Characterisation of Microbial Community Associated with Different Disinfection Treatments in Hospital hot Water Networks

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Water Distribution Systems
2.2. Samples Collection
2.3. Characterization of Microbial Community
Bacterial Community 16S Profiling
2.4. Data Analysis
3. Results
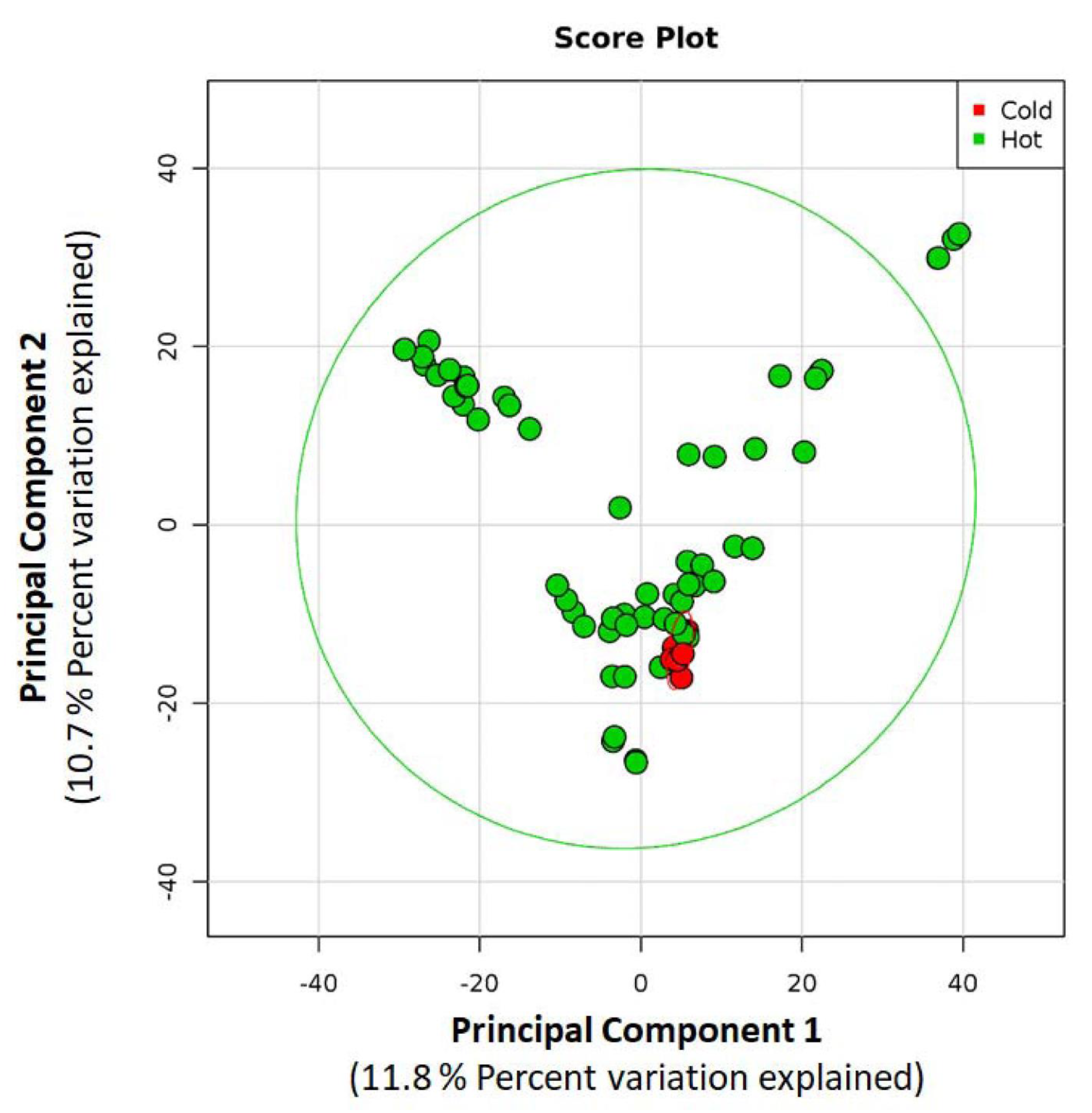
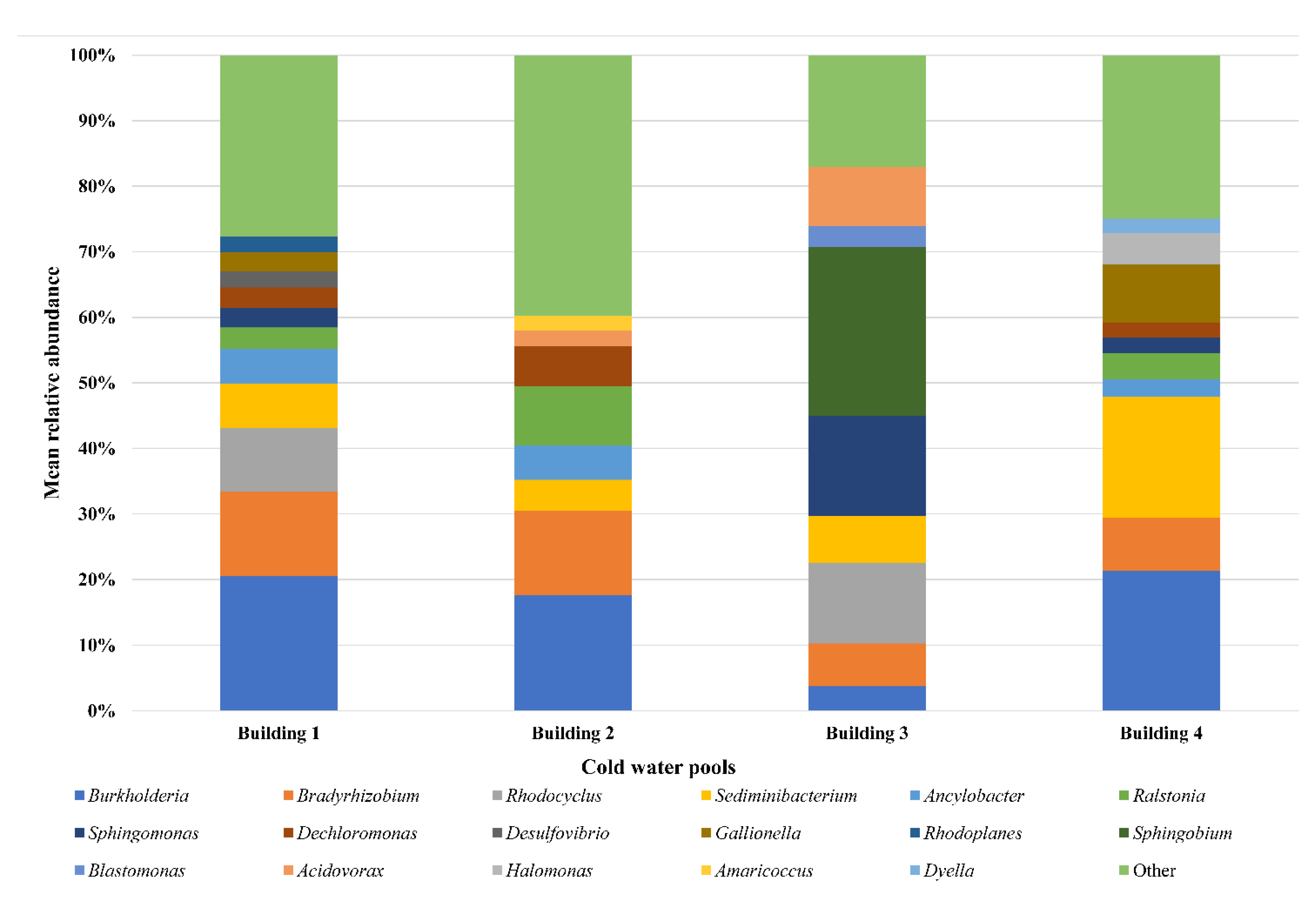
3.1. Cold Water Samples
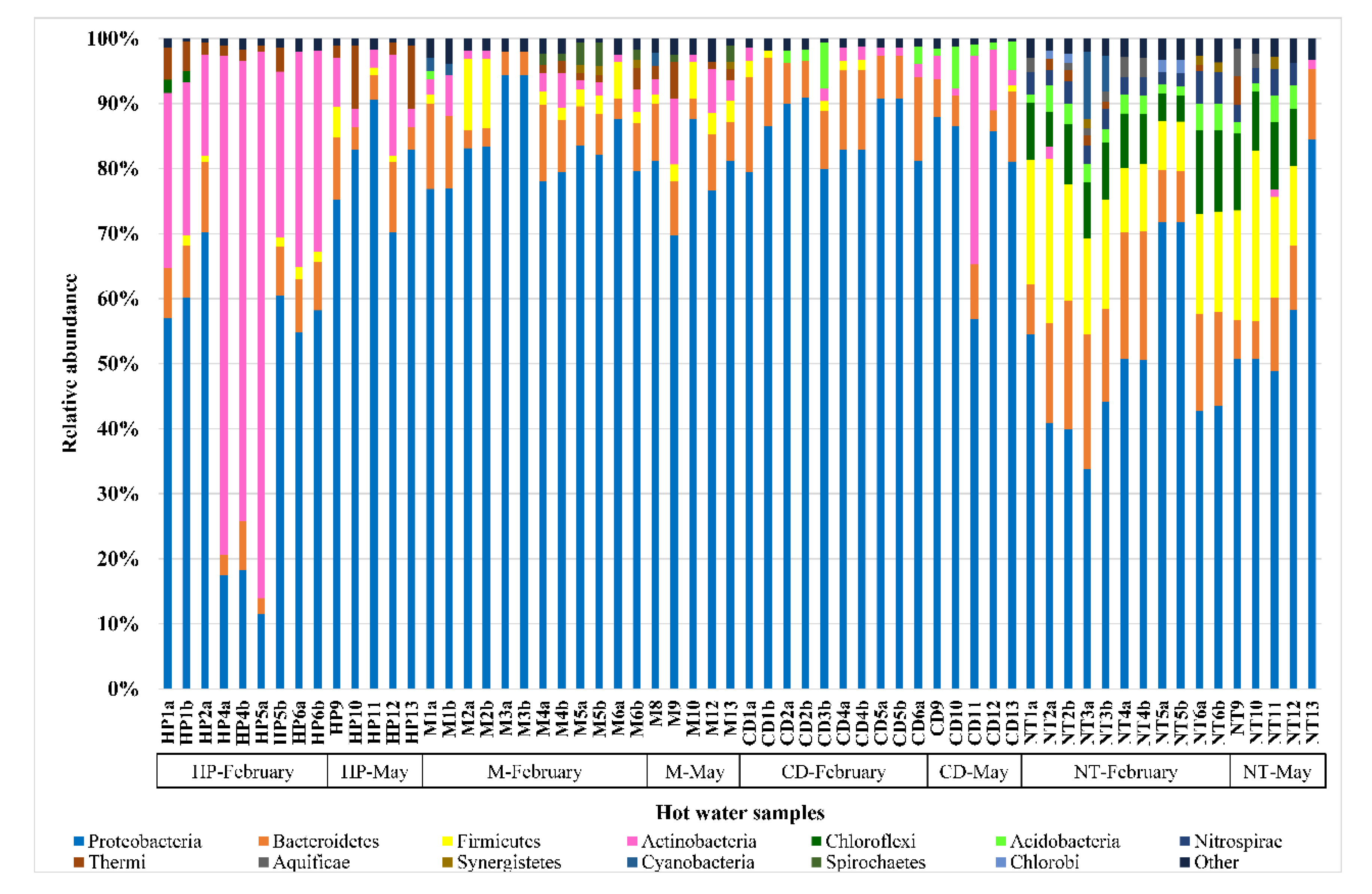
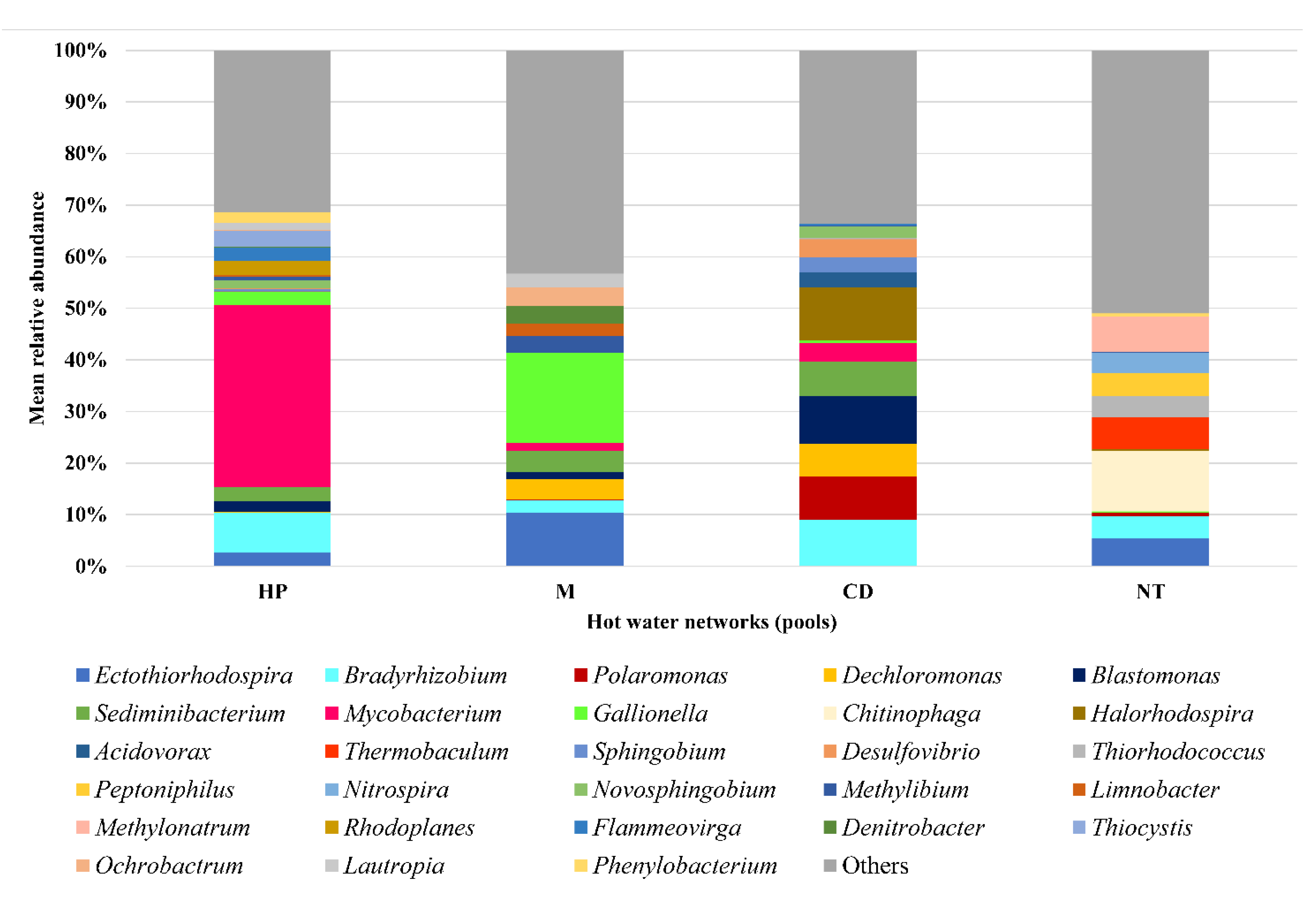
3.2. Hot Water Samples
3.3. Genera Containing Opportunistic Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Decker, B.K.; Palmore, T.N. Hospital water and opportunities for infection prevention. Curr. Infect. Dis. Rep. 2014, 16, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargellini, A.; Marchesi, I.; Righi, E.; Ferrari, A.; Cencetti, S.; Borella, P.; Rovesti, S. Parameters predictive of Legionella contamination in hot water systems: Association with trace elements and heterotrophic plate counts. Water Res. 2011, 45, 2315–2321. [Google Scholar] [CrossRef] [PubMed]
- Giovanardi, R.; Bononi, M.; Messori, M.; Bargellini, A.; Paduano, S.; Borella, P.; Marchesi, I. Corrosion resistance of commonly used plumbing materials for water distribution systems exposed to disinfection treatments. Corros. Eng. Sci. Technol. 2020, 1–8. [Google Scholar] [CrossRef]
- Wang, H.; Masters, S.; Edwards, M.A.; Falkinham, J.O.; Pruden, A. Effect of disinfectant, water age, and pipe materials on bacterial and eukaryotic community structure in drinking water biofilm. Environ. Sci. Technol. 2014, 48, 1426–1435. [Google Scholar] [CrossRef]
- Berry, D.; Xi, C.; Raskin, L. Microbial ecology of drinking water distribution systems. Curr. Opin. Biotechnol. 2006, 17, 297–302. [Google Scholar] [CrossRef]
- Anaissie, E.J.; Penzak, S.R.; Dignani, M.C. The hospital water supply as a source of nosocomial infections: A plea for action. Arch. Intern. Med. 2002, 162, 1483–1492. [Google Scholar] [CrossRef] [Green Version]
- Cervia, J.S.; Ortolano, G.A.; Canonica, F.P. Hospital tap water as a source of Stenotrophomonas maltophilia infection. Clin. Infect. Dis. 2008, 46, 1485–1487. [Google Scholar] [CrossRef]
- Falkinham, J.O., 3rd. Common features of opportunistic premise plumbing pathogens. Int. J. Environ. Res. Public Health 2015, 12, 4533–4545. [Google Scholar] [CrossRef]
- Kusnetsov, J.; Torvinen, E.; Perola, O.; Nousiainen, T.; Katila, M.L. Colonization of hospital water systems by legionellae, mycobacteria and other heterotrophic bacteria potentially hazardous to risk group patients. APMIS 2003, 111, 546–556. [Google Scholar] [CrossRef]
- Merlani, G.M.; Francioli, P. Established and emerging waterborne nosocomial infections. Curr. Opin. Infect. Dis. 2003, 16, 343–347. [Google Scholar] [CrossRef]
- Wang, J.L.; Chen, M.L.; Lin, Y.E.; Chang, S.C.; Chen, Y.C. Association between contaminated faucets and colonization or infection by nonfermenting gram-negative bacteria in intensive care units in Taiwan. J. Clin. Microbiol. 2009, 47, 3226–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.E.; Stout, J.E.; Yu, V.L. Controlling Legionella in Hospital Drinking Water: An Evidence-Based Review of Disinfection Methods. Infect. Control Hosp. Epidemiol. 2011, 32, 166–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, I.; Ferranti, G.; Mansi, A.; Marcelloni, A.M.; Proietto, A.R.; Saini, N.; Borella, P.; Bargellini, A. Control of Legionella Contamination and Risk of Corrosion in Hospital Water Networks following Various Disinfection Procedures. Appl. Environ. Microbiol. 2016, 82, 2959–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, I.; Marchegiano, P.; Bargellini, A.; Cencetti, S.; Frezza, G.; Miselli, M.; Borella, P. Effectiveness of different methods to control legionella in the water supply: Ten-year experience in an Italian university hospital. J. Hosp. Infect. 2011, 77, 47–51. [Google Scholar] [CrossRef] [PubMed]
- The European Centre for Disease Prevention and Control (ECDC). European Technical Guidelines for the Prevention, Control and Investigation of Infections Caused by Legionella Species; ECDC: Solna, Sweden, 2017. [Google Scholar]
- Darelid, J.; Lofgren, S.; Malmvall, B.E. Control of nosocomial Legionnaires’ disease by keeping the circulating hot water temperature above 55 degrees C: Experience from a 10-year surveillance programme in a district general hospital. J. Hosp. Infect. 2002, 50, 213–219. [Google Scholar] [CrossRef]
- Borella, P.; Bargellini, A.; Marchegiano, P.; Vecchi, E.; Marchesi, I. Hospital-acquired Legionella infections: An update on the procedures for controlling environmental contamination. Ann. Ig. 2016, 28, 98–108. [Google Scholar] [CrossRef]
- Coniglio, M.A.; Ferrante, M.; Yassin, M.H. Preventing Healthcare-Associated Legionellosis: Results after 3 Years of Continuous Disinfection of Hot Water with Monochloramine and an Effective Water Safety Plan. Int. J. Environ. Res. Public Health 2018, 15, 1594. [Google Scholar] [CrossRef] [Green Version]
- Casini, B.; Buzzigoli, A.; Cristina, M.L.; Spagnolo, A.M.; Del Giudice, P.; Brusaferro, S.; Poscia, A.; Moscato, U.; Valentini, P.; Baggiani, A.; et al. Long-term effects of hospital water network disinfection on Legionella and other waterborne bacteria in an Italian university hospital. Infect. Control Hosp. Epidemiol. 2014, 35, 293–299. [Google Scholar] [CrossRef]
- Hua, G.; Reckhow, D.A. Comparison of disinfection byproduct formation from chlorine and alternative disinfectants. Water Res. 2007, 41, 1667–1678. [Google Scholar] [CrossRef]
- Rhoads, W.J.; Pruden, A.; Edwards, M.A. Interactive Effects of Corrosion, Copper, and Chloramines on Legionella and Mycobacteria in Hot Water Plumbing. Environ. Sci. Technol. 2017, 51, 7065–7075. [Google Scholar] [CrossRef]
- Abdel-Nour, M.; Duncan, C.; Low, D.E.; Guyard, C. Biofilms: The stronghold of Legionella pneumophila. Int. J. Mol. Sci. 2013, 14, 21660–21675. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.H.; Pressman, J.G.; Wahman, D.G. Three-Dimensional Free Chlorine and Monochloramine Biofilm Penetration: Correlating Penetration with Biofilm Activity and Viability. Environ. Sci. Technol. 2018, 52, 1889–1898. [Google Scholar] [CrossRef]
- Badhai, J.; Ghosh, T.S.; Das, S.K. Taxonomic and functional characteristics of microbial communities and their correlation with physicochemical properties of four geothermal springs in Odisha, India. Front. Microbiol. 2015, 6, 1166. [Google Scholar] [CrossRef] [PubMed]
- Paduano, S.; Valeriani, F.; Romano-Spica, V.; Bargellini, A.; Borella, P.; Marchesi, I. Microbial biodiversity of thermal water and mud in an Italian spa by metagenomics: A pilot study. Water Sci. Technol. Water Supply 2018, 18, 1456–1465. [Google Scholar] [CrossRef]
- Amin, A.; Ahmed, I.; Salam, N.; Kim, B.-Y.; Singh, D.; Zhi, X.-Y.; Xiao, M.; Li, W.-J. Diversity and Distribution of Thermophilic Bacteria in Hot Springs of Pakistan. Microb. Ecol. 2017, 74, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Bautista-de Los Santos, Q.M.; Schroeder, J.L.; Blakemore, O.; Moses, J.; Haffey, M.; Sloan, W.; Pinto, A.J. The impact of sampling, PCR, and sequencing replication on discerning changes in drinking water bacterial community over diurnal time-scales. Water Res. 2016, 90, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Zhang, J.; Xu, W.; Mou, Z. Microbial Community Structure in the Sediments and Its Relation to Environmental Factors in Eutrophicated Sancha Lake. Int. J. Environ. Res. Public Health 2019, 16, 1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, C.R.; Hammes, F. Drinking water microbiology--from measurement to management. Curr. Opin. Biotechnol. 2015, 33, 87–94. [Google Scholar] [CrossRef]
- Valeriani, F.; Crognale, S.; Protano, C.; Gianfranceschi, G.; Orsini, M.; Vitali, M.; Spica, V.R. Metagenomic analysis of bacterial community in a travertine depositing hot spring. New Microbiol. 2018, 41, 126–135. [Google Scholar]
- Potgieter, S.; Pinto, A.; Sigudu, M.; du Preez, H.; Ncube, E.; Venter, S. Long-term spatial and temporal microbial community dynamics in a large-scale drinking water distribution system with multiple disinfectant regimes. Water Res. 2018, 139, 406–419. [Google Scholar] [CrossRef] [Green Version]
- Sidari, F.P.; Stout, J.E.; Vanbriesen, J.M.; Bowman, A.M.; Grubb, D.; Neuner, A.; Wagener, M.M.; Yu, V.L. Keeping Legionella out of water systems. J. Am. Water Works Assoc. 2004, 96, 111–119. [Google Scholar] [CrossRef]
- Kittelmann, S.; Seedorf, H.; Walters, W.A.; Clemente, J.C.; Knight, R.; Gordon, J.I.; Janssen, P.H. Simultaneous amplicon sequencing to explore co-occurrence patterns of bacterial, archaeal and eukaryotic microorganisms in rumen microbial communities. PLoS ONE 2013, 8, e47879. [Google Scholar] [CrossRef] [PubMed]
- Valeriani, F.; Protano, C.; Gianfranceschi, G.; Leoni, E.; Galasso, V.; Mucci, N.; Vitali, M.; Spica, V.R. Microflora Thermarum Atlas project: Biodiversity in thermal spring waters and natural SPA pools. Water Sci. Technol. Water Supply 2018, 18, 1472–1483. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors—Occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef]
- Van Aken, B.; Lin, L.S. Effect of the disinfection agents chlorine, UV irradiation, silver ions, and TiO2 nanoparticles/near-UV on DNA molecules. Water Sci. Technol. 2011, 64, 1226–1232. [Google Scholar] [CrossRef]
- Kong, Y. Btrim: A fast, lightweight adapter and quality trimming program for next-generation sequencing technologies. Genomics 2011, 98, 152–153. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Hou, W.; Dong, H.; Jiang, H.; Huang, L.; Wu, G.; Zhang, C.; Song, Z.; Zhang, Y.; Ren, H.; et al. Control of temperature on microbial community structure in hot springs of the Tibetan Plateau. PLoS ONE 2013, 8, e62901. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.J.; Baetens, J.M.; De Baets, B. Ecological Diversity: Measuring the Unmeasurable. Mathematics 2018, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.-J. Abundance-based similarity indices and their estimation when there are unseen species in samples. Biometrics 2006, 62, 361–371. [Google Scholar] [CrossRef]
- Horn, H.S. Measurement of "Overlap" in Comparative Ecological Studies. Am. Nat. 1966, 100, 419–424. [Google Scholar] [CrossRef]
- Wolda, H. Similarity indices, sample size and diversity. Oecologia 1981, 50, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Xia, J.; Liu, Y.; Zhou, Y.; Guo, A.C.; Cruz, J.A.; Sinelnikov, I.; Budwill, K.; Nesbø, C.L.; Wishart, D.S. METAGENassist: A comprehensive web server for comparative metagenomics. Nucleic Acids Res. 2012, 40, W88–W95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The two-way layout with no replication. Mar. Biol. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Marchesi, I.; Ferranti, G.; Bargellini, A.; Marchegiano, P.; Predieri, G.; Stout, J.E.; Borella, P. Monochloramine and chlorine dioxide for controlling Legionella pneumophila contamination: Biocide levels and disinfection by-product formation in hospital water networks. J. Water Health 2013, 11, 738–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Alvarez, V.; Revetta, R.P.; Santo Domingo, J.W. Metagenomic analyses of drinking water receiving different disinfection treatments. Appl. Environ. Microbiol. 2012, 78, 6095–6102. [Google Scholar] [CrossRef] [Green Version]
- Van Assche, A.; Crauwels, S.; De Brabanter, J.; Willems, K.A.; Lievens, B. Characterization of the bacterial community composition in water of drinking water production and distribution systems in Flanders, Belgium. MicrobiologyOpen 2018, 8, e726. [Google Scholar] [CrossRef]
- Botero, L.M.; Brown, K.B.; Brumefield, S.; Burr, M.; Castenholz, R.W.; Young, M.; McDermott, T.R. Thermobaculum terrenum gen. nov., sp. nov.: A non-phototrophic gram-positive thermophile representing an environmental clone group related to the Chloroflexi (green non-sulfur bacteria) and Thermomicrobia. Arch. Microbiol. 2004, 181, 269–277. [Google Scholar] [CrossRef]
- Buse, H.Y.; Ji, P.; Gomez-Alvarez, V.; Pruden, A.; Edwards, M.A.; Ashbolt, N.J. Effect of temperature and colonization of Legionella pneumophila and Vermamoeba vermiformis on bacterial community composition of copper drinking water biofilms. Microb. Biotechnol. 2017, 10, 773–788. [Google Scholar] [CrossRef]
- Berrada, H.; Fikri-Benbrahim, K. Taxonomy of the Rhizobia: Current Perspectives. Br. Microbiol. Res. J. 2014, 4, 616–639. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Alvarez, V.; Schrantz, K.A.; Pressman, J.G.; Wahman, D.G. Biofilm community dynamics in bench-scale annular reactors simulating arrestment of chloraminated drinking water nitrification. Environ. Sci. Technol. 2014, 48, 5448–5457. [Google Scholar] [CrossRef]
- September, S.M.; Brözel, V.S.; Venter, S.N. Diversity of nontuberculoid Mycobacterium species in biofilms of urban and semiurban drinking water distribution systems. Appl. Environ. Microbiol. 2004, 70, 7571–7573. [Google Scholar] [CrossRef] [Green Version]
- Baron, J.L.; Vikram, A.; Duda, S.; Stout, J.E.; Bibby, K. Shift in the microbial ecology of a hospital hot water system following the introduction of an on-site monochloramine disinfection system. PLoS ONE 2014, 9, e102679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.M.; Santo Domingo, J.W.; Meckes, M.C. Population diversity in model potable water biofilms receiving chlorine or chloramine residual. Biofouling 2005, 21, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Revi. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Faria, S.; Joao, I.; Jordao, L. General Overview on Nontuberculous Mycobacteria, Biofilms, and Human Infection. J. Pathog. 2015, 2015, 809014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, I.W. The biofilm matrix--an immobilized but dynamic microbial environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Ojha, A.; Anand, M.; Bhatt, A.; Kremer, L.; Jacobs, W.R., Jr.; Hatfull, G.F. GroEL1: A dedicated chaperone involved in mycolic acid biosynthesis during biofilm formation in mycobacteria. Cell 2005, 123, 861–873. [Google Scholar] [CrossRef] [Green Version]
- Ojha, A.K.; Baughn, A.D.; Sambandan, D.; Hsu, T.; Trivelli, X.; Guerardel, Y.; Alahari, A.; Kremer, L.; Jacobs, W.R., Jr.; Hatfull, G.F. Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria. Mol. Microbiol. 2008, 69, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Li, Z.; Yu, J.; Cao, N.; Liu, R.; Yang, M. Characterization of bacterial community structure in a drinking water distribution system during an occurrence of red water. Appl. Environ. Microbiol. 2010, 76, 7171–7180. [Google Scholar] [CrossRef] [Green Version]
- Ridgway, H.F.; Means, E.G.; Olson, B.H. Iron bacteria in drinking-water distribution systems: Elemental analysis of Gallionella stalks, using x-ray energy-dispersive microanalysis. Appl. Environ. Microbiol. 1981, 41, 288–297. [Google Scholar] [CrossRef] [Green Version]
- Kaestli, M.; O’Donnell, M.; Rose, A.; Webb, J.R.; Mayo, M.; Currie, B.J.; Gibb, K. Opportunistic pathogens and large microbial diversity detected in source-to-distribution drinking water of three remote communities in Northern Australia. PLoS Negl. Trop. Dis. 2019, 13, e0007672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Tominski, C.; Kappler, A.; Behrens, S.; Roden, E.E. Metagenomic Analyses of the Autotrophic Fe(II)-Oxidizing, Nitrate-Reducing Enrichment Culture KS. Appl. Environ. Microbiol. 2016, 82, 2656–2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-K.; Chu, Z.-R.; Liu, Y.-J.; Zhu, M.-T.; Yang, L.; Zhang, J. Molecular characterization of microbial populations in full-scale biofilters treating iron, manganese and ammonia containing groundwater in Harbin, China. Bioresour. Technol. 2013, 147, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Stanish, L.F.; Hull, N.M.; Robertson, C.E.; Harris, J.K.; Stevens, M.J.; Spear, J.R.; Pace, N.R. Factors Influencing Bacterial Diversity and Community Composition in Municipal Drinking Waters in the Ohio River Basin, USA. PLoS ONE 2016, 11, e0157966. [Google Scholar] [CrossRef]
- Pressman, J.G.; Lee, W.H.; Bishop, P.L.; Wahman, D.G. Effect of free ammonia concentration on monochloramine penetration within a nitrifying biofilm and its effect on activity, viability, and recovery. Water Res. 2012, 46, 882–894. [Google Scholar] [CrossRef]
- Buffet-Bataillon, S.; Bonnaure-Mallet, M.; de la Pintiere, A.; Defawe, G.; Gautier-Lerestif, A.L.; Fauveau, S.; Minet, J. Heterotrophic bacterial growth on hoses in a neonatal water distribution system. J. Microbiol. Biotechnol. 2010, 20, 779–781. [Google Scholar]
- Liu, R.; Yu, Z.; Guo, H.; Liu, M.; Zhang, H.; Yang, M. Pyrosequencing analysis of eukaryotic and bacterial communities in faucet biofilms. Sci. Total Environ. 2012, 435–436, 124–131. [Google Scholar] [CrossRef]
- Soto-Giron, M.J.; Rodriguez-R, L.M.; Luo, C.; Elk, M.; Ryu, H.; Hoelle, J.; Santo Domingo, J.W.; Konstantinidis, K.T. Biofilms on Hospital Shower Hoses: Characterization and Implications for Nosocomial Infections. Appl. Environ. Microbiol. 2016, 82, 2872–2883. [Google Scholar] [CrossRef] [Green Version]
- Dhandayuthapani, S.; Mudd, M.; Deretic, V. Interactions of OxyR with the promoter region of the oxyR and ahpC genes from Mycobacterium leprae and Mycobacterium tuberculosis. J. Bacteriol. 1997, 179, 2401–2409. [Google Scholar] [CrossRef] [Green Version]
- Baron, J.L.; Harris, J.K.; Holinger, E.P.; Duda, S.; Stevens, M.J.; Robertson, C.E.; Ross, K.A.; Pace, N.R.; Stout, J.E. Effect of monochloramine treatment on the microbial ecology of Legionella and associated bacterial populations in a hospital hot water system. Syst. Appl. Microbiol. 2015, 38, 198–205. [Google Scholar] [CrossRef]
- Hwang, C.; Ling, F.; Andersen, G.L.; LeChevallier, M.W.; Liu, W.-T. Microbial community dynamics of an urban drinking water distribution system subjected to phases of chloramination and chlorination treatments. Appl. Environ. Microbiol. 2012, 78, 7856–7865. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cheng, D.; Li, Y.; Yu, L.; Gin, K.Y.-H.; Chen, J.P.; Reinhard, M. Effects of monochloramine and hydrogen peroxide on the bacterial community shifts in biologically treated wastewater. Chemosphere 2017, 189, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Regione Emilia Romagna. Approvazione delle linee guida regionali per la sorveglianza e il controllo della legionellosi—DGR n. 828/2017. 2017. Available online: http://salute.regione.emilia-romagna.it/documentazione/leggi/regionali/dgr-2127-2016/dgr-n-828-2017-linee-guida-regionali-per-la-sorveglianza-e-il-controllo-della-legionellosi/view (accessed on 1 November 2018).
- Wang, H.; Bedard, E.; Prevost, M.; Camper, A.K.; Hill, V.R.; Pruden, A. Methodological approaches for monitoring opportunistic pathogens in premise plumbing: A review. Water Res. 2017, 117, 68–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Edwards, M.A.; Falkinham, J.O., 3rd; Pruden, A. Probiotic approach to pathogen control in premise plumbing systems? A review. Environ. Sci. Technol. 2013, 47, 10117–10128. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
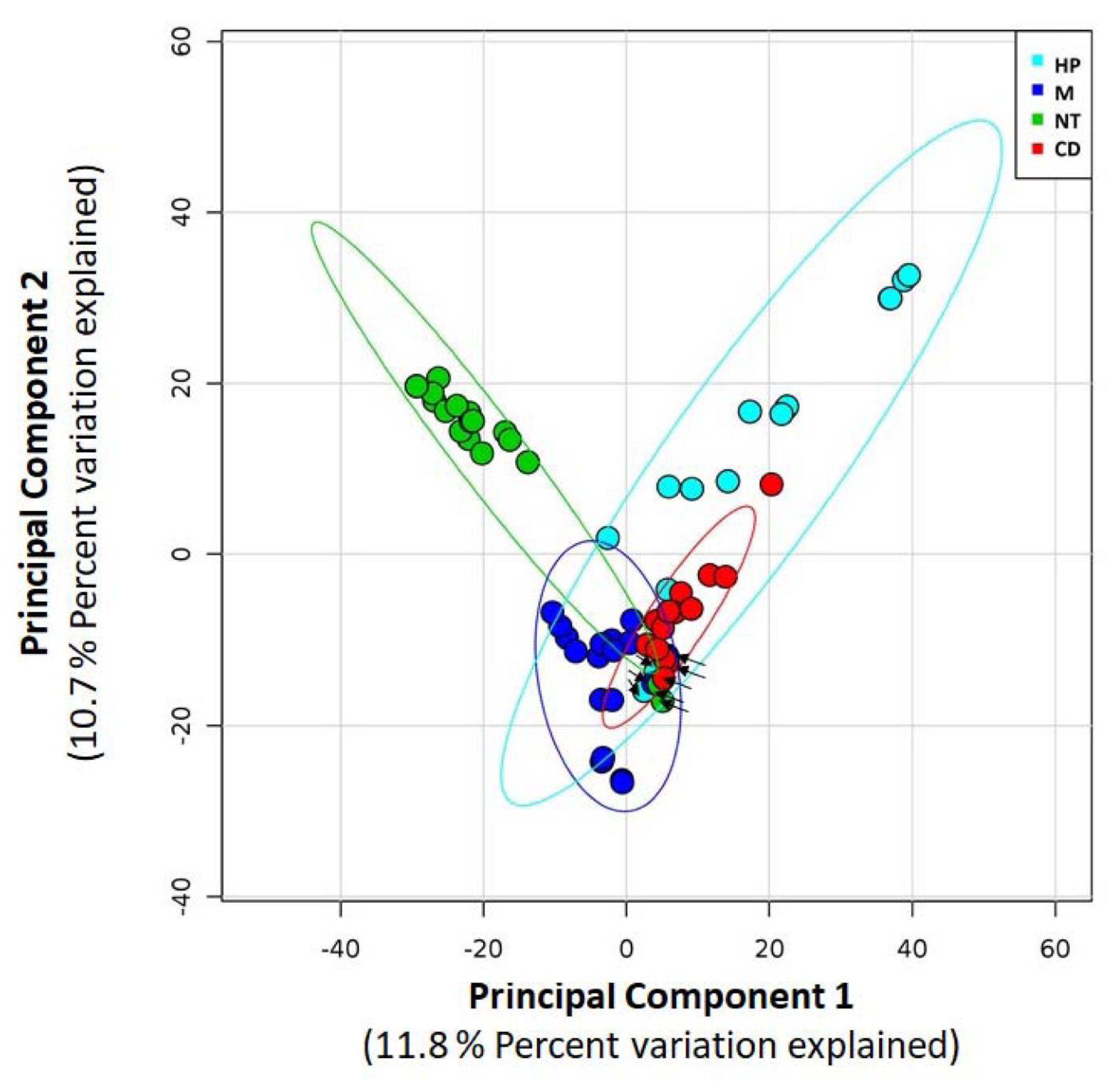{kind=link}
| Inlet Cold Water | |||||
|---|---|---|---|---|---|
| Month | Sample | t (°C) | OTUs | Shannon Index | Equitability Index |
| February | Building 1 | 13.0 | 518 | 1.967 | 0.340 |
| Building 2 | 12.9 | 664 | 2.305 | 0.327 | |
| Building 3 | 12.9 | 350 | 1.772 | 0.363 | |
| Building 4 | 12.8 | 484 | 1.503 | 0.344 | |
| mean | median | mean | mean | ||
| 12.9 | 501 | 1.887 | 0.344 | ||
| May | Building 1 | 13.8 | 291 | 2.527 | 0.267 |
| Building 2 | 13.4 | 383 | 2.507 | 0.332 | |
| Building 3 | 12.5 | 280 | 1.919 | 0.366 | |
| Building 4 | 16.7 | 376 | 2.062 | 0.393 | |
| mean | median | mean | mean | ||
| 14.1 | 334 | 2.254 | 0.340 | ||
| Hot Water Parameters | Network HP | Network M | Network CD | Network NT | p-Value |
|---|---|---|---|---|---|
| Temperature Mean (°C) (Range) | 45.2 (33.4–49.2) | 40.0 (31.0–46.3) | 42.1 (36.2–44.9) | 53.2 (50.4–56.6) | <0.0001 |
| Biocide concentration Mean (ppm) (Range) | 21.3 (18.1–25.0) | 3.0 (2.6–3.6) | 0.2 (0.1–0.4) | - | - |
| OTUs Median (Range) | 341 (274–414) | 384 (252–816) | 410 (271–956) | 509 (312–981) | 0.008 |
| Shannon index Mean (Range) | 2.316 (0.874–3.549) | 2.110 (1.488–2.674) | 1.998 (0.926–2.769) | 2.135 (1.544–2.532) | 0.481 |
| Equitability index Mean (Range) | 0.395 (0.155–0.629) | 0.353 (0.249–0.433) | 0.327 (0.165–0.487) | 0.343 (0.259–0.425) | 0.254 |
| Network | Predominant Genera | Relative Abundance (%) | ||
|---|---|---|---|---|
| Mean | Minimum | Maximum | ||
| HP | Mycobacterium | 36.36 | 1.59 | 84.50 |
| Bradyrhizobium | 8.58 | 2.23 | 22.77 | |
| Thiocystis | 3.45 | 0.43 | 11.10 | |
| M | Gallionella | 17.71 | 0.96 | 60.42 |
| Ectothiorhodospira | 10.42 | 0.44 | 27.11 | |
| Sediminibacterium | 4.24 | 1.45 | 9.87 | |
| CD | Halorhodospira | 10.35 | 0.06 | 51.93 |
| Blastomonas | 9.72 | 1.48 | 37.28 | |
| Bradyrhizobium | 8.91 | 3.37 | 25.82 | |
| NT | Chitinophaga | 11.70 | 4.04 | 21.06 |
| Methylonatrum | 6.90 | 0.02 | 50.20 | |
| Thermobaculum | 6.19 | 2.42 | 12.30 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paduano, S.; Marchesi, I.; Casali, M.E.; Valeriani, F.; Frezza, G.; Vecchi, E.; Sircana, L.; Romano Spica, V.; Borella, P.; Bargellini, A. Characterisation of Microbial Community Associated with Different Disinfection Treatments in Hospital hot Water Networks. Int. J. Environ. Res. Public Health 2020, 17, 2158. https://doi.org/10.3390/ijerph17062158
Paduano S, Marchesi I, Casali ME, Valeriani F, Frezza G, Vecchi E, Sircana L, Romano Spica V, Borella P, Bargellini A. Characterisation of Microbial Community Associated with Different Disinfection Treatments in Hospital hot Water Networks. International Journal of Environmental Research and Public Health. 2020; 17(6):2158. https://doi.org/10.3390/ijerph17062158
Chicago/Turabian StylePaduano, Stefania, Isabella Marchesi, Maria Elisabetta Casali, Federica Valeriani, Giuseppina Frezza, Elena Vecchi, Luca Sircana, Vincenzo Romano Spica, Paola Borella, and Annalisa Bargellini. 2020. "Characterisation of Microbial Community Associated with Different Disinfection Treatments in Hospital hot Water Networks" International Journal of Environmental Research and Public Health 17, no. 6: 2158. https://doi.org/10.3390/ijerph17062158