikawa, R.; Matsuda, S.; Arizono, N. Enhancement of apoptosis with loss of cellular adherence in the villus epithelium of the small intestine after infection with the nematode Nippostrongylus brasiliensis in rats. Parasitology 1999, 119, 199–207. [Google Scholar] [CrossRef]
- Healer, J.; Ashall, F.; Maizels, R.M. Characterization of proteolytic enzymes from larval and adult Nippostrongylus brasiliensis. Parasitology 1991, 103, 305–314. [Google Scholar] [CrossRef]
- Nakazawa, M.; Yamada, M.; Uchikawa, R.; Arizono, N. Immunocytochemical localization of secretory acetylcholinesterase of the parasitic nematode Nippostrongylus brasiliensis. Cell Tissue Res. 1995, 280, 59–64. [Google Scholar] [CrossRef]
- Kuroda, A.; Uchikawa, R.; Matsuda, S.; Yamada, M.; Tegoshi, T.; Arizono, N. Up-regulation of Fas (CD95) and induction of apoptosis in intestinal epithelial cells by nematode-derived molecules. Infect. Immun. 2002, 70, 4002–4008. [Google Scholar] [CrossRef] [Green Version]
- Buret, A.G.; Chin, A.C.; Scott, K.G. Infection of human and bovine epithelial cells with Cryptosporidium andersoni induces apoptosis and disrupts tight junctional ZO-1: Effects of epidermal growth factor. Int. J. Parasitol. 2003, 33, 1363–1371. [Google Scholar] [CrossRef]
- Chen, X.M.; Gores, G.J.; Paya, C.V.; LaRusso, N.F. Cryptosporidium parvum induces apoptosis in biliary epithelia by a Fas/Fas ligand-dependent mechanism. Am. J. Physiol. 1999, 277, G599–G608. [Google Scholar]
- McCole, D.F.; Eckmann, L.; Laurent, F.; Kagnoff, M.F. Intestinal epithelial cell apoptosis following Cryptosporidium parvum infection. Infect. Immun. 2000, 68, 1710–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, R.; Gomez Morales, M.A.; Tosini, F.; Pozio, F. Cryptosporidium parvum at different developmental stages modulates host cell apoptosis in vitro. Infect. Immun. 2004, 72, 6061–6067. [Google Scholar] [CrossRef] [Green Version]
- Ojcius, D.M.; Perfettini, J.L.; Bonnin, A.; Laurent, F. Caspase-dependent apoptosis during infection with Cryptosporidium parvum. Microbes Infect. 1999, 1, 1163–1168. [Google Scholar] [CrossRef]
- Sasahara, T.; Maruyama, H.; Aoki, M.; Kikuno, R.; Sekiguchi, T.; Takahashi, A.; Satoh, Y.; Kitasato, H.; Takayama, Y.; Inoue, M. Apoptosis of intestinal crypt epithelium after Cryptosporidium parvum infection. J. Infect. Chemother. 2003, 9, 278–281. [Google Scholar] [CrossRef]
- Liu, J.; Enomoto, S.; Lancto, C.A.; Abrahamsen, M.S.; Rutherford, M.S. Inhibition of apoptosis in Cryptosporidium parvum-infected intestinal epithelial cells is dependent on survivin. Infect. Immun. 2008, 76, 3784–3792. [Google Scholar] [CrossRef] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward | Reverse | |
|---|---|---|
| GAPDH | ATG ACT CTA CCC ACG GCA AG | CTG GAA GAT GGT GAT GGG TT |
| Bcl-2 | ATC CAG GAT AAC GGA GGC TG | CAG GTA TGC ACC CAG AGT GA |
| Bax | GGG TGG TTG CCC TTT TCT ACT | AGT CCA GTG TCC AGC CCA TG |
| Cas-3 | AAT TCA AGG GAC GGG TCA TG | TGA CAC AAT ACA CGG GAT CT |
| Cas-9 | AGC CAG ATG CTG TCC CAT AC | CAG GAG ACA AAA CCT GGG AA |
| Marker | Part of Intestine | 0 dpi (Control) | 8 dpi | 25 dpi | 40 dpi | 60 dpi |
|---|---|---|---|---|---|---|
| BAX | small | +/− | +++ | ++ | + | +++ |
| large | +/− | +++ | +++ | + | ++ | |
| Bcl-2 | small | + | +/− | +/− | ++ | ++ |
| large | + | + | +/− | +/− | +/− | |
| Caspase-3 | small | +/− | + | ++ | +++ | ++ |
| large | + | ++ | + | ++ | + | |
| Caspase-9 | small | +/− | ++ | ++ | + | +/− |
| large | +/− | + | + | ++ | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapczuk, P.; Kosik-Bogacka, D.; Kupnicka, P.; Kopytko, P.; Tarnowski, M.; Kolasa, A.; Chlubek, D.; Baranowska-Bosiacka, I. Hymenolepis diminuta Infection Affects Apoptosis in the Small and Large Intestine. Int. J. Environ. Res. Public Health 2022, 19, 9753. https://doi.org/10.3390/ijerph19159753
Kapczuk P, Kosik-Bogacka D, Kupnicka P, Kopytko P, Tarnowski M, Kolasa A, Chlubek D, Baranowska-Bosiacka I. Hymenolepis diminuta Infection Affects Apoptosis in the Small and Large Intestine. International Journal of Environmental Research and Public Health. 2022; 19(15):9753. https://doi.org/10.3390/ijerph19159753
Chicago/Turabian StyleKapczuk, Patrycja, Danuta Kosik-Bogacka, Patrycja Kupnicka, Patrycja Kopytko, Maciej Tarnowski, Agnieszka Kolasa, Dariusz Chlubek, and Irena Baranowska-Bosiacka. 2022. "Hymenolepis diminuta Infection Affects Apoptosis in the Small and Large Intestine" International Journal of Environmental Research and Public Health 19, no. 15: 9753. https://doi.org/10.3390/ijerph19159753