
Changes of Sand Fly Populations and Leishmania infantum Infection Rates in an Irrigated Village Located in Arid Central Tunisia

Abstract
:1. Introduction
2. Materials and Methods
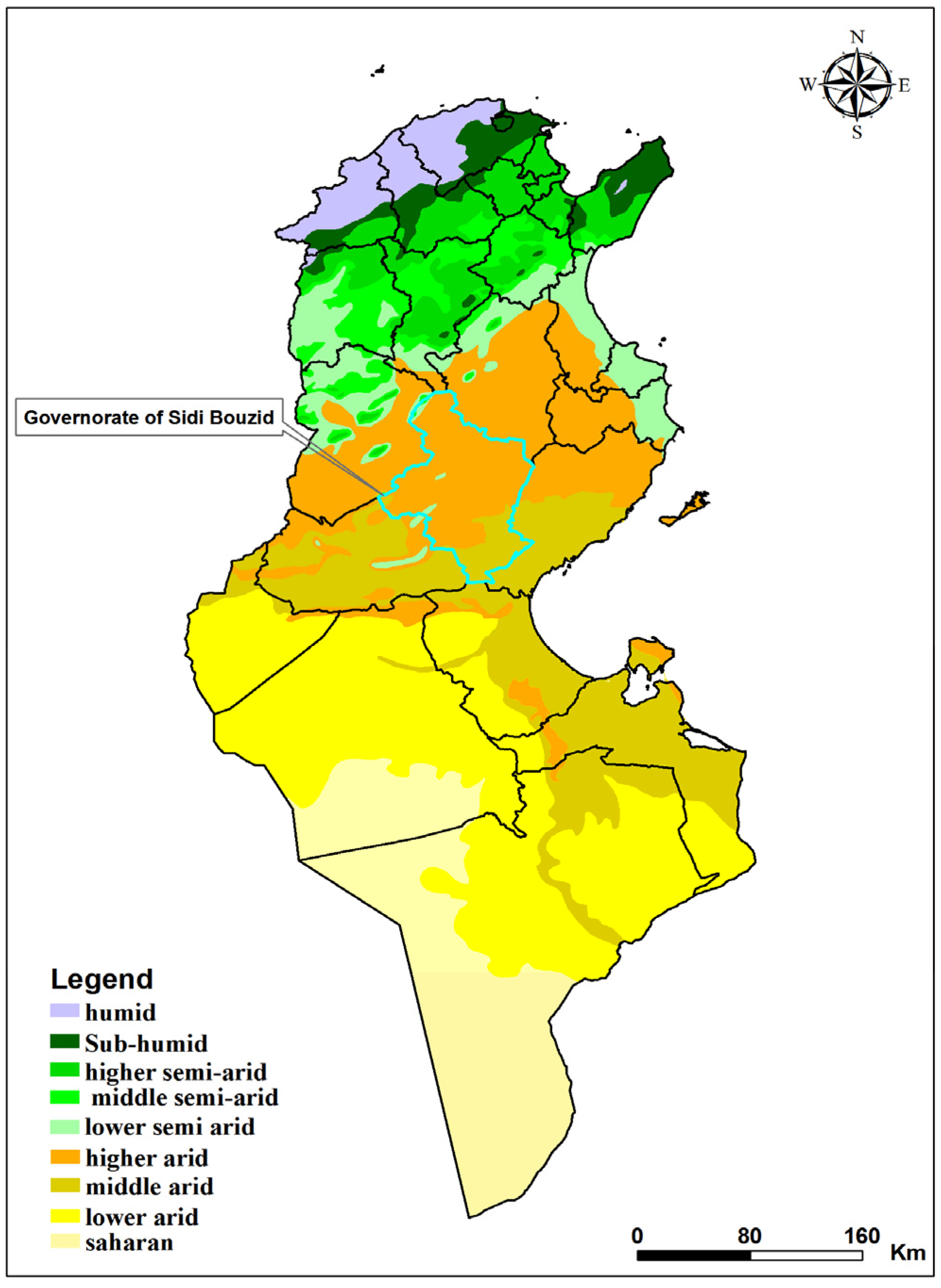
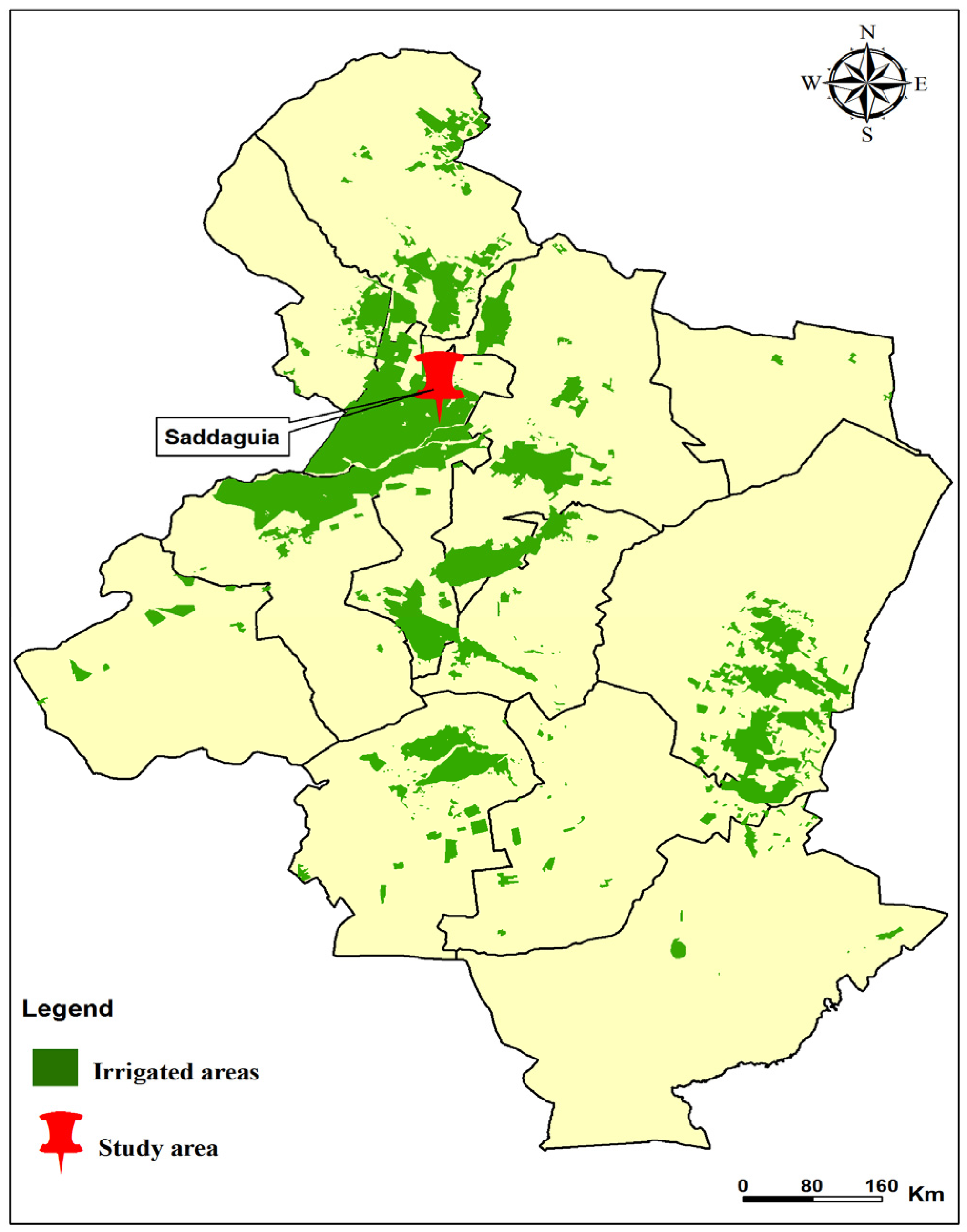
2.1. Study Site
2.2. Collection and Identification of Sand Flies
2.3. Detection of Leishmania Spp in Female Sand Flies
2.4. DNA Sequencing
3. Results
3.1. Sand Fly Species and Relative Abundance
3.2. Leishmania Infection in Sand Flies
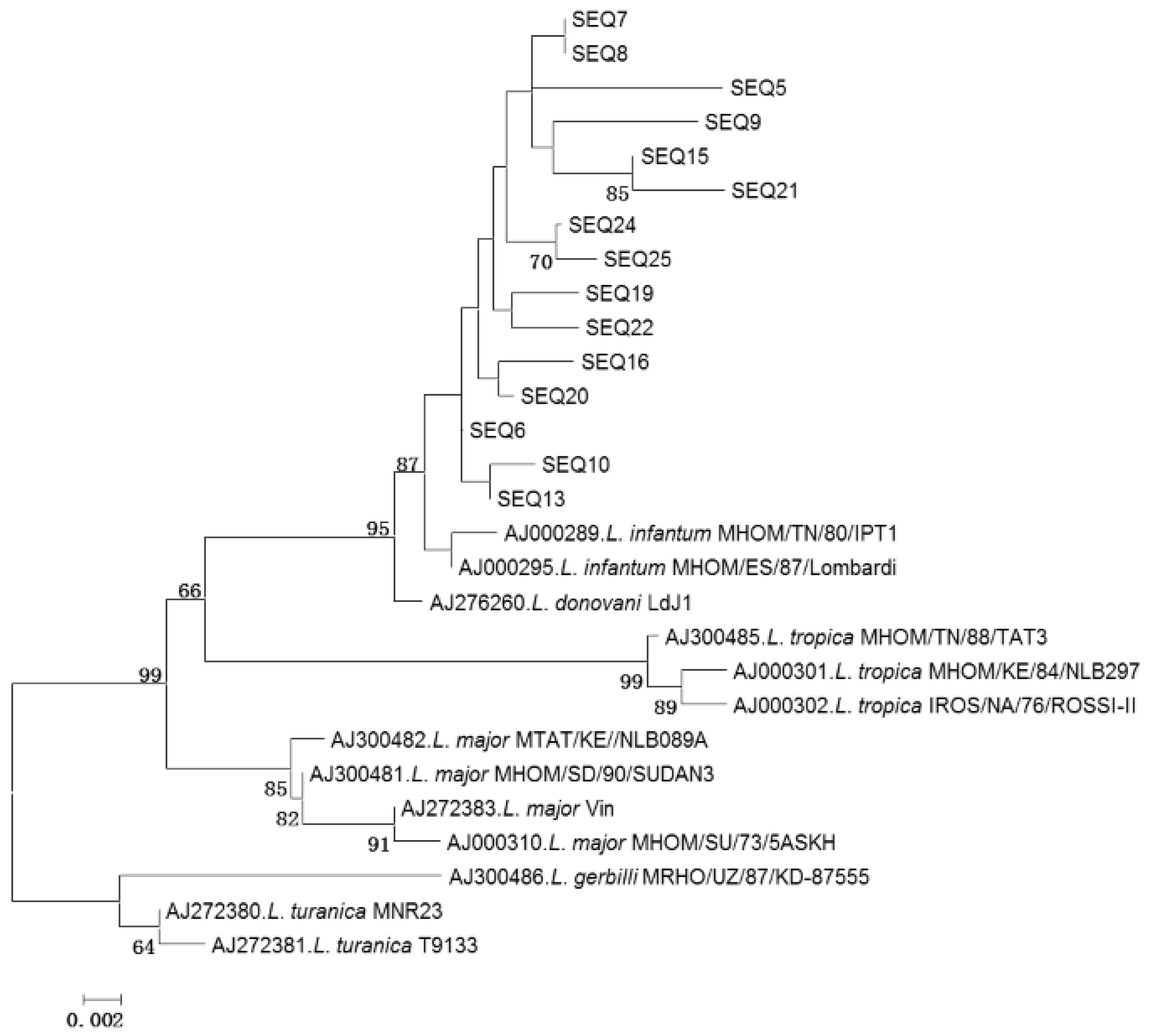
3.3. Leishmania Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [PubMed]
- Killick-Kendrick, R. Phlebotomine vector of the leishmaniases: A review. Med. Vet. Entomol. 1990, 4, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Ben Ismail, R. Incrimination de Phlebotomus perniciosus comme vecteur de Leishmania infantum. Arch. Inst. Pasteu. Tunis 1993, 70, 91–110. [Google Scholar]
- Izri, M.A.; Belazzoug, S.; Boudjebla, Y.; Dereure, J.P.; Pratlong, F.; Delabre-Belmonte, A.; Rioux, J.A. Leishmania infantum MON1 isolé de Phlebotomus perniciosus en Kabylie (Algérie). Ann. Parasitol. Hum. Comp. 1990, 65, 151–152. [Google Scholar] [PubMed]
- Es-Sette, N.; Ajouad, M.; Laamarani-Idrissi, A.; Mellouk, F.; Lemrani, A. Molecular detection and identification of Leishmania infection in naturally infected sand flies in a focus of cutaneous leishmaniasis in northern Morocco. Parsit. Vectors 2014, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zoghlami, Z.; Chouihi, E.; Barhoumi, W.; Dachraoui, K.; Massoudi, N.; Ben Helel, K.; Habboul, Z.; Hadhri, M.H.; Limam, S.; Mhadhbi, M.; et al. Interaction between canine and human visceral leishmaniases in a holoendemic focus of Central Tunisia. Acta Trop. 2014, 139, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; Den Boer, M. The WHO leishmaniasis control team. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C. Chronique du kala azar in Tunisia. Arch. Inst. Pasteur Tunis 1938, 27, 97–104. [Google Scholar]
- Chadli, A.; Ben Rachid, M.S.; Fhaïel, A. Chronique des leishmanioses en Tunisie. Arch. Inst. Pasteur Tunis 1968, 45, 1–14. [Google Scholar]
- Ben Rachid, M.S.; Hamza, B.; Tabbane, C.; Gharbi, R.; Jedidi, H. Etat actuel des leishmanioses en Tunisie. Ann. Soc. Belg. Méd. Trop. 1983, 63, 29–40. [Google Scholar] [PubMed]
- Rioux, J.A.; Lanotte, G.; Petter, F.; Deureure, J.; Akalay, O.; Pratlong, F.; Velez, I.D.; Fikri, N.B.; Maazoun, R.; Denial, M.; et al. Les leishmanioses cutanées du bassin Méditerranéen occidental, de l’identification enzymatique à l’analyse éco-épidémiologique. L’expemple de trois «foyers» tunisien, marocain et français. In Leishmania. Taxonomie et Phylogénèse, In Proceedings of the Applications éco-Epidémiologiques, International Colloquium CNRS/INSERM, Montpellier, France, 2–6 July 1984; IMEEE: Montpellier, France, 1986; pp. 471–478. [Google Scholar]
- Keddous, A. Enquête éco-Epidémilogique sur le Kala Azar Dans le Gouvernorat de Sidi Bouzid. Etude d’un Réservoir Canin. Ph.D. Thesis, Medical School of Tunis, Tunis, Tunisia, July 1988. [Google Scholar]
- Ayadi, A.; Ben Ismail, R.; Ben Rachid, M.S. Extension de l’aire de transmission du Kala Azar à Leishmania infantum (Nicolle 1908) vers le centre et le sud de la Tunisie. Arch. Inst. Pasteur Tunis 1991, 68, 269–273. [Google Scholar] [PubMed]
- Besbes, A.; Pousse, H.; Ben Said, M.; Kharrat, H.; Chenimi, L. Leishmanioses viscérales infantiles du centre Tunisien (221 cas). Méd. Mal. Infect. 1994, 24, 628–634. [Google Scholar] [CrossRef]
- Bouratbine, A.; Aoun, K.; Chahed, M.K.; Ben Ismail, R. Données épidémiologiques sur la leishmaniose viscerale infantile en Tunisie en 1993. Méd. Mal. Infect. 1998, 28, 446–447. [Google Scholar] [CrossRef]
- Ben Salah, A.; Ben Ismail, R.; Amri, F.; Chlif, S.; Ben Rzig, F.; Karrat, H.; Hadhri, H.; Hassouna, M.; Dellagi, K. Investigation of the spread of human visceral leishmaniasis in central Tunisia. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 382–386. [Google Scholar] [CrossRef]
- Ben Ahmed, K.B.; Aoun, K.; Jeddi, F.; Ghrab, J.; El-Aroui, M.A.; Bouratbine, A. Visceral leishmaniasis in Tunisia: Spatial distribution and association with climatic factors. Am. J. Trop. Med. Hyg. 2009, 8, 40–45. [Google Scholar]
- Zhioua, E.; Kaabi, B.; Chelbi, I. Entomological investigations following the spread of visceral leishmaniasis in Tunisia. J. Vector Ecol. 2007, 32, 371–374. [Google Scholar] [CrossRef]
- Barhoumi, W.; Qualls, W.A.; Archer, R.S.; Fuller, D.O.; Chelbi, I.; Cherni, S.; Derbali, M.; Arheart, K.L.; Zhioua, E.; Beier, J.C. Irrigation in the arid regions of Tunisia impacts the abundance and apparent density of sand fly vectors of Leishmania infantum. Acta Trop. 2014, 141, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Ben Ismail, R.; Gramiccia, M.; Gradoni, L.; Helal, H.; Ben Rachid, M.S. Isolation of Leishmania major from Phlebotomus papatasi in Tunisia. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 749. [Google Scholar] [CrossRef]
- Chelbi, I.; Derbali, M.; Al-Ahmadi, Z.; Zaafouri, B.; El Fahem, A.; Zhioua, E. Phenology of Phlebotomus papatasi (Diptera: Psychodidae) relative to the seasonal prevalence of zoonotic cutaneous leishmaniasis in Central Tunisia. J. Med. Entomol. 2007, 44, 385–388. [Google Scholar] [CrossRef]
- Ben Salah, A.; Kamarianakis, Y.; Chlif, S.; Ben Alya, N.; Prastacos, P. Zoonotic cutaneous leishmaniasis in central Tunisia: Spatio-temporal dynamics. Int. J. Epidemiol. 2007, 36, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Chelbi, I.; Kaabi, B.; Bejaoui, M.; Derbali, M.; Zhioua, E. Spatial correlation between Phlebotomus papatasi Scopoli (Diptera: Psychodidae) and incidence of zoonotic cutaneous leishmaniasis in Tunisia. J. Med. Entomol. 2009, 46, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, W.; Chelbi, I.; Zhioua, E. Effects of the development of irrigation systems in the arid areas on the establishment of Phlebotomus (Larroussius) perfiliewi Parrot, 1939. Bull. Soc. Pathol. Exot. 2012, 105, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Dachraoui, K.; Piorkowski, G.; Chelbi, I.; Moureau, G.; Cherni, S.; de Lamballerie, X.; Sakhria, S.; Charrel, R.N.; Zhioua, E. Isolation of Toscana virus from sand flies, Tunisia. Emerg. Infect. Dis. 2013, 19, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Croset, H.; Rioux, J.A.; Master, M.; Bayar, N. Les phlébotomes de la Tunisie (Diptera, Phlebotominae). Mise au point systématique, chorologique et éthologique. Ann. Parasitol. Hum. Comp. 1978, 53, 711–749. [Google Scholar] [PubMed]
- Léger, N.; Pesson, B.; Madulo-Leblond, G.; Abonnenc, E. Sur la différentiation des femelles du sous-genre Larroussius Nitzulescu, 1931 (Diptera: Phlebotominae) de la région méditerranéenne. Ann. Parasitol. Hum. Comp. 1983, 58, 611–623. [Google Scholar] [PubMed]
- Pesson, B.; Ready, J.S.; Benabdennbi, I.; Martín-Sánchez, J.; Esseghir, S.; Cadi-Soussi, M.; Morillas-Marquez, F.; Ready, P.D. Sandflies of the Phlebotomus perniciosus complex: Mitochondrial introgression and a new sibling species of Phlebotomus longicuspis in the Moroccan Rif. Med. Vet. Entomol. 2004, 18, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Boussaa, S.; Boumezzough, A.; Remy, P.E.; Glasser, N.; Pesson, B. Morphological and isoenzymatic differentitation of Phlebotomus pernicious and Phlebotomus longicuspis (Diptera: Psychodidae) in Southern Morocco. Acta Trop. 2008, 106, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Mauricio, I.; Aransay, A.M.; Miles, M.A.; Ready, P.D. First detection of Leishmania major in peridomestic Phlebotomus papatasi from Isfahan province, Iran: Comparison of nested PCR of nuclear ITS ribosomal DNA and semi-nested PCR of minicircle kinetoplast DNA. Acta Trop. 2005, 93, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Ready, P.D. Nested PCRs and sequencing of nuclear ITS-rDNA fragments detect three Leishmania species of gerbils in sandflies from Iranian foci of zoonotic cutaneous leishamniasis. Trop. Med. Int. Health 2008, 13, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Moradi, G.; Akbari, G.; Farhmand, M.; Ready, P.; Piazak, N.; Assmar, M.; Amirkhani, A. PCR detection and sequencing of parasite ITS-rDNA gene from reservoir host of zoonotic cutaneous leishmaniasis in central Iran. Parasitol. Res. 2008, 103, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetic Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Chelbi, I.; Zhioua, E. Confirmation de la présence en Tunisie de Sergentomyia (Sintonius) clydei (Sinton, 1928). Bull. Soc. Pathol. Exot. 2012, 105, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Dancesco, P.; Dedet, J.P.; Ben Osman, F.; Chadli, A. Les phlébotomes capturés dans des foyers de leishmaniose canine à Tunis. Rôle probable de Phlebotomus perniciosus et Phlebotomus perfiliewi dans la transmission. Arch. Inst. Pasteur Tunis 1970, 52, 65–88. [Google Scholar]
- Ghrab, J.; Rhim, A.; Bhach-Hamba, D.; Chahed, M.K.; Aoun, K.; Nouira, S.; Bouratbine, A. Phlebotominae (Diptera: Psychodidae) of human leishmaniasis sites in Tunisia. Parasite 2006, 13, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Belazzoug, S.; Mahzoul, D.; Rioux, J.A. Les phlébotomes (Diptera, Psychodidae) de M’Sila et Bou-Saada. Arch. Inst. Pasteur Alger. 1986, 55, 117–124. [Google Scholar] [PubMed]
- Russo, J.; Rioux, J.A.; Lambert, M.; Rispal, P.; Belmonte, A.; Bechri, S. Chorologie des phlébotomes de l’Est Algérien (Diptera, Psychodidae). Ann. Parasitol. Hum. Comp. 1991, 66, 247–251. [Google Scholar]
- Maroli, M.; Gramiccia, M.; Gradoni, L. Natural infections of sand fly Phlebotomus perfiliewi with Leishmania infantum in a cutaneous leishmaniasis focus of the Abruzzi region, Italy. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 596–598. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Tarallo, V.D.; Latrofa, M.S.; Falchi, A.; Lia, R.P.; Otranto, D. Ecology of phlebotomine sand flies and Leishmania infantum infection in a rural area of southern Italy. Acta Trop. 2014, 137, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Izri, M.; Belazzoug, S. Phlebotomus (Larroussius) perfiliewi naturally infected with dermatropic Leishmania infantum at Tenes, Algeria. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 399. [Google Scholar] [CrossRef]
- Maroli, M.; Gramiccia, M.; Gradoni, L.; Troiani, M.; Ascione, R. Natural infection of Phlebotomus perniciosus with an enzymatic variant of Leishmania infantum in the Campania region of Italy. Acta Trop. 1994, 57, 333–335. [Google Scholar] [CrossRef]
- Chargui, N.; Houas, N.; Slama, D.; Gorcii, M.; Jaouadi, K.; Essabah-Aguir, N.; Mezhoud, H.; Baba, H. Transmission of visceral leishmaniasis in a previously non-endemic region of Tunisia: Detection of Leishmmania DNA in Phlebotomus perniciosus. J. Vector Ecol. 2012, 38, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Deureure, J.; Velez, I.D.; Partlong, F. La leishmaniose viscérale autochtone au Maroc méridional. Présence de Leishmania infantum MON-1 chez le chien en zone présaharienne. In Leishmania. Taxonomie et Phylogenèse, In Proceedings of the Applications éco-Epidémiologiques, International Colloquium CNRS/INSERM, Montpellier, France, 2–6 July 1984; IMEEE: Montpellier, France, 1986; pp. 421–425. [Google Scholar]
- Rioux, J.A.; Akalay, O.; Perieres, J.; Deureure, J.; Mahjour, J.; Le Houerou, H.N.; Leger, N.; Desjeux, P.; Gallego, M.; Saddiki, A.; et al. L’évaluation éco-épidémiologique du «risque leishmanien au Sahara atlantique marocain. Intérêt heuristique de la relation phlébotomes—Bioclimats». Ecol. Mediterr. 1997, 23, 73–92. [Google Scholar]
- Berdjane-Brouk, Z.; Charrel, R.N.; Hamrioui, B.; Izri, A. First detection of Leishmania infantum DNA in Phlebotomus longicuspis Nitzulescu, 1930 from visceral leishmaniasis endemic focus in Algeria. Parasitol. Res. 2012, 111, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Shaw, J.J. Evolution, classification and geographical distribution. In The Leishmaniases in Biology and Medicine; Peters, W., Killick-Kendrick, R., Eds.; Academic Press: London, UK, 1987; pp. 1–120. [Google Scholar]
- Maia, C.; Parreira, R.; Cristovao, J.M.; Freitas, F.B.; Afonso, O.; Campino, L. Molecular detection of Leishmania DNA and identification of blood meals in wild-caught phlebotomine sand flies (Diptera: Psychodidae) from Southern Portugal. Parasit. Vectors 2015, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aransay, A.M.; Scoulica, E.; Tselentis, Y. Detection and identification of Leishmania DNA with naturally infected sand flies by seminested PCR on minicircle kinetoplastic DNA. App. Environ. Microbiol. 2000, 66, 1933–1938. [Google Scholar] [CrossRef]
- Yavar, R.; Hadi, K.; Reza, A.M.; Mohebali, M.; Hasan, B.; Ali, O.M.; Sina, R.; Habib, B.H.; Abodolrahim, H.; Manuchehr, G. First detection of Leishmania infantum DNA in wild caught Phlebotomus papatasi in endemic focus of cutaneous leishmaniasis, south of Iran. Asian Pac. J. Trop. Biomed. 2013, 3, 825–829. [Google Scholar] [CrossRef]
- Killick-Kendrick, R.; Ward, R.D. Ecology of Leishmania. In Proceedings of the 3rd European Multicolloqium of Parasitology, Cambridge, UK, 7–13 September 1980.
- Bongiorno, G.; Habluetzel, A.; Khoury, C.; Maroli, M. Host preferences of phlebotomine sand flies at a hypoendemic focus of canine leishmaniasis in central Italy. Acta Trop. 2003, 88, 109–116. [Google Scholar] [CrossRef]
- Rossi, E.; Bongiorno, G.; Ciolli, E.; Di Muccio, T.; Scalone, A.; Gramiccia, M.; Gradoni, L.; Maroli, M. Seasonal phenology, host-blood feeding preferences and natural infection of Phlebotomus perniciosus (Diptera, Psychodidae) in a high-endemic focus of canine leishmaniasis in Rome province, Italy. Acta Trop. 2008, 105, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Qualls, W.A.; Muller, G.C.; Khallaayoune, K.; Revay, E.E.; Zhioua, E.; Kravchenko, V.D.; Arheart, K.L.; Xue, R.D.; Schlein, Y.; Hausmann, A.; et al. Control of sand flies with attractive toxic sugar baits (ATSB) and potential impact on non-target organisms in Morocco. Parasit. Vectors 2015, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Code of Positive Pool | Date of Collection | Biotope | Number of Female Sandflies/Pool | Leishmania Species |
|---|---|---|---|---|
| Farm A | ||||
| T 3 | 4 September 2014 | Bedroom | 1. P. perniciosus | L. infantum |
| T 20 | 18 September 2014 | Bedroom | 3. P. longicuspis | L. infantum |
| T 23 | 18 September 2014 | Bedroom | 1. S. minuta | L. infantum |
| T 33 | 7 October 2014 | Sheep shelter | 1. P. longicuspis | L. infantum |
| T 36 | 7 October 2014 | Chicken coop | 2. P. longicuspis | L. infantum |
| Farm B | ||||
| T 61 | 18 September 2014 | Sheep shelter | 9. P. perfiliewi | L. infantum |
| T 63 | 7 October 2014 | Bedroom | 24. P. perfiliewi | - |
| T 68 | 7 October 2014 | Sheep shelter | 13. P. perfiliewi | - |
| T 72 | 21 October 2014 | Sheep shelter | 30. P. papatasi | L. infantum |
| T 74 | 21 October 2014 | Sheep shelter | 30. P. perfiliewi | - |
| Farm C | ||||
| T 8 | 4 September 2014 | Rabbit hole | 4. P. papatasi | L. infantum |
| T 93 | 4 September 2014 | Chicken coop | 30. P. perfiliewi | L. infantum |
| T 110 | 4 September 2014 | Sheep and cattle shelter | 30. P. perfiliewi | - |
| T 112 | 4 September 2014 | Sheep and cattle shelter | 1. P. perniciosus | - |
| T 128 | 18 September 2014 | Sheep and cattle shelter | 26. P. perniciosus | L. infantum |
| T 130 | 18 September 2014 | Sheep and cattle shelter | 9. P. longicuspis | L. infantum |
| T 131 | 18 September 2014 | Sheep and cattle shelter | 4. P. papatasi | L. infantum |
| T 133 | 18 September 2014 | Bedroom | 7. P. perniciosus | L. infantum |
| T 135 | 18 September 2014 | bedroom | 1. P. papatasi | - |
| T 140 | 7 October 2014 | Rabbit hole | 7. P. perniciosus | L. infantum |
| T 145 | 7 January 2014 | Sheep shelter | 12. P. perfiliewi | L. infantum |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barhoumi, W.; Fares, W.; Cherni, S.; Derbali, M.; Dachraoui, K.; Chelbi, I.; Ramalho-Ortigao, M.; Beier, J.C.; Zhioua, E. Changes of Sand Fly Populations and Leishmania infantum Infection Rates in an Irrigated Village Located in Arid Central Tunisia. Int. J. Environ. Res. Public Health 2016, 13, 329. https://doi.org/10.3390/ijerph13030329
Barhoumi W, Fares W, Cherni S, Derbali M, Dachraoui K, Chelbi I, Ramalho-Ortigao M, Beier JC, Zhioua E. Changes of Sand Fly Populations and Leishmania infantum Infection Rates in an Irrigated Village Located in Arid Central Tunisia. International Journal of Environmental Research and Public Health. 2016; 13(3):329. https://doi.org/10.3390/ijerph13030329
Chicago/Turabian StyleBarhoumi, Walid, Wasfi Fares, Saifedine Cherni, Mohamed Derbali, Khalil Dachraoui, Ifhem Chelbi, Marcelo Ramalho-Ortigao, John C. Beier, and Elyes Zhioua. 2016. "Changes of Sand Fly Populations and Leishmania infantum Infection Rates in an Irrigated Village Located in Arid Central Tunisia" International Journal of Environmental Research and Public Health 13, no. 3: 329. https://doi.org/10.3390/ijerph13030329