Growth Inhibition of Phaeocystis Globosa Induced by Luteolin-7-O-glucuronide from Seagrass Enhalus acoroides

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Culture Conditions
2.2. Preparation of E. acoroides Aqueous Extract
2.3. Treatments with E. acoroides Aqueous Extract
2.4. Chlorophyll-a Concentration and Photosynthetic Efficiency (Fv/Fm) Assays
2.5. Transmission Electron Microscopy (TEM) Analysis
2.6. Ultraperformance Liquid Chromatography-High Resolution Mass Spectrometry (UPLC-HRMS) Analysis
2.7. Treatments with Luteolin-7-O-glucuronide and Quantification of Luteolin-7-O-glucuronide
2.8. Data Analysis
3. Results
3.1. Effects of E. acoroides Aqueous Extract on the Growth of P. globosa
3.2. Effects of E. acoroides Aqueous Extract on Chlorophyll-a Concentration and Fv/Fm of P. globosa
3.3. Effects of E. acoroides Aqueous Extract on Subcellular Structure of P. globosa
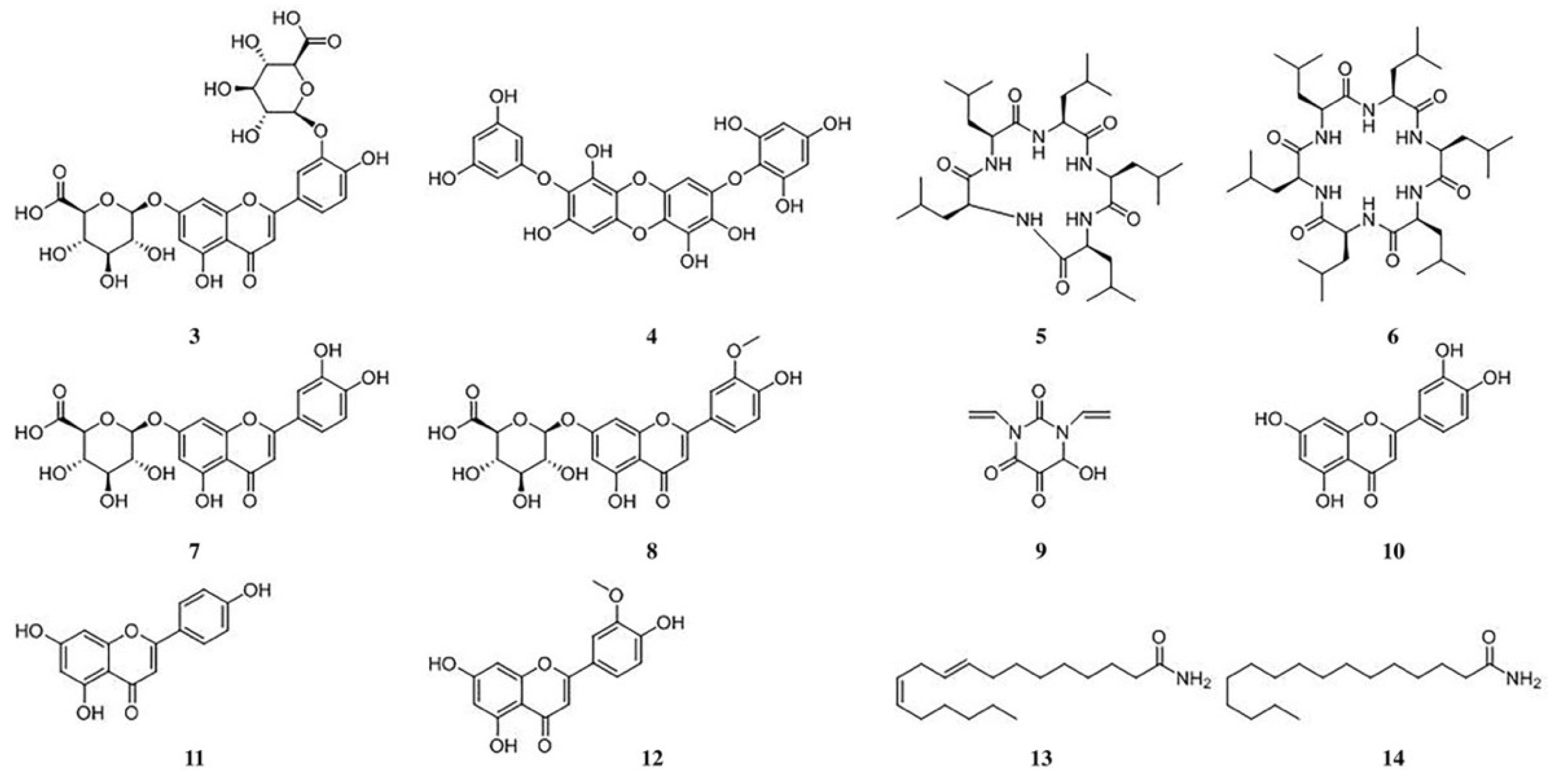
3.4. Chemical Constituent Analysis of Aqueous Extract from E. acoroides
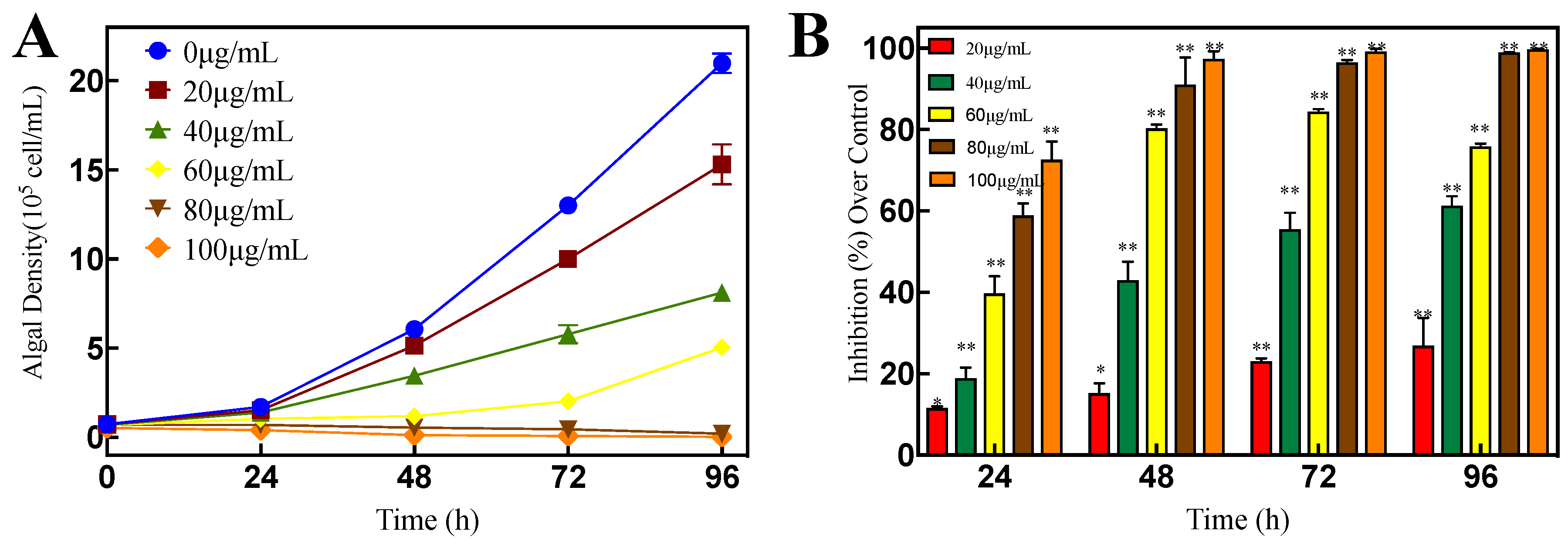
3.5. Antialgal Activity of Luteolin-7-O-glucuronide
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sellner, K.G.; Doucette, G.J.; Kirkpatrick, G.J. Harmful algal blooms: Causes impacts and detection. J. Ind. Microbiol. Biotechnol. 2003, 30, 383–406. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Hu, X.; Yin, P. Photosynthetic inhibition and oxidative stress to the toxic Phaeocystis globosa caused by a diketopiperazine isolated from products of algicidal bacterium metabolism. J. Microbiol. 2016, 54, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Blauw, A.N.; Los, F.J.; Huisman, J.; Peperzak, L. Nuisance foam events and Phaeocystis globosa blooms in Dutch coastal waters analyzed with fuzzy logic. J. Mar. Syst. 2010, 83, 115–126. [Google Scholar] [CrossRef]
- Qi, Y.; Chen, J.; Wang, Z.; Xu, N.; Wang, Y.; Shen, P.; Lu, S.; Hodgkiss, I.J. Some observations on harmful algal bloom (HAB) events along the coast of Guangdong southern China in 1998. In Asian Pacific Phycol. 21st Century Prospect Challenges; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Hai, D.; Lam, N.; Dippner, J.W. Development of Phaeocystis globosa blooms in the upwelling waters of the South Central coast of Viet Nam. J. Mar. Syst. 2010, 83, 253–261. [Google Scholar] [CrossRef]
- Lamy, D.; Obernosterer, I.; Laghdass, M.; Artigas, F.; Breton, E.; Grattepanche, J.D.; Lecuyer, E.; Degros, N.; Lebaron, P.; Christaki, U. Temporal changes of major bacterial groups and bacterial heterotrophic activity during a Phaeocystis globosa bloom in the eastern English Channel. Aquat. Microb. Ecol. 2009, 58, 95–107. [Google Scholar] [CrossRef]
- Laabir, M.; Grignon-Dubois, M.; Cecchi, P.; Rezzonico, B.; Rouquette, M.; Masseret, E. Allelopathic Effects of Zostera Spp. on the Growth and Photosynthetic Activity of the Toxic Dinoflagellate Alexandrium Catenella. In Proceedings of the 4th Mediterranean Symposium on Marine Vegetation. Regional Activity Center for Specially Protected Areas, Yasmine-Hammamet, 2–4 December 2010. [Google Scholar]
- Sun, Y.; Zhou, W.; Wang, H.; Guo, G.; Su, Z.; Pu, Y. Antialgal compounds with antialgal activity against the common red tide microalgae from a green algae Ulva pertusa. Ecotoxicol. Environ. Saf. 2018, 157, 61–66. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Gobler, C.J. The green macroalga, Ulva lactuca, inhibits the growth of seven common harmful algal bloom species via allelopathy. Harmful Algae 2011, 10, 480–488. [Google Scholar] [CrossRef]
- Short, F.; Carruthers, T.; Dennison, W.; Waycott, M. Global seagrass distribution and diversity: A bioregional model. J. Exp. Mar. Biol. Ecol. 2007, 350, 3–20. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Arumugam, R.; Anantharaman, P. Antibacterial potential of three seagrasses against human pathogens. Asian Pac. J. Trop. Med. 2010, 3, 890–893. [Google Scholar] [CrossRef] [Green Version]
- Ragupathi Raja Kannan, R.; Arumugam, R.; Thangaradjou, T.; Anantharaman, P. Phytochemical constituents, antioxidant properties and p-coumaric acid analysis in some seagrasses. Food Res. Int. 2013, 54, 1229–1236. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Arumugam, R.; Anantharaman, P. In vitro antioxidant activities of ethanol extract from Enhalus acoroides (L.F.) Royle. Asian Pac. J. Trop. Med. 2010, 3, 898–901. [Google Scholar] [CrossRef]
- Nguyen, X.V.; Tran, M.H.; Papenbrock, J. Different organs of Enhalus acoroides (Hydrocharitaceae) can serve as specific bioindicators for sediment contaminated with different heavy metals. S. Afr. J. Bot. 2017, 113, 389–395. [Google Scholar] [CrossRef]
- Zhao, M.; Xiao, H.; Sun, D.; Duan, S. Investigation of the Inhibitory Effects of Mangrove Leaves and Analysis of Their Active Components on Phaeocystis globosa during Different Stages of Leaf Age. Int. J. Environ. Res. Public Health 2018, 15, 2434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qi, S.; Qian, P.; Wang, B. Antifeedant, antibacterial, and antilarval compounds from the South China Sea seagrass Enhalus acoroides. Bot. Mar. 2008, 51, 5–7. [Google Scholar]
- Zhang, H.; Wang, H.; Zheng, W.; Yao, Z.; Peng, Y.; Zhang, S. Toxic Effects of Prodigiosin Secreted by Hahella sp. KA22 on Harmful Alga Phaeocystis globosa. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Li, C.; Huang, H.; Lee, Y.P. Inhibition effect of natural flavonoids on red tide alga Phaeocystis globosa and its quantitative structure-activity relationship. Environ. Sci. Pollut. Res. 2019, 1, 14. [Google Scholar] [CrossRef]
- St-pierre, A.; Blondeau, D.; Lajeunesse, A.; Bley, J.; Bourdeau, N.; Desgagn, I. Phytochemical Screening of Quaking Aspen (Populus tremuloides) Extracts by UPLC-QTOF-MS and Evaluation of their Antimicrobial Activity. Molecules 2018, 23, 1739. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Barros, L.; Santos-Buelga, C.; Silva, S.; Henriques, M.; Ferreira, I.C.F.R. Decoction, infusion and hydroalcoholic extract of cultivated thyme: Antioxidant and antibacterial activities, and phenolic characterisation. Food Chem. 2015, 167, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salawu, S.O.; Ogundare, A.O.; Ola-Salawu, B.B.; Akindahunsi, A.A. Antimicrobial activities of phenolic containing extracts of some tropical vegetables. Afr. J. Pharm. Pharmacol. 2011, 5, 486–492. [Google Scholar]
- Hua, Q.; Liu, Y.G.; Yan, Z.L.; Zeng, G.M.; Liu, S.B.; Wang, W.J.; Tan, X.G.; Deng, J.Q.; Tang, X.; Wang, Q.P. Allelopathic effect of the rice straw aqueous extract on the growth of Microcystis aeruginosa. Ecotoxicol. Environ. Saf. 2018, 148, 953–959. [Google Scholar] [CrossRef]
- Tazart, Z.; Douma, M.; Tebaa, L.; Loudiki, M. Use of macrophytes allelopathy in the biocontrol of harmful Microcystis aeruginosa blooms. Water Sci. Technol. Water Supply 2018, 19, 245–253. [Google Scholar] [CrossRef]
- Jeong, J.; Jin, H.; Sohn, C.H.; Suh, K.; Hong, Y. Algicidal activity of the seaweed Corallina pilulifera against red tide microalgae. J. Appl. Phycol. 2000, 12, 37–43. [Google Scholar] [CrossRef]
- Loura, P.; Singh, R. Allelopathic activity of hexane and ethyl acetate fractions of crude extract of leaves, bark and seeds of Albizia-lebbek L (Benth.). IJCS 2018, 6, 766–768. [Google Scholar]
- Wang, R.; Tang, X.; Feng, L.; Xiao, H.; Qu, L.; Cai, H. Inhibitory effect of Surgassum thunbergii on Heterosigma akashiwo and Skeletonema costatum. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2006, 17, 2421–2425. [Google Scholar]
- Chen, S.; Zheng, T.; Ye, C.; Huannixi, W.; Yakefu, Z.; Meng, Y.; Peng, X.; Tian, Z.; Wang, J.; Ma, Y.; et al. Algicidal properties of extracts from Cinnamomum camphora fresh leaves and their main compounds. Ecotoxicol. Environ. Saf. 2018, 163, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xing, J.; Zhang, J.; Zhou, W.; Pu, Y. Sesquiterpenoids with antialgal activity against the common red tide microalgae from marine macroalga Porphyra yezoensis. Environ. Sci. Pollut. Res. 2018, 25, 7844–7859. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Xie, H.; Gong, Y.; Wang, Q.; Yang, Y. Secondary metabolites from the seaweed Gracilaria lemaneiformis and their allelopathic effects on Skeletonema costatum. Biochem. Syst. Ecol. 2011, 39, 397–400. [Google Scholar] [CrossRef]
- Botanik, L.; Wiirzburg, U. Assessment of Photosystem I1 Photochemical Quantum Yield by Chlorophyll Fluorescence Quenching Analysis. Funct. Plant Biol. 1995, 22, 209–220. [Google Scholar]
- Zhang, H.; Zhang, S.; Peng, Y.; Li, Y. Effectiveness and toxicity of a novel isolated actinomycete strain Streptomyces sp. JS01 on a harmful alga Phaeocystis globosa. Appl. Microbiol. Biotechnol. 2015, 99, 4807–4814. [Google Scholar] [CrossRef]
- Zhang, B.; Cai, G.; Wang, H.; Li, D.; Yang, X.; An, X.; Zheng, X.; Tian, Y. Streptomyces alboflavus RPS and Its Novel and High Algicidal Activity against Harmful Algal Bloom Species Phaeocystis globosa. PLoS ONE 2014, 9, e92907. [Google Scholar] [CrossRef]
- Nakamura, Y.; Sano, M. Comparison between community structures of fishes in Enhalus acoroides-and Thalassia hemprichii-dominated seagrass beds on fringing coral reefs in the Ryukyu Islands, Japan. Ichthyol. Res. 2004, 51, 38–45. [Google Scholar] [CrossRef]
- Kirakosyan, A.; Kaufman, P.B. Recent Advances in Plant Biotechnology; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Sareedenchai, V.; Zidorn, C. Flavonoids as chemosystematic markers in the tribe Cichorieae of the Asteraceae. Biochem. Syst. Ecol. 2010, 38, 935–957. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Flavonoid and phenolic glycosides from Salvia officinalis. Phytochemistry 2000, 55, 263–267. [Google Scholar] [CrossRef]
- Zhang, H.; Peng, Y.; Zhang, S.; Cai, G.; Li, Y.; Yang, X.; Yang, K. Algicidal Effects of Prodigiosin on the Harmful Algae Phaeocystis globosa. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gulluce, M.; Karadayi, M.; Guvenalp, Z.; Ozbek, H.; Arasoglu, T.; Baris, O. Isolation of some active compounds from Origanum vulgare L. ssp. vulgare and determination of their genotoxic potentials. Food Chem. 2012, 130, 248–253. [Google Scholar] [CrossRef]
- Orhan, F.; Çeker, S.; Anar, M.; Agar, G.; Arasoglu, T. Protective effects of three luteolin derivatives on aflatoxin B1-induced genotoxicity on human blood cells. Med. Chem. Res. 2016, 25, 2567–2577. [Google Scholar] [CrossRef]
- Jeyadevi, R.; Sivasudha, T.; Rameshkumar, A.; Dinesh Kumar, L. Anti-arthritic activity of the Indian leafy vegetable Cardiospermum halicacabum in Wistar rats and UPLC--QTOF--MS/MS identification of the putative active phenolic components. Inflamm. Res. 2013, 62, 115–126. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | Exact Mass (m/z) [M + H]+ [M − H]− | Types | Molecular Formula | Name of the Chemical Constituent | |
|---|---|---|---|---|---|---|
| 1 | 2.83 | 614.1519 | 612.1376 | polypeptides | polypeptide (unknown) | |
| 2 | 2.90 | 614.1519 | 612.1376 | polypeptides | polypeptide (unknown) | |
| 3 | 3.00 | 639.1227 | 637.1060 | flavonoids | C27H26O18 | luteolin-3′,7-O-diglucuronides |
| 4 | 3.22 | 513.0708 | 511.0551 | tannins | C24H16O13 | diphlorethohydroxycarmalol |
| 5 | 3.27 | 566.4286 | 610.4195 | homocyclic peptides | C30H55N5O5 | cyclo (l-leucyl-l-leucyl-l-leucyl-l-leucyl-l-leucyl) |
| 6 | 3.42 | 679.5132 | 723.5030 | homocyclic peptides | C36H66N6O6 | cyclo (l-leucyl-l-leucyl-l-leucyl-l-leucyl-l-leucyl-l-leucyl) |
| 7 | 3.55 | 463.0871 | 461.0717 | flavonoids | C21H18O12 | luteolin-7-O-glucuronide |
| 8 | 3.66 | 475.0868 | flavonoids | C22H20O12 | chrysoeriol-7-O-glucuronide | |
| 9 | 3.95 | 183.0794 | nitrogen heterocyclic compounds | C8H8N2O4 | 6-hydroxy-1,3-divinyldihydropyrimidine-2,4,5(3H)-trione | |
| 10 | 4.07 | 287.0548 | 285.0419 | flavonoids | C15H10O6 | luteolin |
| 11 | 4.48 | 271.0620 | 269.0449 | flavonoids | C15H10O5 | apigenin |
| 12 | 5.18 | 299.0556 | flavonoids | C16H12O6 | chrysoeriol | |
| 13 | 9.36 | 280.2655 | aliphatic amides | C18H33NO | (9E,12Z)-octadeca-9,12-dienamide | |
| 14 | 10.04 | 256.2672 | aliphatic amides | C16H33NO | palmitamide | |
| No. | Rt (min) | Types | Molecular Formula | Name of the Chemical Constituent | Peak Area |
|---|---|---|---|---|---|
| 3 | 3.00 | flavonoids | C27H26O18 | luteolin-3′,7-O-diglucuronides | 430,617 |
| 7 | 3.55 | C21H18O12 | luteolin-7-O-glucuronide | 268,581 | |
| 8 | 3.66 | C22H20O12 | chrysoeriol-7-O-glucuronide | 623 | |
| 10 | 4.07 | C15H10O6 | luteolin | 48,305 | |
| 11 | 4.48 | C15H10O5 | apigenin | 41,554 | |
| 12 | 5.18 | C16H12O6 | chrysoeriol | 4996 |
| Rt (min) | Molecular Formula | Compound | Content (μg/mL) |
|---|---|---|---|
| 3.55 | C21H18O12 | luteolin-7-O-glucuronide | 68.125 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Xiao, H.; Chen, Q.; Zhao, M.; Sun, D.; Duan, S. Growth Inhibition of Phaeocystis Globosa Induced by Luteolin-7-O-glucuronide from Seagrass Enhalus acoroides. Int. J. Environ. Res. Public Health 2019, 16, 2615. https://doi.org/10.3390/ijerph16142615
Zhu J, Xiao H, Chen Q, Zhao M, Sun D, Duan S. Growth Inhibition of Phaeocystis Globosa Induced by Luteolin-7-O-glucuronide from Seagrass Enhalus acoroides. International Journal of Environmental Research and Public Health. 2019; 16(14):2615. https://doi.org/10.3390/ijerph16142615
Chicago/Turabian StyleZhu, Jingyi, Han Xiao, Qi Chen, Min Zhao, Dong Sun, and Shunshan Duan. 2019. "Growth Inhibition of Phaeocystis Globosa Induced by Luteolin-7-O-glucuronide from Seagrass Enhalus acoroides" International Journal of Environmental Research and Public Health 16, no. 14: 2615. https://doi.org/10.3390/ijerph16142615