
Antibiotic Susceptibility of Bacterial Pathogens That Infect Olive Flounder (Paralichthys olivaceus) Cultivated in Korea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. DNA Extraction
2.3. Polymerase Chain Reaction (PCR)
2.4. Antibiotic Susceptibility Testing
2.4.1. Broth Dilution Test
2.4.2. Disk Diffusion Test
3. Results
3.1. Identification of Strains
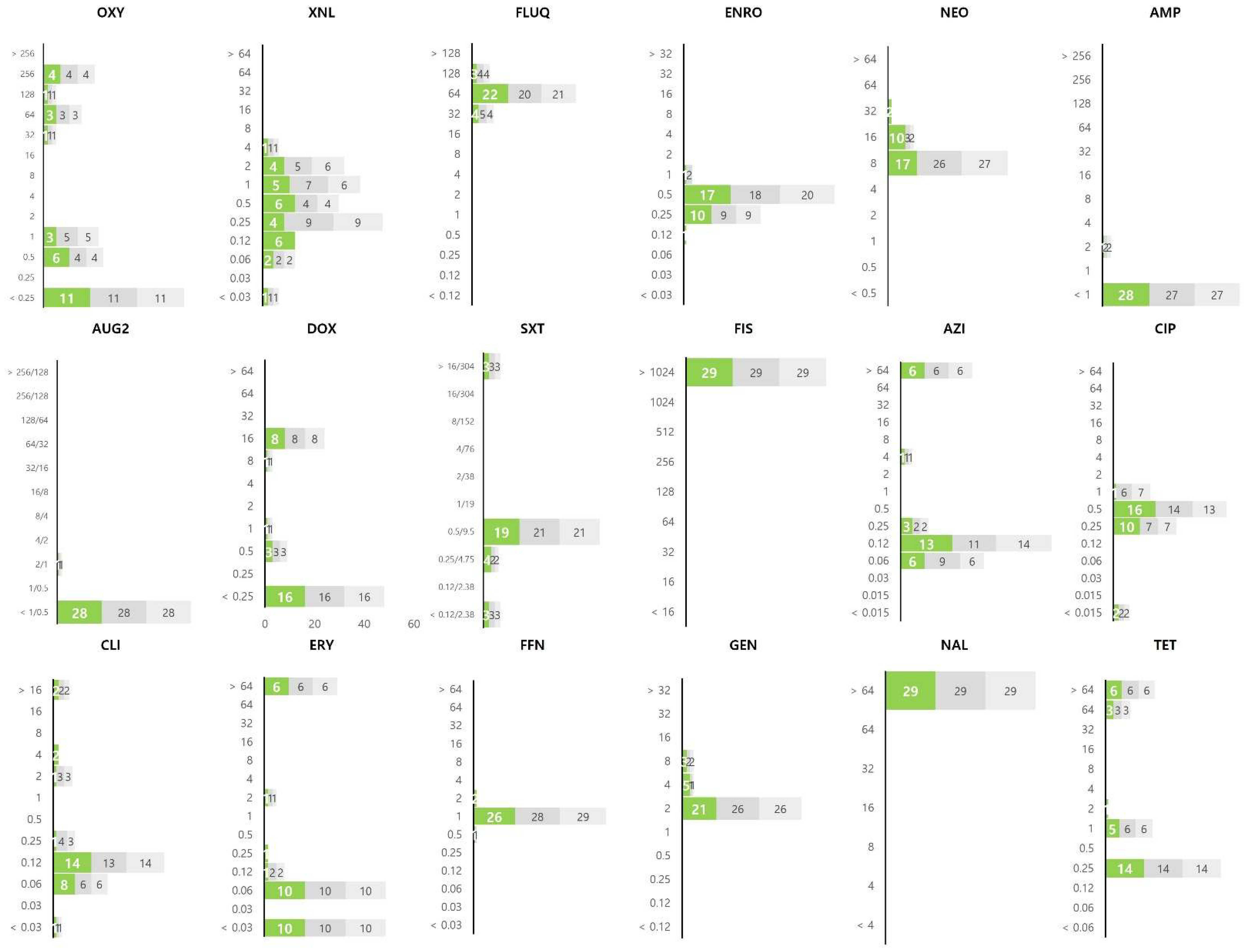
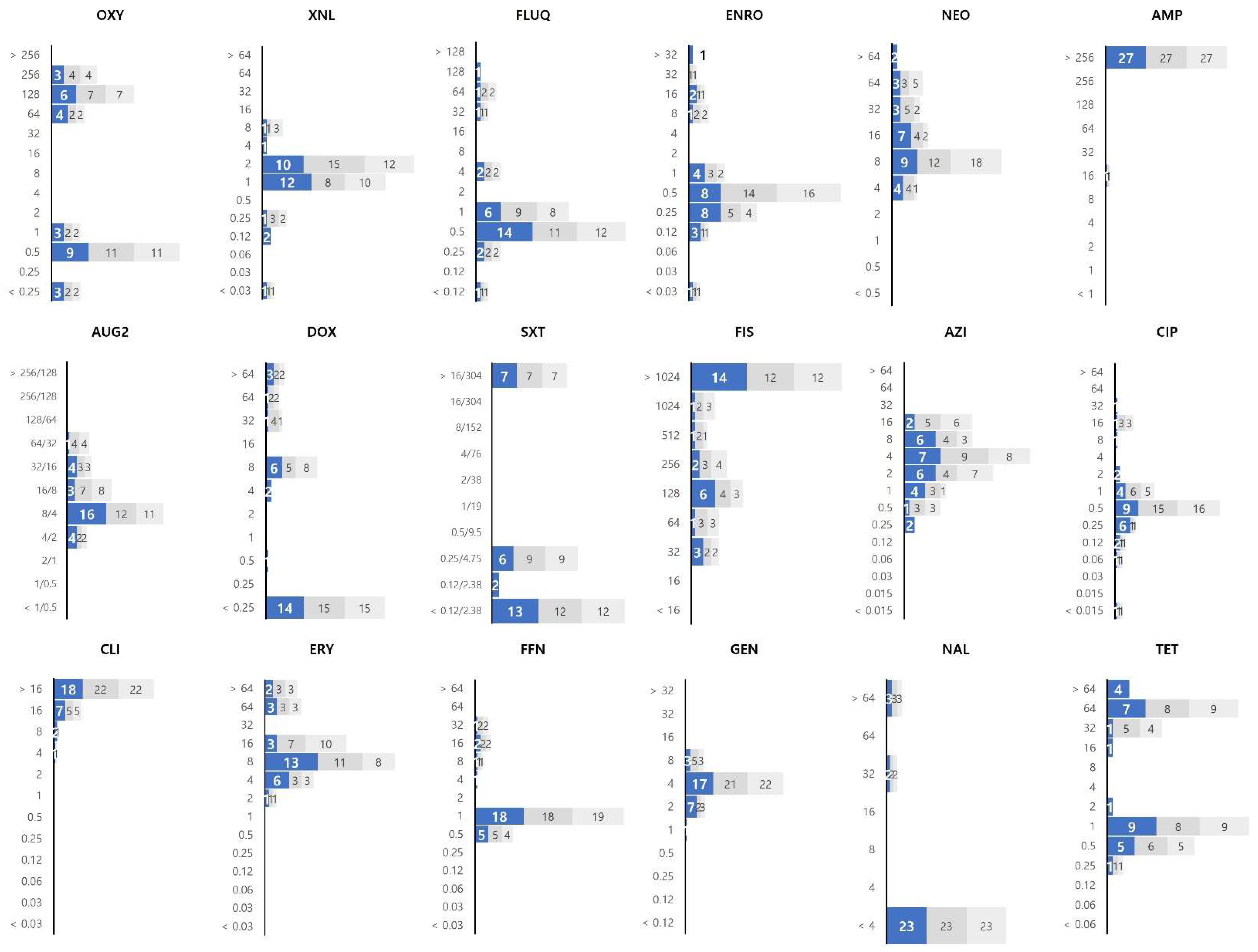
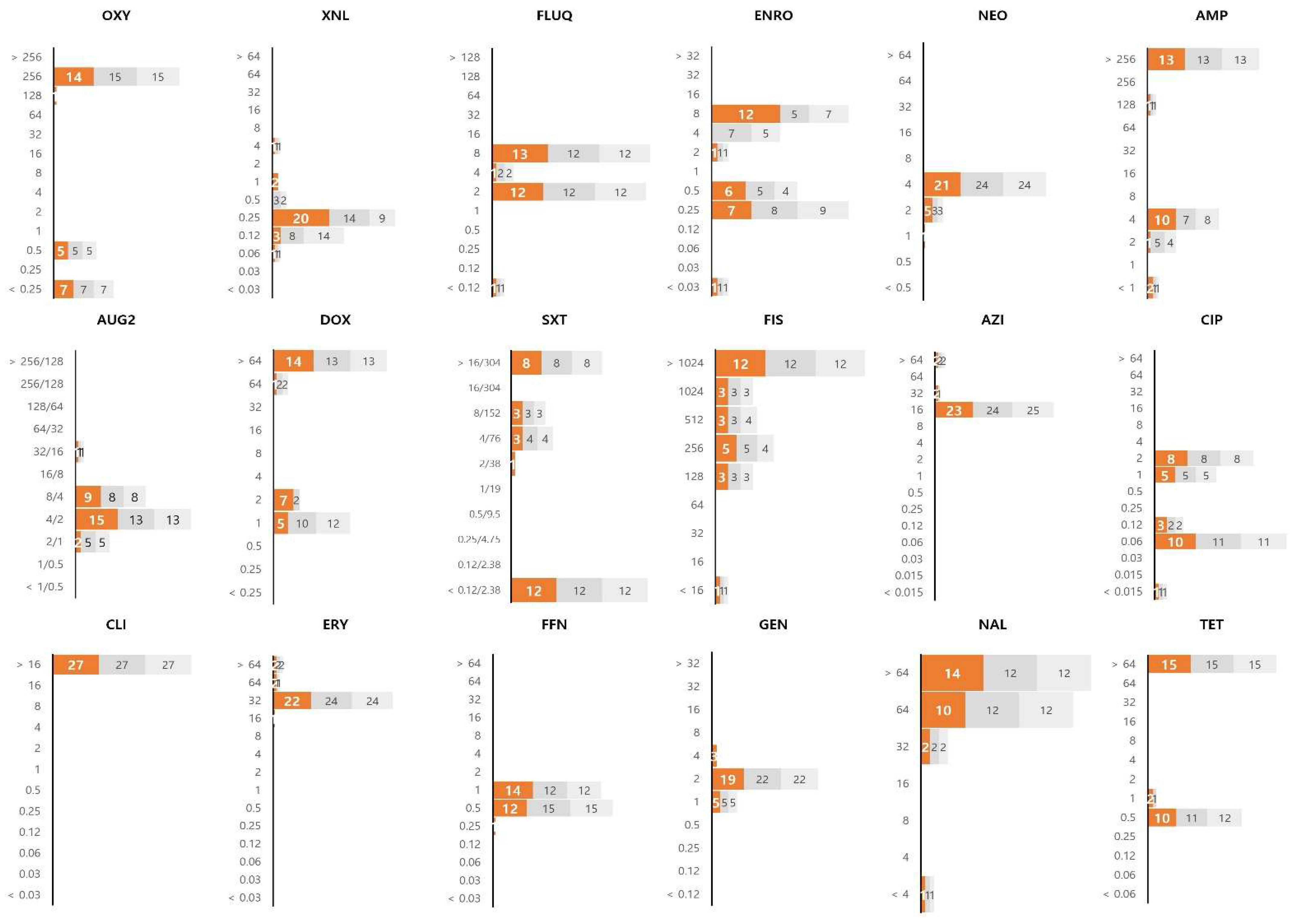
3.2. Distribution of MIC Values
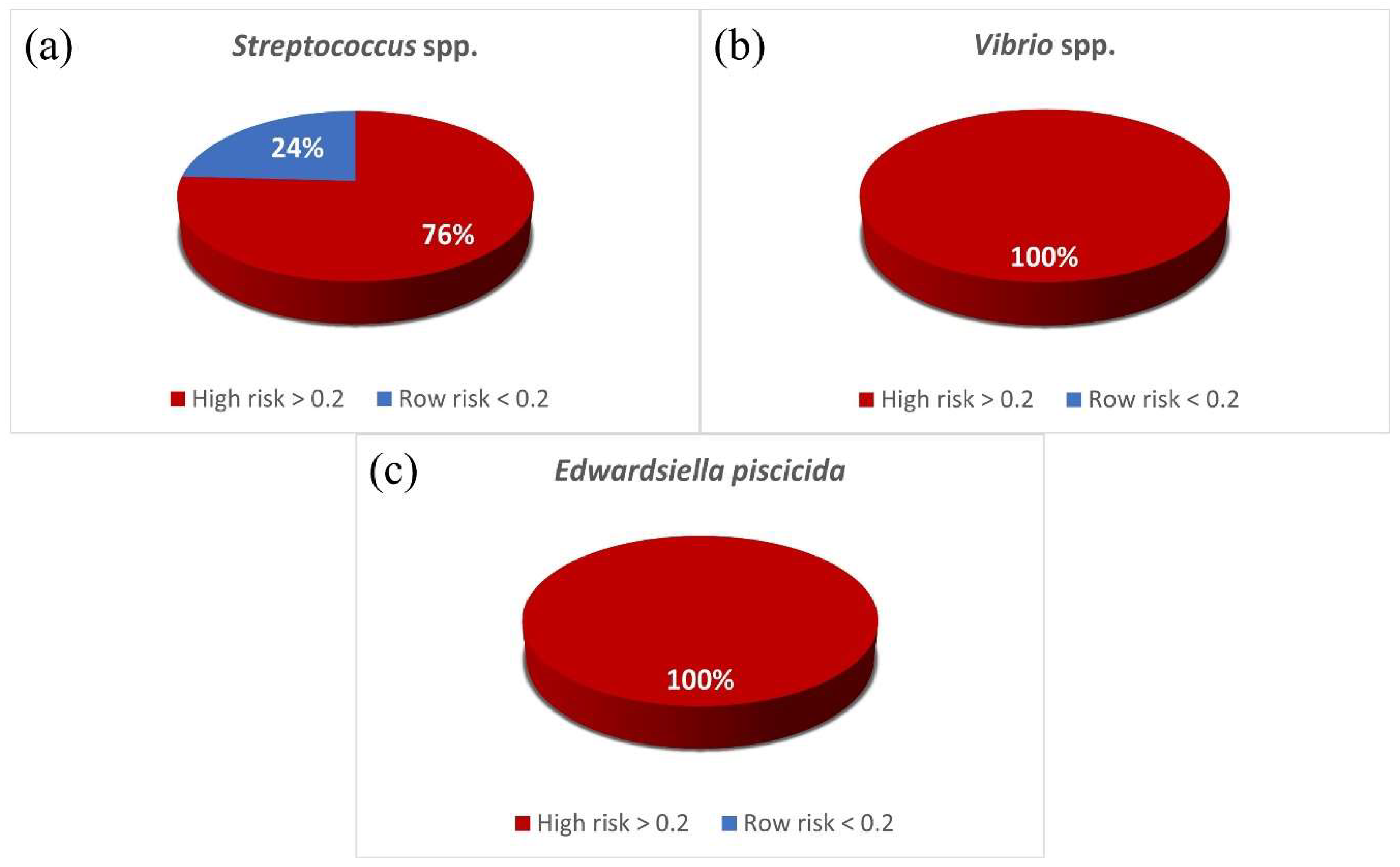
3.3. MAR Index and Antibiotic Resistance Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture. Meeting the Sustainable Developing Goals; FAO: Rome, Italy, 2020. [Google Scholar]
- KOSIS, Korean Statistical Information Service. Available online: http://kosis.kr (accessed on 5 April 2022).
- Jang, H.M.; Kim, Y.B.; Choi, S.; Lee, Y.; Shin, S.G.; Unno, T.; Kim, Y.M. Prevalence of antibiotic resistance genes from effluent of coastal aquaculture, South Korea. Environ. Pollut. 2018, 233, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in aquaculture. Sci. Rep. 2020, 10, 21878. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Cho, M.Y.; Park, G.H.; Won, K.M.; Choi, H.S.; Kim, M.S.; Park, M.A. Statistical data on infectious diseases of cultured olive flounder Paralichthys olivaceus from 2005 to 2007. J. Fish Pathol. 2010, 23, 369–377. [Google Scholar]
- Giraud, E.; Douet, D.G.; Le Bris, H.; Bouju-Albert, A.; Donnay-Moreno, C.; Thorin, C.; Pouliquen, H. Survey of antibiotic resistance in an integrated marine aquaculture system under oxolinic acid treatment. FEMS Microbiol. Ecol. 2006, 55, 439–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bris, H.; Dhaouadi, R.; Naviner, M.; Giraud, E.; Mangion, C.; Armand, F.; De La Cotte, N.; Thorin, C.; Ganière, J.-P.; Pouliquen, H. Experimental approach on the selection and persistence of anti-microbial-resistant Aeromonads in faecal matter of rainbow trout during and after an oxolinic acid treatment. Aquaculture 2007, 273, 416–422. [Google Scholar] [CrossRef]
- Guidi, L.R.; Santos, F.A.; Ribeiro, A.C.; Fernandes, C.; Silva, L.H.; Gloria, M.B. A simple, fast and sensitive screening LC-ESI-MS/MS method for antibiotics in fish. Talanta 2017, 163, 85–93. [Google Scholar] [CrossRef]
- Kim, Y.B.; Jeon, J.H.; Choi, S.; Shin, J.; Lee, Y.; Kim, Y.M. Use of a filtering process to remove solid waste and antibiotic resistance genes from effluent of a flow-through fish farm. Sci. Total Environ. 2018, 615, 289–296. [Google Scholar] [CrossRef]
- Jo, H.; Raza, S.; Farooq, A.; Kim, J.; Unno, T. Fish farm effluents as a source of antibiotic resistance gene dissemination on Jeju Island, South Korea. Environ. Pollut. 2021, 276, 116764. [Google Scholar] [CrossRef]
- Pepi, M.; Focardi, S. Antibiotic-resistant bacteria in aquaculture and climate change: A challenge for health in the mediterranean area. Int. J. Environ. Res. Public Health 2021, 18, 5723. [Google Scholar] [CrossRef]
- Walia, K.; Sharma, M.; Vijay, S.; Shome, B.R. Understanding policy dilemmas around antibiotic use in food animals & offering potential solutions. Indian J. Med. Res. 2019, 149, 107–118. [Google Scholar]
- Syal, K.; Mo, M.; Yu, H.; Iriya, R.; Jing, W.; Guodong, S.; Wang, S.; Grys, T.E.; Haydel, S.E.; Tao, N. Current and emerging techniques for antibiotic susceptibility tests. Theranostics 2017, 7, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, J.H.; Ferraro, M.J. Antimicrobial susceptibility testing: A review of general principles and contemporary practices. Clin. Infect. Dis. 2009, 49, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Ayandele, A.A.; Oladipo, E.K.; Oyebisi, O.; Kaka, M.O. Prevalence of multi-antibiotic resistant Escherichia coli and Klebsiella species obtained from a Tertiary Medical Institution in Oyo State, Nigeria. Qatar Med. J. 2020, 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.H.; Kim, H.J.; Lee, J.S.; Kim, J.W.; Park, S.I. Pathogenicity and classification of streptococci isolated from cultured marine fishesd. J. Fish Pathol. 2006, 19, 17–33. [Google Scholar]
- Griffin, M.J.; Ware, C.; Quiniou, S.M.; Steadman, J.M.; Gaunt, P.S.; Khoo, L.H.; Soto, E. Edwardsiella piscicida identified in the Southeastern USA by gyrB sequence, species-specific and repetitive sequence-mediated PCR. Dis. Aquat. Organ. 2014, 108, 23–35. [Google Scholar] [CrossRef]
- Kim, H.J.; Ryu, J.O.; Lee, S.Y.; Kim, E.S.; Kim, H.Y. Multiplex PCR for detection of the Vibrio genus and five pathogenic Vibrio species with primer sets designed using comparative genomics. BMC Microbiol. 2015, 15, 239. [Google Scholar] [CrossRef] [Green Version]
- Demircan, D.; Candan, A. Identification of Vibrio anguillarum by PCR (rpoN gene) associated with vibriosis in marine fish in Turkey. Turk. J. Vet. Anim. Sci. 2006, 30, 305–310. [Google Scholar]
- Kim, M.S.; Cho, J.Y.; Choi, H.S. Identification of Vibrio harveyi, Vibrio ichthyoenteri, and Photobacterium damselae isolated from olive flounder Paralichthys olivaceus in Korea by multiplex PCR developed using the rpoB gene. Fish. Sci. 2014, 80, 333–339. [Google Scholar] [CrossRef]
- Kim, Y.J.; Jun, L.J.; Kang, M.R.; Lee, D.W.; Woo, S.J.; Kim, M.S.; Jeong, J.B. Establishing of optimal culture conditions for MIC panels for MIC determination of fish bacterial pathogens. Korean J. Fish. Aquat. Sci. 2020, 53, 443–450. [Google Scholar]
- CLSI. Methods for Antimicrobial Disk Susceptibility Testing of Bacteria Isolated from Aquatic Animals: Approved Guideline; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2006. [Google Scholar]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Nho, S.W.; Shin, G.W.; Park, S.B.; Jang, H.B.; Cha, I.S.; Ha, M.A.; Kim, Y.R.; Park, Y.K.; Dalvi, R.S.; Kang, B.J.; et al. Phenotypic characteristics of Streptococcus iniae and Streptococcus parauberis isolated from olive flounder (Paralichthys olivaceus). FEMS Microbiol. Lett. 2009, 293, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarr, C.L.; Patel, J.S.; Puhr, N.D.; Sowers, E.G.; Bopp, C.A.; Strockbine, N.A. Identification of Vibrio isolates by a multiplex PCR assay and rpoB sequence determination. J. Clin. Microbiol. 2007, 45, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.B.; Aoki, T.; Jung, T.S. Pathogenesis of and strategies for preventing Edwardsiella tarda infection in fish. Vet. Res. 2012, 43, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Gong, C.; Gu, H.; Huang, H.; Xian, J.; Hu, Y. Bicistronic operon YhaO-YhaM contributes to antibiotic resistance and virulence of pathogen Edwardsiella piscicida. Aquaculture 2021, 541, 736849. [Google Scholar] [CrossRef]
- Grover, G.; Kini, S.G. Synthesis and evaluation of new quinazolone derivatives of nalidixic acid as potential antibacterial and antifungal agents. Eur. J. Med. Chem. 2006, 41, 256–262. [Google Scholar] [CrossRef]
- Cho, M.Y.; Oh, Y.K.; Lee, D.C.; Kim, J.H.; Park, M.A. Geographical comparison on different methods for identification of Streptococcus parauberis isolated from cultured olive flounder, Paralichthys olivaceus. J. Fish Pathol. 2007, 20, 49–60. [Google Scholar]
- Boby, N.; Abbas, M.A.; Lee, E.B.; Park, S.C. Pharmacodynamics of ceftiofur selected by genomic and proteomic approaches of Streptococcus parauberis isolated from the flounder, Paralichthys olivaceus. Int. J. Genom. 2020, 2020, 4850290. [Google Scholar] [CrossRef]
- Park, J.Y.; Birhanu, B.T.; Lee, S.J.; Park, N.H.; Kim, J.Y.; Mechesso, A.F.; Boby, N.; Park, S.C. Pharmacodynamics of amoxicillin against field isolates of Streptococcus parauberis from olive flounder (Paralichthys olivaceus). Aquac. Res. 2018, 49, 1060–1071. [Google Scholar] [CrossRef]
- Laganà, P.; Caruso, G.; Minutoli, E.; Zaccone, R.; Delia, S. Susceptibility to antibiotics of Vibrio spp. and Photobacterium damsela ssp. piscicida strains isolated from Italian aquaculture farms. New Microbiol. 2011, 34, 53–63. [Google Scholar]
- Raissy, M.; Moumeni, M.; Ansari, M.; Rahimi, E. Antibiotic resistance pattern of some Vibrio strains isolated from seafood. Iran. J. Fish. Sci. 2012, 3, 618–626. [Google Scholar]
- Letchumanan, V.; Pusparajah, P.; Tan, L.T.; Yin, W.F.; Lee, L.H.; Chan, K.G. Occurrence and antibiotic resistance of Vibrio parahaemolyticus from shellfish in Selangor, Malaysia. Front. Microbiol. 2015, 6, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanetti, S.; Spanu, T.; Deriu, A.; Romano, L.; Sechi, L.A.; Fadda, G. In vitro susceptibility of Vibrio spp. isolated from the environment. Int. J. Antimicrob. Agents 2001, 17, 407–409. [Google Scholar] [CrossRef]
- Petković, H.; Lukežič, T.; Šušković, J. Biosynthesis of oxytetracycline by Streptomyces rimosus: Past, present and future directions in the development of tetracycline antibiotics. Food Technol. Biotechnol. 2017, 55, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Stock, I.; Wiedemann, B. Natural antibiotic susceptibilities of Edwardsiella tarda, E. ictaluri, and E. hoshinae. Antimicrob. Agents Chemother. 2001, 45, 2245–2255. [Google Scholar] [CrossRef] [Green Version]
- Niu, G.; Wongsathein, D.; Boonyayatra, S.; Khattiya, R. Occurrence of multiple antibiotic resistance and genotypic characterization in Edwardsiella tarda isolated from cage-cultured hybrid red tilapia (Oreochromis sp.) in the Ping River, Northern Thailand. Aquac. Res. 2019, 50, 3643–3652. [Google Scholar] [CrossRef]
- Niestepski, S.; Harnisz, M.; Korzeniewska, E.; Osinska, A. Markers specific to Bacteroides fragilis group bacteria as indicators of anthropogenic pollution of surface waters. Int. J. Environ. Res. Public Health 2020, 17, 7137. [Google Scholar] [CrossRef]
- Almeida, A.R.; Tacao, M.; Soares, J.; Domingues, I.; Henriques, I. Tetracycline-resistant bacteria selected from water and zebrafish after antibiotic exposure. Int. J. Environ. Res. Public Health 2021, 18, 3218. [Google Scholar] [CrossRef]
- Cho, K.T.; Kim, Y.C.; Kwon, W.J.; Jeong, H.D. Characterization of Tetracycline-Resistant Genes of Staphylococci isolates from Human and Marine Environment. J. Fish. Mar. Sci. Educ. 2016, 28, 59–68. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing of Bacteria Isolated from Aquatic Animals; Second Information Supplement; CLSI: Wayne, PA, USA, 2014. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Pathogen | Oligonucleotide Sequences | Product Size (bp) | Annealing Temp. (°C) | Cycles | Reference |
|---|---|---|---|---|---|
| Streptococcus iniae | 5′-AAGAGACGCAGTGTCAAAAG CGTTTCTTATCTTGTTACTC-3′ | 107 | 55 | 33 | [16] |
| S. parauberis | 5′-TCCAGTCTTTCGACCTTCTT CAAAGAGATGTTCGGCTTG-3′ | 220 | |||
| Edwardsiella tarda | 5′-CAGTGATAAAAAGGGGTGGA CTACACAGCAACGACAACG-3′ | 114 | 58 | 30 | [17] |
| E. piscicida | 5′-CTTTGATCATGGTTGCGGAA CGGCGTTTTCTTTTCTCG-3′ | 130 | |||
| Vibrio genus | 5′-GTCARATTGAAAARCARTTYGGTAAAGG ACYTTRATRCGNGTTTCRTTRCC-3′ | 689 | 60 | 30 | [18] |
| V. alginolyticus | 5′-ACGGCATTGGAAATTGCGACTG TACCCGTCTCACGAGCCCAAG-3′ | 199 | |||
| V. parahaemolyticus | 5′-AGCTTATTGGCGGTTTCTGTCGG CKCAAGACCAAGAAAAGCCGTC-3′ | 297 | |||
| V. anguillarum | 5′-GTTCATAGCATCAATGAGGAG GAGCAGACAATATGTTGGATG-3′ | 519 | [19] | ||
| V. harveyi | 5′-GTGATGAAGAAGCTTATCGCGATT CGCCTTCTTCAGTTAACGCAGGA-3′ | 601 | [20] | ||
| V. ichthyoenteri/V. scophthalmi | 5′-ATGCAATCATGCCTCAAGATCTA AAATGTACCTTCTTCAGTCAACTT-3′ | 434 | |||
| Photobacterium damselae | 5′-CAAGACATCATCGATGTGATGCGT GAAACTTTACCATCTACCACTTTG-3′ | 533 |
| Species | Year | No. of Isolates |
|---|---|---|
| Streptococcus parauberis | 2019 | 28 |
| S. iniae | 2015 | 1 |
| Vibrio harveyi | 2013–2017 | 19 |
| Photobacterium damselae | 2016–2020 | 3 |
| V. alginolyticus | 2018–2020 | 3 |
| V. anguillarum | 2013 | 1 |
| V. scophthalmi | 2015 | 1 |
| V. parahaemolyticus | 2020 | 1 |
| Edwardsiella piscicida | 2010 | 4 |
| E. piscicida | 2013 | 1 |
| E. piscicida | 2014 | 2 |
| E. piscicida | 2015 | 3 |
| E. piscicida | 2019 | 16 |
| E. piscicida | 2020 | 1 |
| Total | 84 |
| No. | Antibiotic Resistant Pattern | MAR Index (No. of Antibiotics Resistance) | % of Isolates | ||
|---|---|---|---|---|---|
| S | V | E | |||
| 1 | NAL | 0.11 (1) | 24.14 | 0 | 0 |
| 2 | ERY/NAL | 0.22 (2) | 3.45 | 0 | 0 |
| 3 | GEN/NAL | 24.14 | 0 | 0 | |
| 4 | DXT/TET/NAL | 0.33 (3) | 3.45 | 0 | 0 |
| 5 | ERY/TET/NAL | 3.45 | 0 | 0 | |
| 6 | CLI/ERY/NAL | 3.45 | 0 | 14.81 | |
| 7 | GEN/NAL/ENRO | 6.90 | 0 | 0 | |
| 8 | GEN/ERY/NAL | 3.45 | 0 | 0 | |
| 9 | AMP/CLI/ERY | 0 | 28.57 | 0 | |
| 10 | ERY/DXT/TET/NAL | 0.44 (4) | 3.45 | 0 | 0 |
| 11 | CLI/ERY/TET/NAL | 10.34 | 0 | 0 | |
| 12 | CLI/ERY/DXT/NAL | 3.45 | 0 | 0 | |
| 13 | GEN/ERY/NAL/ENRO | 3.45 | 0 | 0 | |
| 14 | CLI/ERY/DXT/TET | 0 | 3.57 | 0 | |
| 15 | AMP/CLI/ERY/ENRO | 0 | 7.14 | 0 | |
| 16 | AMP/CLI/ERY/TET | 0 | 17.86 | 0 | |
| 17 | CLI/ERY/TET/NAL | 0 | 0 | 3.70 | |
| 18 | AMP/CLI/ERY/ENR | 0 | 0 | 3.70 | |
| 19 | AMP/ERY/TET/NAL/ENR | 0.56 (5) | 3.45 | 0 | 0 |
| 20 | AMP/CLI/ERY/NAL/ENR | 0 | 10.71 | 0 | |
| 21 | AMP/CLI/ERY/TET/NAL | 0 | 3.57 | 0 | |
| 22 | AMP/CLI/ERY/DXT/TET | 0 | 10.71 | 0 | |
| 23 | AMP/GEN/CLI/ERY/ENRO | 0 | 3.57 | 0 | |
| 24 | CLI/ERY/DXT/TET/NAL | 0 | 0 | 3.70 | |
| 25 | GEN/ERY/TET/NAL/ENRO | 0 | 0 | 3.70 | |
| 26 | AMP/CLI/ERY/NAL/ENRO | 0 | 0 | 11.11 | |
| 27 | GEN/CLI/ERY/DXT/TET/NAL | 0.67 (6) | 3.45 | 0 | 0 |
| 28 | AMP/CLI/ERY/DXT/TET/ENRO | 0 | 7.14 | 0 | |
| 29 | CLI/ERY/DXT/TET/NAL/ENRO | 0 | 0 | 3.70 | |
| 30 | AMP/CLI/ERY/TET/NAL/ENRO | 0 | 0 | 3.70 | |
| 31 | AMP/GEN/CLI/ERY/NAL/ENRO | 0 | 0 | 3.70 | |
| 32 | AMP/GEN/CLI/ERY/DXT/TET/ENRO | 0.78 (7) | 0 | 3.57 | 0 |
| 33 | AMP/CLI/ERY/DXT/TET/NAL/ENRO | 0 | 0 | 44.44 | |
| 34 | AMP/GEN/CLI/ERY/DXT/TET/NAL/ENRO | 0.89 (8) | 0 | 3.57 | 3.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Jun, L.J.; Lee, D.W.; Lee, Y.J.; Ko, Y.J.; Oh, Y.E.; Woo, S.J.; Kim, M.S.; Kim, S.M.; Jeong, J.B. Antibiotic Susceptibility of Bacterial Pathogens That Infect Olive Flounder (Paralichthys olivaceus) Cultivated in Korea. Int. J. Environ. Res. Public Health 2022, 19, 8110. https://doi.org/10.3390/ijerph19138110
Kim YJ, Jun LJ, Lee DW, Lee YJ, Ko YJ, Oh YE, Woo SJ, Kim MS, Kim SM, Jeong JB. Antibiotic Susceptibility of Bacterial Pathogens That Infect Olive Flounder (Paralichthys olivaceus) Cultivated in Korea. International Journal of Environmental Research and Public Health. 2022; 19(13):8110. https://doi.org/10.3390/ijerph19138110
Chicago/Turabian StyleKim, Ye Ji, Lyu Jin Jun, Da Won Lee, Young Juhn Lee, Ye Jin Ko, Yeong Eun Oh, Soo Ji Woo, Myoung Sug Kim, Seung Min Kim, and Joon Bum Jeong. 2022. "Antibiotic Susceptibility of Bacterial Pathogens That Infect Olive Flounder (Paralichthys olivaceus) Cultivated in Korea" International Journal of Environmental Research and Public Health 19, no. 13: 8110. https://doi.org/10.3390/ijerph19138110