

Maximizing Polysaccharides and Phycoerythrin in Porphyridium purpureum via the Addition of Exogenous Compounds: A Response-Surface-Methodology Approach
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of Single Factors (CG, MG, and BT) on Microalgal Growth and the Accumulation of Bioactive Substances
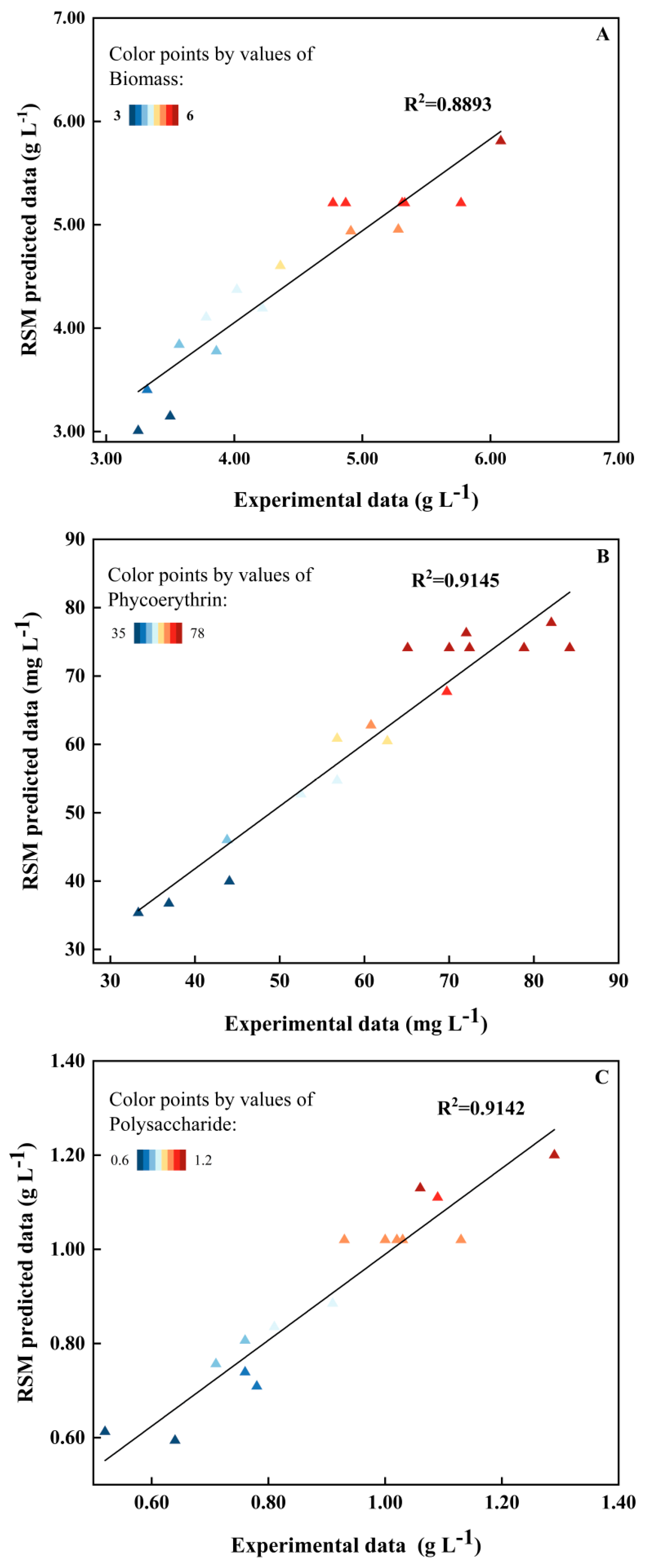
2.2. Model Fitting of RSM
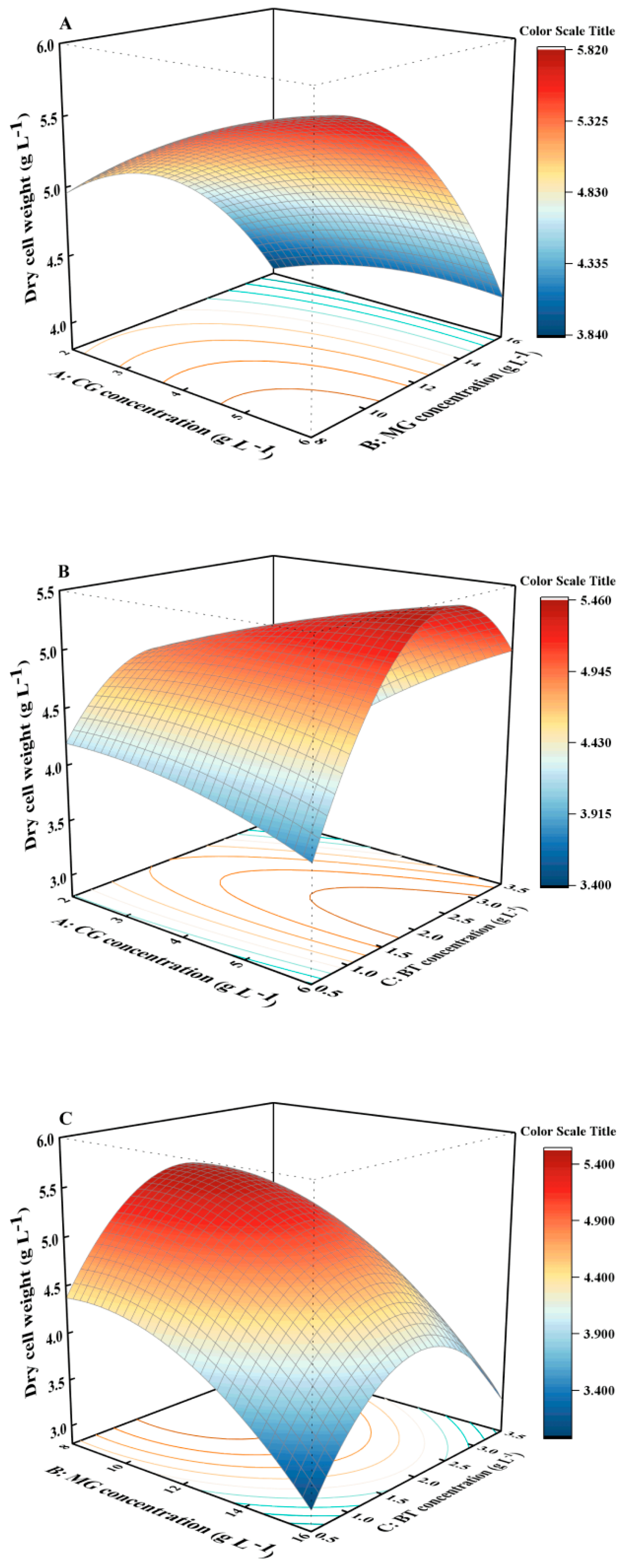
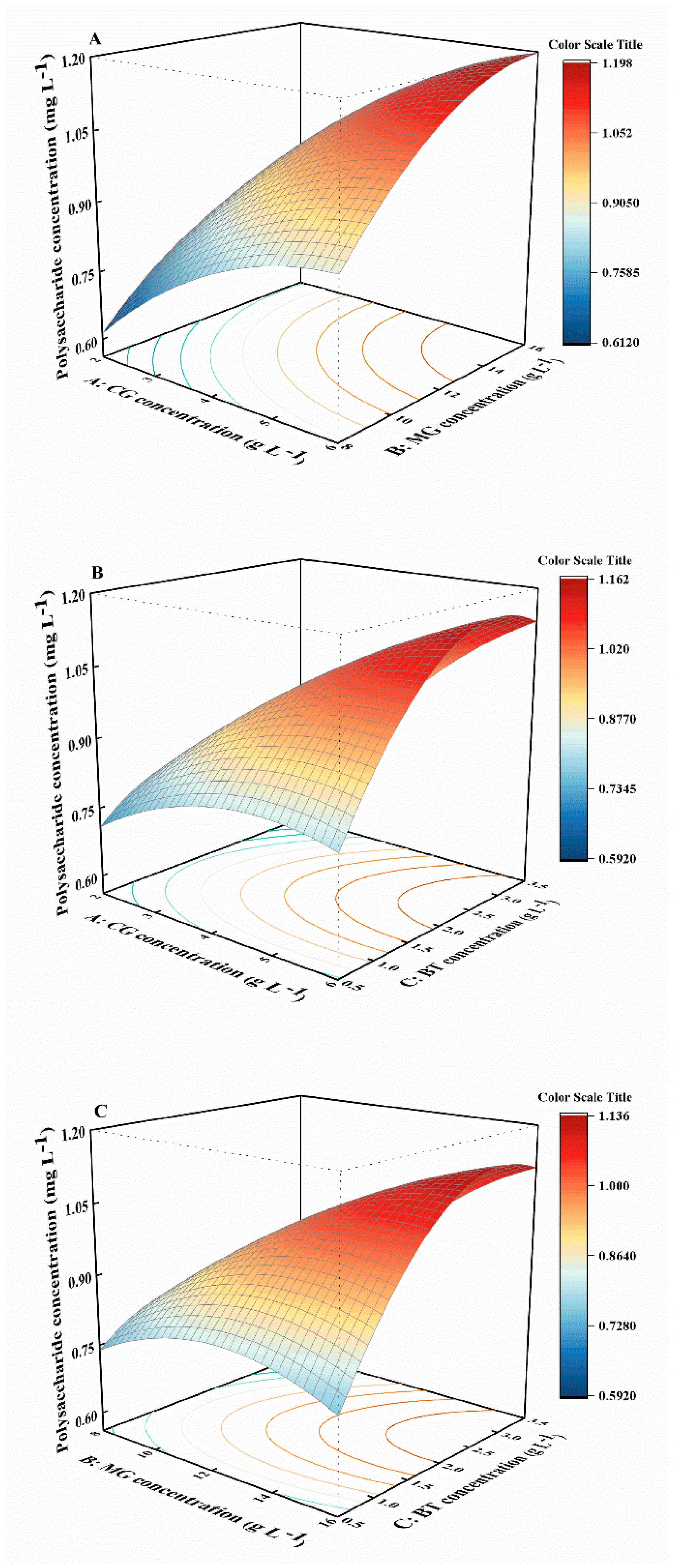
2.3. Combined Effects of Variables on Biomass, Phycoerythrin, and Polysaccharide Concentrations
2.4. Optimization and Experimental Validation
3. Discussion
4. Materials and Methods
4.1. Microalgal Strain
4.2. Experimental Design
4.2.1. Experiments to Determine the Optimal Concentrations of Exogenous Substances
4.2.2. Response Surface Experiments for Three Exogenous Substances
4.3. Dry Cell Weight
4.4. Phycoerythrin Concentration
4.5. Polysaccharide Concentration
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kathiresan, S.; Sarada, R.; Bhattacharya, S.; Ravishankar, G.A. Culture media optimization for growth and phycoerythrin production from Porphyridium purpureum. Biotechnol. Bioeng. 2007, 96, 456–463. [Google Scholar] [CrossRef]
- Juin, C.; Chérouvrier, J.-R.; Thiéry, V.; Gagez, A.-L.; Bérard, J.-B.; Joguet, N.; Kaas, R.; Cadoret, J.-P.; Picot, L. Microwave-Assisted Extraction of Phycobiliproteins from Porphyridium purpureum. Appl. Biochem. Biotechnol. 2015, 175, 1–15. [Google Scholar] [CrossRef]
- Durmaz, Y.; Monteiro, M.; Bandarra, N.; Gökpinar, Ş.; Işik, O. The effect of low temperature on fatty acid composition and tocopherols of the red microalga, Porphyridium cruentum. J. Appl. Phycol. 2007, 19, 223–227. [Google Scholar] [CrossRef]
- Lu, Q.; Li, H.; Xiao, Y.; Liu, H. A state-of-the-art review on the synthetic mechanisms, production technologies, and practical application of polyunsaturated fatty acids from microalgae. Algal Res. 2021, 55, 102281. [Google Scholar] [CrossRef]
- Arad, S.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, E.; Fedewa, E.J.; Richardson, T.L. Extraction protocols for the quantification of phycobilins in aqueous phytoplankton extracts. J. Appl. Phycol. 2011, 23, 865–871. [Google Scholar] [CrossRef]
- Borovkov, A.B.; Gudvilovich, I.N.; Lelekov, A.S.; Avsiyan, A.L. Effect of specific irradiance on productivity and pigment and protein production of Porphyridium purpureum (Rhodophyta) semi-continuous culture. Bioresour. Technol. 2023, 374, 128771. [Google Scholar] [CrossRef]
- Yabuta, Y.; Fujimura, H.; Kwak, C.S.; Enomoto, T.; Watanabe, F. Antioxidant Activity of the Phycoerythrobilin Compound Formed from a Dried Korean Purple Laver (Porphyra sp.) during in vitro Digestion. Food Sci. Technol. Res. 2010, 16, 347–352. [Google Scholar] [CrossRef]
- Wang, C.; Shen, Z.; Li, L.; Li, Y.; Zhao, H.; Jiang, X. Immunomodulatory activity of R-phycoerythrin from Porphyra haitanensis via TLR4/NF-κB-dependent immunocyte differentiation. Food Funct. 2020, 11, 2173–2185. [Google Scholar] [CrossRef]
- Cai, C.; Wang, Y.; Li, C.; Guo, Z.; Jia, R.; Wu, W.; Hu, Y.; He, P. Purification and photodynamic bioactivity of phycoerythrin and phycocyanin from Porphyra yezoensis Ueda. J. Ocean Univ. China 2014, 13, 479–484. [Google Scholar] [CrossRef]
- Simovic, A.; Combet, S.; Cirkovic Velickovic, T.; Nikolic, M.; Minic, S. Probing the stability of the food colourant R-phycoerythrin from dried Nori flakes. Food Chem. 2022, 374, 131780. [Google Scholar] [CrossRef]
- Serive, B.; Kaas, R.; Bérard, J.-B.; Pasquet, V.; Picot, L.; Cadoret, J.-P. Selection and optimisation of a method for efficient metabolites extraction from microalgae. Bioresour. Technol. 2012, 124, 311–320. [Google Scholar] [CrossRef]
- Patel, A.K.; Laroche, C.; Marcati, A.; Ursu, A.V.; Jubeau, S.; Marchal, L.; Petit, E.; Djelveh, G.; Michaud, P. Separation and fractionation of exopolysaccharides from Porphyridium cruentum. Bioresour. Technol. 2013, 145, 345–350. [Google Scholar] [CrossRef]
- Morais, M.G.; Santos, T.D.; Moraes, L.; Vaz, B.S.; Morais, E.G.; Costa, J.A.V. Exopolysaccharides from microalgae: Production in a biorefinery framework and potential applications. Bioresour. Technol. Rep. 2022, 18, 101006. [Google Scholar] [CrossRef]
- Medina-Cabrera, E.V.; Rühmann, B.; Schmid, J.; Sieber, V. Characterization and comparison of Porphyridium sordidum and Porphyridium purpureum concerning growth characteristics and polysaccharide production. Algal Res. 2020, 49, 101931. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.-K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Sosa-Hernández, J.E.; Rodas-Zuluaga, L.I.; Castillo-Zacarías, C.; Rostro-Alanís, M.; de la Cruz, R.; Carrillo-Nieves, D.; Salinas-Salazar, C.; Fuentes Grunewald, C.; Llewellyn, C.A.; Olguín, E.J.; et al. Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum. Mar. Drugs 2019, 17, 460. [Google Scholar] [CrossRef] [PubMed]
- García, A.B.; Longo, E.; Murillo, M.C.; Bermejo, R. Using a B-Phycoerythrin Extract as a Natural Colorant: Application in Milk-Based Products. Molecules 2021, 26, 297. [Google Scholar] [CrossRef] [PubMed]
- Hsieh-Lo, M.; Castillo, G.; Ochoa-Becerra, M.A.; Mojica, L. Phycocyanin and phycoerythrin: Strategies to improve production yield and chemical stability. Algal Res. 2019, 42, 101600. [Google Scholar] [CrossRef]
- Jiao, K.; Chang, J.; Zeng, X.; Ng, I.S.; Xiao, Z.; Sun, Y.; Tang, X.; Lin, L. 5-Aminolevulinic acid promotes arachidonic acid biosynthesis in the red microalga Porphyridium purpureum. Biotechnol. Biofuels 2017, 10, 168. [Google Scholar] [CrossRef]
- You, T.; Barnett, S.M. Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem. Eng. J. 2004, 19, 251–258. [Google Scholar] [CrossRef]
- Jiao, K.; Xiao, W.; Xu, Y.; Zeng, X.; Ho, S.-H.; Laws, E.A.; Lu, Y.; Ling, X.; Shi, T.; Sun, Y.; et al. Using a trait-based approach to optimize mixotrophic growth of the red microalga Porphyridium purpureum towards fatty acid production. Biotechnol. Biofuels 2018, 11, 273. [Google Scholar] [CrossRef]
- Li, S.; Ji, L.; Shi, Q.; Wu, H.; Fan, J. Advances in the production of bioactive substances from marine unicellular microalgae Porphyridium spp. Bioresour. Technol. 2019, 292, 122048. [Google Scholar] [CrossRef]
- Sánchez-Saavedra, M.d.P.; Castro-Ochoa, F.Y.; Nava-Ruiz, V.M.; Ruiz-Güereca, D.A.; Villagómez-Aranda, A.L.; Siqueiros-Vargas, F.; Molina-Cárdenas, C.A. Effects of nitrogen source and irradiance on Porphyridium cruentum. J. Appl. Phycol. 2018, 30, 783–792. [Google Scholar] [CrossRef]
- Seemashree, M.H.; Chauhan, V.S.; Sarada, R. Phytohormone supplementation mediated enhanced biomass production, lipid accumulation, and modulation of fatty acid profile in Porphyridium purpureum and Dunaliella salina cultures. Biocatal. Agric. Biotechnol. 2022, 39, 102253. [Google Scholar] [CrossRef]
- Gorain, P.C.; Bagchi, S.K.; Mallick, N. Effects of calcium, magnesium and sodium chloride in enhancing lipid accumulation in two green microalgae. Environ. Technol. 2013, 34, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.T.; Warren, M.J.; Smith, A.G. Algae need their vitamins. Eukaryot. Cell 2006, 5, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Dahuja, A.; Tiwari, S.; Punia, S.; Tak, Y.; Amarowicz, R.; Bhoite, A.G.; Singh, S.; Joshi, S.; Panesar, P.S.; et al. Recent trends in extraction of plant bioactives using green technologies: A review. Food Chem. 2021, 353, 129431. [Google Scholar] [CrossRef] [PubMed]
- Guihéneuf, F.; Stengel, D.B. Towards the biorefinery concept: Interaction of light, temperature and nitrogen for optimizing the co-production of high-value compounds in Porphyridium purpureum. Algal Res. 2015, 10, 152–163. [Google Scholar] [CrossRef]
- Yao, T.; Huang, J.; Su, B.; Wei, L.; Zhang, A.; Zhang, D.-F.; Zhou, Y.; Ma, G.-s. Enhanced phycocyanin production of Arthrospira maxima by addition of mineral elements and polypeptides using response surface methodology. Front. Mar. Sci. 2022, 9, 1057201. [Google Scholar] [CrossRef]
- Cañete-Rodríguez, A.M.; Santos-Dueñas, I.M.; Jiménez-Hornero, J.E.; Ehrenreich, A.; Liebl, W.; García-García, I. Gluconic acid: Properties, production methods and applications—An excellent opportunity for agro-industrial by-products and waste bio-valorization. Process Biochem. 2016, 51, 1891–1903. [Google Scholar] [CrossRef]
- Pang, N.; Gu, X.; Fu, X.; Chen, S. Effects of gluconate on biomass improvement and light stress tolerance of Haematococcus pluvialis in mixotrophic culture. Algal Res. 2019, 43, 101647. [Google Scholar] [CrossRef]
- Nemanja, T.; Bojanić, N.; Rakić, D.; Takači, A.; Zeković, Z.; Fišteš, A.; Bodroža-Solarov, M.; Pavlić, B. Defatted wheat germ as source of polyphenols—Optimization of microwave-assisted extraction by RSM and ANN approach. Chem. Eng. Process. 2019, 143, 107634. [Google Scholar]
- Mousavi, M.; Mehrzad, J.; Najafi, M.F.; Zhiani, R.; Shamsian, S.A.A. Nitrate and ammonia: Two key nitrogen sources for biomass and phycocyanin production by Arthrospira (Spirulina) platensis. J. Appl. Phycol. 2022, 34, 2271–2281. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Zhai, J.; Wei, H. Effect of nitrogen limitation on biochemical composition and photosynthetic performance for fed-batch mixotrophic cultivation of microalga Spirulina platensis. Bioresour. Technol. 2018, 263, 555–561. [Google Scholar] [CrossRef]
- Arumugam, M.; Agarwal, A.; Arya, M.C.; Ahmed, Z. Influence of nitrogen sources on biomass productivity of microalgae Scenedesmus bijugatus. Bioresour. Technol. 2013, 131, 246–249. [Google Scholar] [CrossRef]
- Gladfelter, M.F.; Buley, R.P.; Belfiore, A.P.; Fernandez-Figueroa, E.G.; Gerovac, B.L.; Baker, N.D.; Wilson, A.E. Dissolved nitrogen form mediates phycocyanin content in cyanobacteria. Freshw. Biol. 2022, 67, 954–964. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl. Microbiol. Biotechnol. 2008, 81, 629–636. [Google Scholar] [CrossRef]
- Ji, L.; Li, S.; Chen, C.; Jin, H.; Wu, H.; Fan, J. Physiological and transcriptome analysis elucidates the metabolic mechanism of versatile Porphyridium purpureum under nitrogen deprivation for exopolysaccharides accumulation. Bioresour. Bioprocess. 2021, 8, 73. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, J.; Liu, D.; Zeng, Y.; Tang, S.; Han, Y.; Jiang, Y.; Cai, Z. A growth-boosting synergistic mechanism of Chromochloris zofingiensis under mixotrophy. Algal Res. 2022, 66, 102812. [Google Scholar] [CrossRef]
- Grama, B.S.; Agathos, S.N.; Jeffryes, C.S. Balancing Photosynthesis and Respiration Increases Microalgal Biomass Productivity during Photoheterotrophy on Glycerol. ACS Sustain. Chem. Eng. 2016, 4, 1611–1618. [Google Scholar] [CrossRef]
- Cecchin, M.; Benfatto, S.; Griggio, F.; Mori, A.; Cazzaniga, S.; Vitulo, N.; Delledonne, M.; Ballottari, M. Molecular basis of autotrophic vs mixotrophic growth in Chlorella sorokiniana. Sci. Rep. 2018, 8, 6465. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, J.; Wu, H.; Jiang, P.; Chen, Z.; Xiang, W. Growth and Biochemical Composition of Porphyridium purpureum SCS-02 under Different Nitrogen Concentrations. Mar. Drugs 2019, 17, 124. [Google Scholar] [CrossRef] [PubMed]
- Soanen, N.; Da Silva, E.; Gardarin, C.; Michaud, P.; Laroche, C. Improvement of exopolysaccharide production by Porphyridium marinum. Bioresour. Technol. 2016, 213, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Velea, S.; Ilie, L.I. Optimization of Porphyridium purpureum culture growth using two variables experimental design: Light and sodium bicarbonate. UPB Sci. Bull. 2011, 73, 81–94. [Google Scholar]
- Razaghi, A.; Godhe, A.; Albers, E. Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Cent. Eur. J. Biol. 2014, 9, 156–162. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Liqin, S.; Wang, C.; Lei, S. Effects of Light Regime on Extracellular Polysaccharide Production by Porphyridium cruentum Cultured in Flat Plate Photobioreactors. In Proceedings of the 2nd International Conference on Bioinformatics and Biomedical Engineering, Shanghai, China, 16–18 May 2008; IEEE: Piscataway, NJ, USA, 2008; pp. 1488–1491. [Google Scholar]
- Singh, S.; Arad, S.; Richmond, A. Extracellular polysaccharide production in outdoor mass cultures of Porphyridium sp. in flat plate glass reactors. J. Appl. Phycol. 2000, 12, 269–275. [Google Scholar] [CrossRef]
- Aslan, N.; Cebeci, Y. Application of Box–Behnken design and response surface methodology for modeling of some Turkish coals. Fuel 2007, 86, 90–97. [Google Scholar] [CrossRef]
- Marjakangas, J.M.; Chen, C.-Y.; Lakaniemi, A.-M.; Puhakka, J.A.; Whang, L.-M.; Chang, J.-S. Selecting an indigenous microalgal strain for lipid production in anaerobically treated piggery wastewater. Bioresour. Technol. 2015, 191, 369–376. [Google Scholar] [CrossRef]
- Beer, S.; Eshel, A. Determining Phycoerythrin and Phycocyanin Concentrations in Aqueous Crude Extracts of Red Algae. Mar. Freshw. Res. 1985, 36, 785–792. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Modified Equations with Significant Terms | p-Value | R2 | Adj.R2 | SD | Lack of Fit | C.V.% |
|---|---|---|---|---|---|---|---|
| Biomass concentration | 5.21 + 0.28A − 0.705B + 0.0925C − 0.1475AB + 0.4875AC − 0.0225BC − 0.1187A2 − 0.4137B2 − 1.01C2 | 0.0123 | 0.8893 | 0.7469 | 0.4449 | 0.3384 | 9.93 |
| Phycoerythrin concentration | 74.11 − 1.6A − 0.85B − 8.84C − 5.89AB − 1.42AC + 0.165BC + 1.99A2 − 4.96B2 − 24.28C2 | 0.0054 | 0.9145 | 0.8046 | 6.78 | 0.6649 | 11.07 |
| Polysaccharide concentration | 1.02 + 0.1588A + 0.1338B + 0.0525C + 0.0225AB + 0.11AC + 0.125BC − 0.0647A2 − 0.0748B2 − 0.1472C2 | 0.0054 | 0.9142 | 0.8039 | 0.0915 | 2.45 | 10.32 |
| Std | Run | Variables | Responses | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CG | MG | BT | Biomass | Phycoerythrin Concentration | Polysaccharide Concentration | |||||
| (g L−1) | (g L−1) | (g L−1) | (g L−1) | (mg L−1) | (g L−1) | |||||
| Coded | Actural | Coded | Actural | Coded | Actural | Actural | Actural | Actural | ||
| 1 | 5 | −1 | 2 | −1 | 8 | 0 | 2 | 5.77 | 84.23 | 1 |
| 2 | 13 | 1 | 6 | −1 | 8 | 0 | 2 | 4.87 | 70.01 | 1.13 |
| 3 | 16 | −1 | 2 | 1 | 16 | 0 | 2 | 4.02 | 56.78 | 0.76 |
| 4 | 8 | 1 | 6 | 1 | 16 | 0 | 2 | 4.36 | 36.91 | 0.64 |
| 5 | 6 | −1 | 2 | 0 | 12 | −1 | 0.5 | 5.28 | 69.73 | 0.52 |
| 6 | 17 | 1 | 6 | 0 | 12 | −1 | 0.5 | 4.22 | 56.78 | 0.78 |
| 7 | 9 | −1 | 2 | 0 | 12 | 1 | 3.5 | 4.91 | 44.05 | 1.06 |
| 8 | 7 | 1 | 6 | 0 | 12 | 1 | 3.5 | 3.78 | 60.77 | 1.29 |
| 9 | 3 | 0 | 4 | −1 | 8 | −1 | 0.5 | 3.32 | 43.79 | 0.64 |
| 10 | 14 | 0 | 4 | 1 | 16 | −1 | 0.5 | 3.5 | 33.3 | 1.09 |
| 11 | 4 | 0 | 4 | −1 | 8 | 1 | 3.5 | 5.31 | 65.09 | 1.03 |
| 12 | 10 | 0 | 4 | 1 | 16 | 1 | 3.5 | 5.33 | 72.4 | 1.02 |
| 13 | 2 | 0 | 4 | 0 | 12 | 0 | 2 | 6.08 | 72.02 | 0.91 |
| 14 | 15 | 0 | 4 | 0 | 12 | 0 | 2 | 3.25 | 52.51 | 0.71 |
| 15 | 12 | 0 | 4 | 0 | 12 | 0 | 2 | 4.77 | 78.83 | 0.93 |
| 16 | 11 | 0 | 4 | 0 | 12 | 0 | 2 | 3.57 | 82.06 | 0.81 |
| 17 | 1 | 0 | 4 | 0 | 12 | 0 | 2 | 3.86 | 62.7 | 0.76 |
| Number | Medium | Special Culture Conditions | Culturing Scale | Biomass Concentration or Cell Number | Polysaccharide Concentration (g L−1) | PB and PE Concentrations (mg L−1) | Refs |
|---|---|---|---|---|---|---|---|
| 1 | F/2 | N: P ratio | 250 mL flask | 5.94 × 109 cell L−1 | 0.23 | NA | [46] |
| 2 | F/2-RSE | Light, temperature, and nitrogen | 250 mL glass flask | 3.4 g L−1 | 0.92 | PB: 47.20 PE: 38.80 | [29] |
| 3 | OMII | Consumption of N and P | 30 L flat-plate photobioreactor | 1.71 × 1010 cell L−1 | 0.73 | NA | [47] |
| 4 | OMI | Light regime | 15 L plate photobioreactor | 1.38 × 1010 cell L−1 | 0.95 | NA | [48] |
| 5 | ASW | Outdoor mass culture | 72 L flat plate glass reactor | 1.37 × 1011 cell L−1 | 1.32 | NA | [49] |
| 6 | ASW | Different nitrogen concentrations | 6 × 60 cm photobioreactor | 5.53 g L−1 | 2.14 | PB: 1010 | [43] |
| 7 | Pm | Light, temperature and nitrogen | 5 L photobioreactor | 6.12 × 1010 cell L−1 | 4.10 | NA | [44] |
| 8 | ASW | Optimization of light and sodium bicarbonate | 3 L batch culture of photobioreactor | 15.2 g L−1 | 4.5 | PB 12.17 g/100 g | [45] |
| 9 | ASW | light intensities | BioIII fermenter | 4.44 × 109 cell L−1 | 4.63 | NA | [21] |
| 10 | ASW | Addition of CG, MG, and BT | 500 mL flask | 5.97 g L−1 | 1.42 | 102.95 |
| Parameters | Lable | Coded Levels and Concentrations | ||
|---|---|---|---|---|
| (g L−1) | −1 | 0 | +1 | |
| CG | A | 2 | 4 | 6 |
| MG | B | 8 | 12 | 16 |
| BT | C | 0.5 | 2 | 3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, S.; Zhang, A.-H.; Huang, J.; Yao, T.; Feng, B.; Zhou, X.; Hu, Y.; Pan, M. Maximizing Polysaccharides and Phycoerythrin in Porphyridium purpureum via the Addition of Exogenous Compounds: A Response-Surface-Methodology Approach. Mar. Drugs 2024, 22, 138. https://doi.org/10.3390/md22030138
Yi S, Zhang A-H, Huang J, Yao T, Feng B, Zhou X, Hu Y, Pan M. Maximizing Polysaccharides and Phycoerythrin in Porphyridium purpureum via the Addition of Exogenous Compounds: A Response-Surface-Methodology Approach. Marine Drugs. 2024; 22(3):138. https://doi.org/10.3390/md22030138
Chicago/Turabian StyleYi, Sanjiong, Ai-Hua Zhang, Jianke Huang, Ting Yao, Bo Feng, Xinghu Zhou, Yadong Hu, and Mingxuan Pan. 2024. "Maximizing Polysaccharides and Phycoerythrin in Porphyridium purpureum via the Addition of Exogenous Compounds: A Response-Surface-Methodology Approach" Marine Drugs 22, no. 3: 138. https://doi.org/10.3390/md22030138