
Synthetic Antimicrobial Peptides Fail to Induce Leucocyte Innate Immune Functions but Elicit Opposing Transcriptomic Profiles in European Sea Bass and Gilthead Seabream


Abstract
:1. Introduction
2. Results
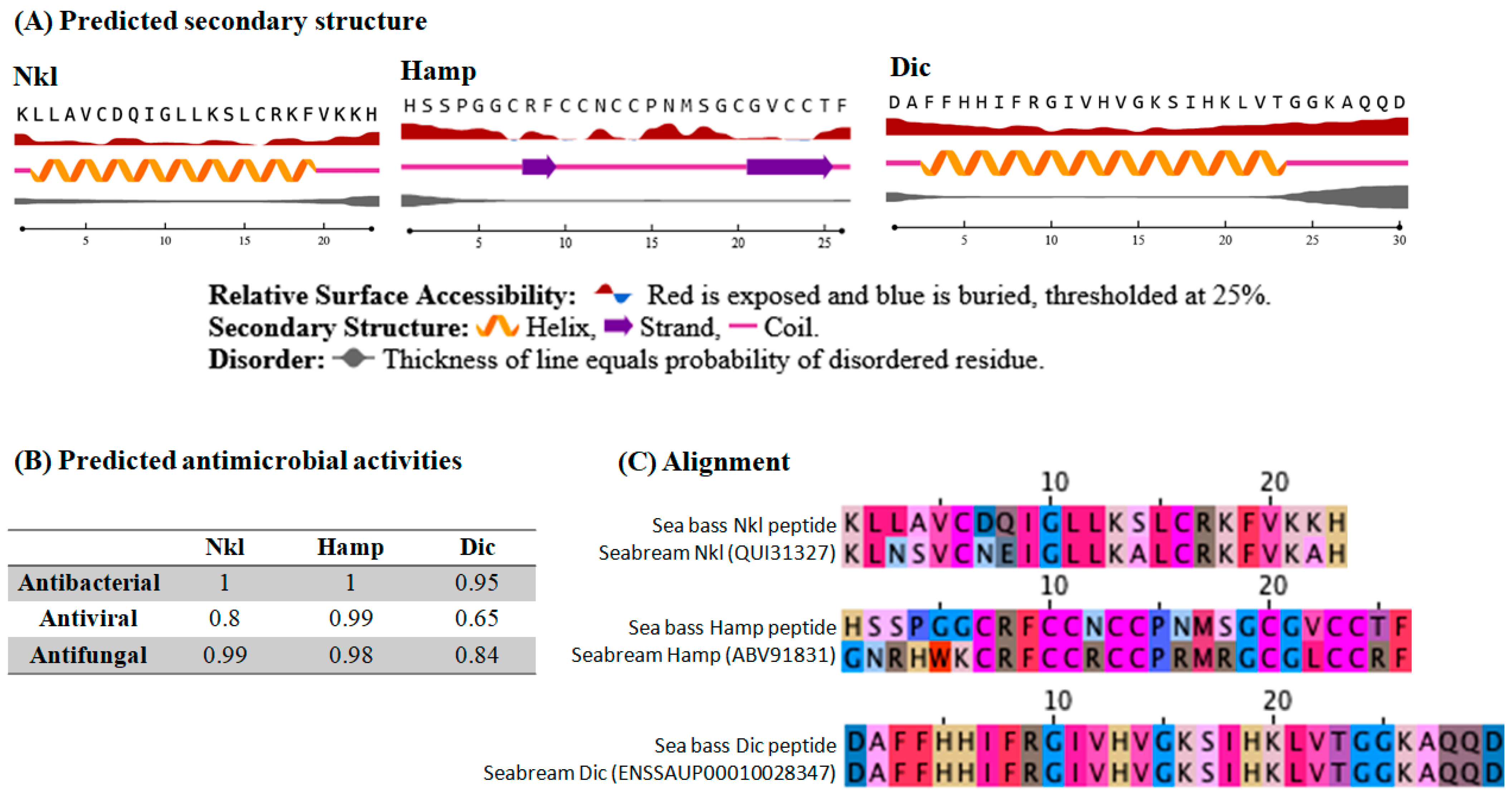
2.1. In Silico Predictions Show Homology between European Sea Bass and Gilthead Seabream Putative Peptides
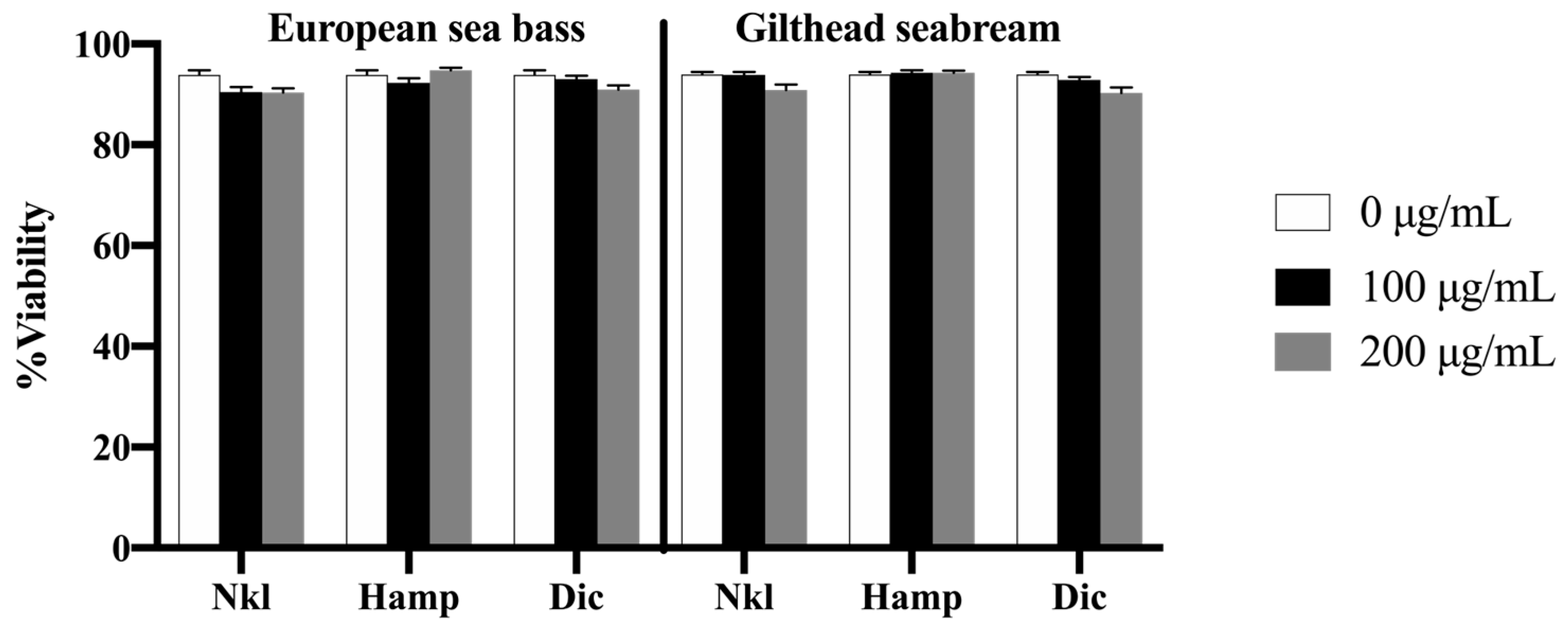
2.2. Nkl, Hamp, and Dic Synthetic Peptides Were Not Toxic for HK Cells
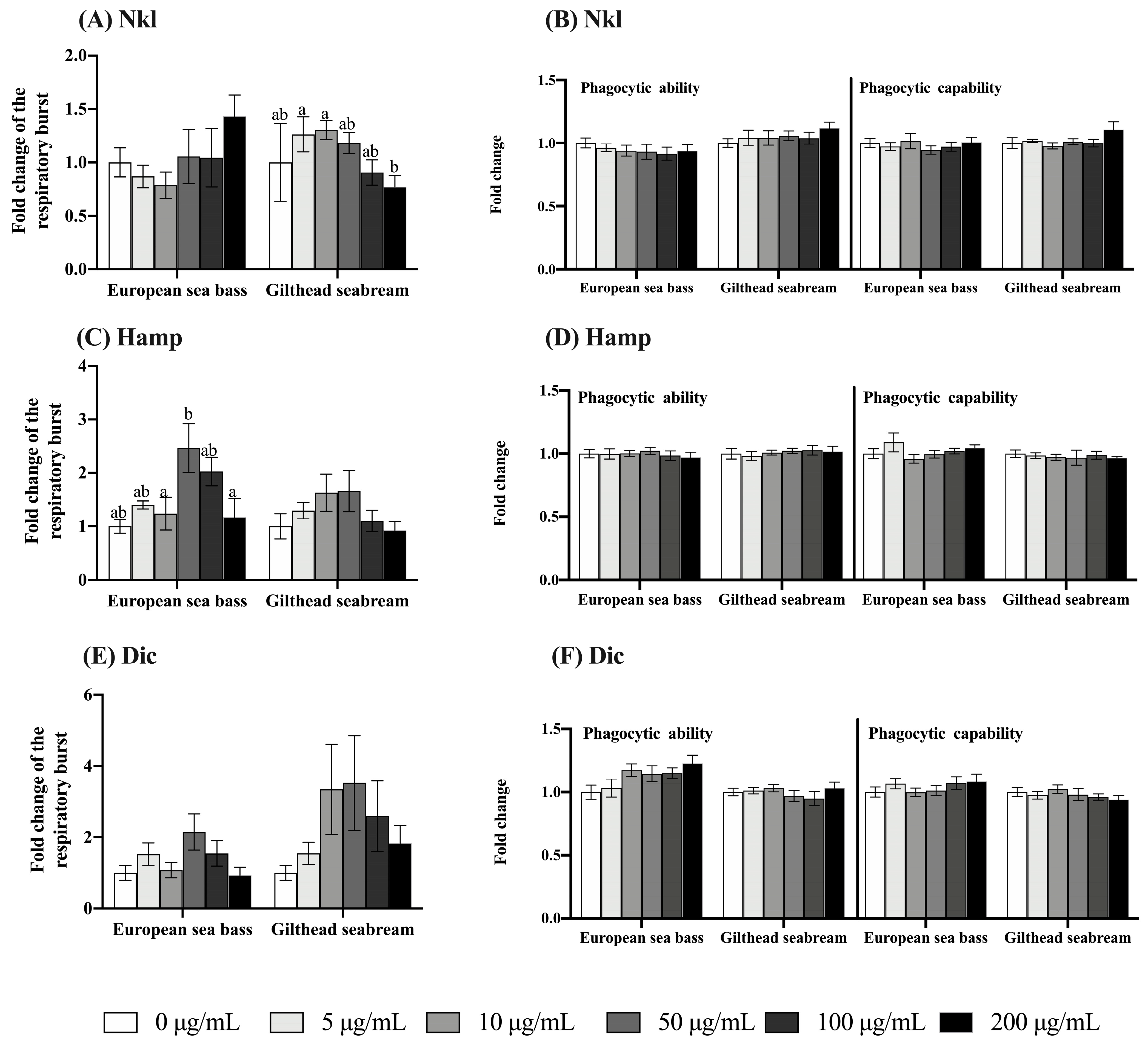
2.3. Nkl, Hamp, and Dic Synthetic Peptides Failed to Modulate Innate Immune Activities
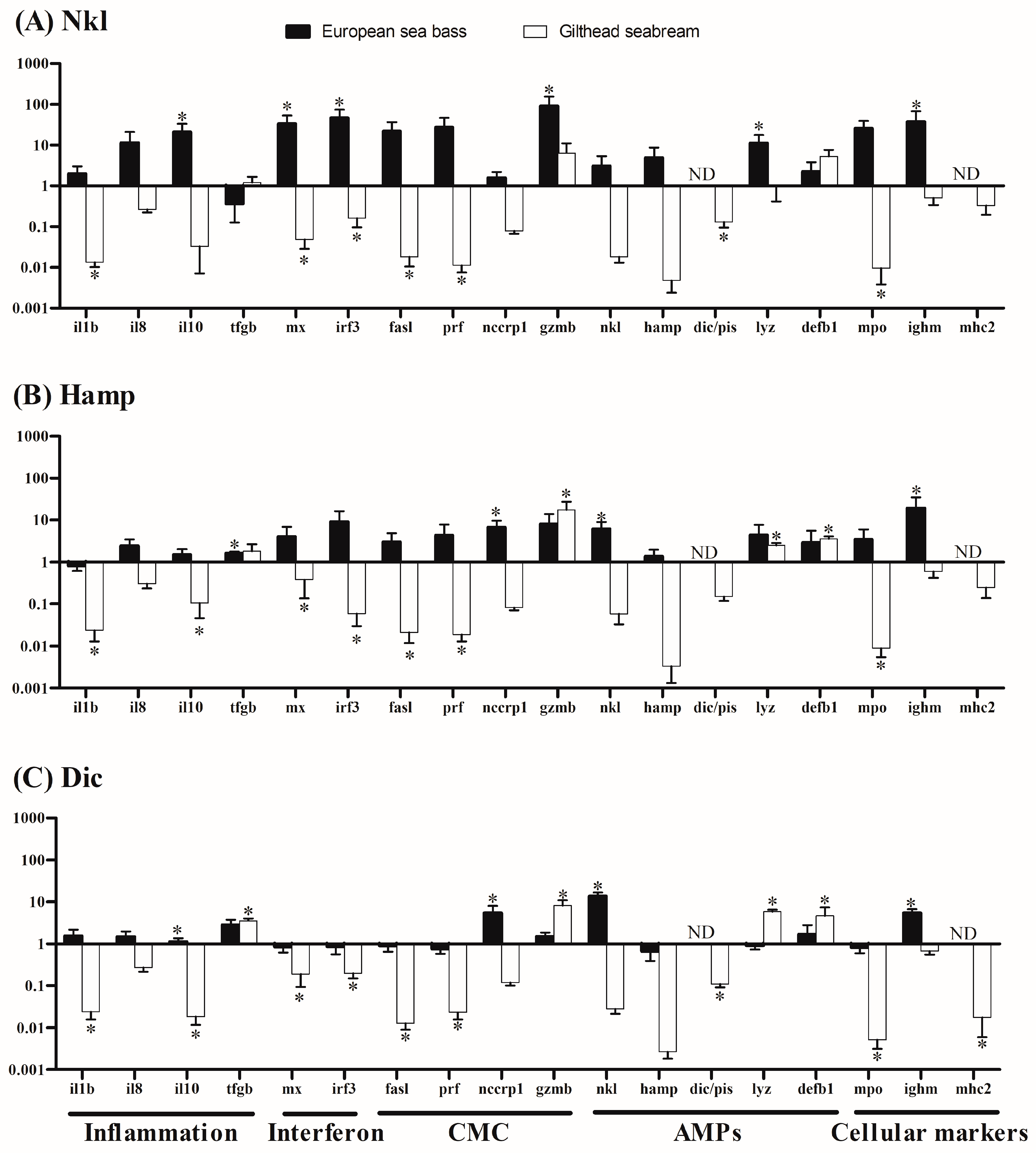
2.4. Nkl, Hamp, or Dic Synthetic Peptides Induced Species-Opposed Transcriptional Profiles
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Peptide Design and Production
4.3. Sampling and Head-Kidney Cell Isolation
4.4. HK Cell Stimulation with AMPs
4.5. Viability of Head-Kidney Cells
4.6. Respiratory Burst Activity
4.7. Phagocytosis
4.8. Gene Expression Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cabello, F.C.; Godfrey, H.P.; Buschmann, A.H.; Dölz, H.J. Aquaculture as yet Another Environmental Gateway to the Development and Globalisation of Antimicrobial Resistance. Lancet Infect. Dis. 2016, 16, e127–e133. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Jiang, C. Antimicrobial Peptides: Structure, Mechanism, and Modification. Eur. J. Med. Chem. 2023, 255, 115377. [Google Scholar] [CrossRef]
- Katzenback, B.A. Antimicrobial Peptides as Mediators of Innate Immunity in Teleosts. Biology 2015, 4, 607–639. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; Chaves-Pozo, E.; Meseguer, J.; Esteban, M.A.; Cuesta, A. Biological Role of Fish Antimicrobial Peptides. In Antimicrobial Peptides; Centro Oceanográfico de Murcia: Murcia, Spain, 2013; pp. 31–60. [Google Scholar]
- Bahar, A.A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef]
- Schuerholz, T.; Brandenburg, K.; Marx, G. Antimicrobial Peptides and Their Potential Application in Inflammation and Sepsis. Crit. Care 2012, 16, 207. [Google Scholar] [CrossRef] [PubMed]
- Ghodsi, Z.; Kalbassi, M.R.; Farzaneh, P.; Mobarez, A.M.; Beemelmanns, C.; Amiri Moghaddam, J. Immunomodulatory Function of Antimicrobial Peptide EC-Hepcidin1 Modulates the Induction of Inflammatory Gene Expression in Primary Cells of Caspian Trout (Salmo trutta Caspius Kessler, 1877). Fish Shellfish Immunol. 2020, 104, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.C.; Pan, C.Y.; Chen, J.Y. Tilapia Hepcidin (TH)2-3 as a Transgene in Transgenic Fish Enhances Resistance to Vibrio Vulnificus Infection and Causes Variations in Immune-Related Genes after Infection by Different Bacterial Species. Fish Shellfish Immunol. 2010, 29, 430–439. [Google Scholar] [CrossRef]
- Huang, X.; Hu, B.; Yang, X.; Gong, L.; Tan, J.; Deng, L. The Putative Mature Peptide of Piscidin-1 Modulates Global Transcriptional Profile and Proliferation of Splenic Lymphocytes in Orange-Spotted Grouper (Epinephelus coioides). Fish Shellfish Immunol. 2019, 86, 1035–1043. [Google Scholar] [CrossRef]
- Lin, W.C.; Chang, H.Y.; Chen, J.Y. Electrotransfer of the Tilapia Piscidin 3 and Tilapia Piscidin 4 Genes into Skeletal Muscle Enhances the Antibacterial and Immunomodulatory Functions of Oreochromis niloticus. Fish Shellfish Immunol. 2016, 50, 200–209. [Google Scholar] [CrossRef]
- Cervera, L.; González-Fernández, C.; Cano, D.; Esteban, M.Á.; Mercado, L.; Chaves-Pozo, E.; Cuesta, A. Immunity Elicited by AMP-Encoding Plasmids Fails to Increase the Protection of European Sea Bass against Nodavirus. Fish Shellfish Immunol. 2023, 132, 108507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Long, H.; Sun, L. A NK-Lysin from Cynoglossus Semilaevis Enhances Antimicrobial Defense against Bacterial and Viral Pathogens. Dev. Comp. Immunol. 2013, 40, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.F.; Li, C.H.; Chen, J. Molecular Characterization of the NK-Lysin in a Teleost Fish: Antimicrobial Activity and Immunomodulatory Activity on Monocytes/Macrophages. Fish Shellfish Immunol. 2019, 92, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yan, X.; Wang, G.; Liu, W.; Wang, Y.; Hao, D.; Liu, H.; Zhang, M. NKHs27, a Sevenband Grouper NK-Lysin Peptide That Possesses Immunoregulatory and Antimicrobial Activity. Fish Shellfish Immunol. 2023, 136, 108715. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; González-Fernández, C.; Cárdenas, C.; Guzmán, F.; León, R.; Cuesta, A. NK-Lysin Peptides Ameliorate Viral Encephalopathy and Retinopathy Disease Signs and Provide Partial Protection against Nodavirus Infection in European Sea Bass. Antivir. Res. 2021, 192, 105104. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhang, H.; Li, Y.; Wang, G.; Tang, B.; Zhao, J.; Huang, Y.; Zheng, J. Cathelicidin-Derived Antimicrobial Peptides Inhibit Zika Virus through Direct Inactivation and Interferon Pathway. Front. Immunol. 2018, 9, 722. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Mas, V.; Tafalla, C.; Perez, L.; Coll, J.M.; Estepa, A. Dual Antiviral Activity of Human Alpha-Defensin-1 against Viral Haemorrhagic Septicaemia Rhabdovirus (VHSV): Inactivation of Virus Particles and Induction of a Type I Interferon-Related Response. Antiviral Res. 2007, 76, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Chico, V.; Marroquí, L.; Perez, L.; Coll, J.M.; Estepa, A. Expression and Antiviral Activity of a β-Defensin-like Peptide Identified in the Rainbow Trout (Oncorhynchus mykiss) EST Sequences. Mol. Immunol. 2008, 45, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.Y.; Peng, K.C.; Lin, C.H.; Chen, J.Y. Transgenic Expression of Tilapia Hepcidin 1-5 and Shrimp Chelonianin in Zebrafish and Their Resistance to Bacterial Pathogens. Fish Shellfish Immunol. 2011, 31, 275–285. [Google Scholar] [CrossRef]
- Brogden, K.A.; Heidari, M.; Sacco, R.E.; Palmquist, D.; Guthmiller, J.M.; Johnson, G.K.; Jia, H.P.; Tack, B.F.; McCray, P.B. Defensin-Induced Adaptive Immunity in Mice and Its Potential in Preventing Periodontal Disease. Oral Microbiol. Immunol. 2003, 18, 95–99. [Google Scholar] [CrossRef]
- Ma, J.Y.; Shao, S.; Wang, G. Antimicrobial Peptides: Bridging Innate and Adaptive Immunity in the Pathogenesis of Psoriasis. Chin. Med. J. 2020, 133, 2966–2975. [Google Scholar] [CrossRef]
- Mookherjee, N.; Brown, K.L.; Bowdish, D.M.E.; Doria, S.; Falsafi, R.; Hokamp, K.; Roche, F.M.; Mu, R.; Doho, G.H.; Pistolic, J.; et al. Modulation of the TLR-Mediated Inflammatory Response by the Endogenous Human Host Defense Peptide LL-37. J. Immunol. 2006, 176, 2455–2464. [Google Scholar] [CrossRef]
- Ting, C.H.; Pan, C.Y.; Chen, Y.C.; Lin, Y.C.; Chen, T.Y.; Rajanbabu, V.; Chen, J.Y. Impact of Tilapia Hepcidin 2-3 Dietary Supplementation on the Gut Microbiota Profile and Immunomodulation in the Grouper (Epinephelus lanceolatus). Sci. Rep. 2019, 9, 19047. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The Immunology of Host Defence Peptides: Beyond Antimicrobial Activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef]
- Kang, H.K.; Kim, C.; Seo, C.H.; Park, Y. The Therapeutic Applications of Antimicrobial Peptides (AMPs): A Patent Review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Q.; Niu, J.; Tang, J.; Wang, B.; Abarike, E.D.; Lu, Y.; Cai, J.; Jian, J. NK-Lysin from Oreochromis niloticus Improves Antimicrobial Defence against Bacterial Pathogens. Fish Shellfish Immunol. 2018, 72, 259–265. [Google Scholar] [CrossRef]
- Zhang, M.; Li, M.F.; Sun, L. NKLP27: A Teleost NK-Lysin Peptide That Modulates Immune Response, Induces Degradation of Bacterial DNA, and Inhibits Bacterial and Viral Infection. PLoS ONE 2014, 9, e0106543. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; Chaves-Pozo, E.; Cuesta, A. NK-Lysin Is Highly Conserved in European Sea Bass and Gilthead Seabream but Differentially Modulated during the Immune Response. Fish Shellfish Immunol. 2020, 99, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Shen, M.; Wang, L.; Wang, C.; Gao, M.; Yu, G.; Chang, Z.; Zhang, X. Antibacterial and Immunoregulatory Activity of an Antimicrobial Peptide Hepcidin in Loach (Misgurnus anguillicaudatus). Int. J. Biol. Macromol. 2023, 24, 124833. [Google Scholar] [CrossRef] [PubMed]
- Neves, J.V.; Caldas, C.; Vieira, I.; Ramos, M.F.; Rodrigues, P.N.S. Multiple Hepcidins in a Teleost Fish, Dicentrarchus labrax : Different Hepcidins for Different Roles. J. Immunol. 2015, 195, 2696–2709. [Google Scholar] [CrossRef] [PubMed]
- León, R.; Ruiz, M.; Valero, Y.; Cárdenas, C.; Guzman, F.; Vila, M.; Cuesta, A. Exploring Small Cationic Peptides of Different Origin as Potential Antimicrobial Agents in Aquaculture. Fish Shellfish Immunol. 2020, 98, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Erdem Büyükkiraz, M.; Kesmen, Z. Antimicrobial Peptides (AMPs): A Promising Class of Antimicrobial Compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Habibullah-Al-Mamun, M.; Nagano, I.; Masunaga, S.; Kitazawa, D.; Matsuda, H. Antibiotics, Antibiotic-Resistant Bacteria, and Resistance Genes in Aquaculture: Risks, Current Concern, and Future Thinking. Environ. Sci. Pollut. Res. 2022, 29, 11054–11075. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.; Roa, F.; González-Chavarría, I.; Astuya, A.; Maura, R.; Montesino, R.; Muñoz, C.; Camacho, F.; Saavedra, P.; Valenzuela, A.; et al. In Vitro Immunomodulatory Activities of Peptides Derived from Salmo salar NK-Lysin and Cathelicidin in Fish Cells. Fish Shellfish Immunol. 2019, 88, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.A.; Santana, P.A.; Salinas-Parra, N.; Beltrán, D.; Guzmán, F.; Vega, B.; Acosta, F.; Mercado, L. Immune Modulation Ability of Hepcidin from Teleost Fish. Animals 2022, 12, 1586. [Google Scholar] [CrossRef] [PubMed]
- Chia, T.J.; Wu, Y.C.; Chen, J.Y.; Chi, S.C. Antimicrobial Peptides (AMP) with Antiviral Activity against Fish Nodavirus. Fish Shellfish Immunol. 2010, 28, 434–439. [Google Scholar] [CrossRef]
- Valero, Y.; Arizcun, M.; Cortés, J.; Ramírez-Cepeda, F.; Guzmán, F.; Mercado, L.; Esteban, M.Á.; Chaves-Pozo, E.; Cuesta, A. NK-Lysin, Dicentracin and Hepcidin Antimicrobial Peptides in European Sea Bass. Ontogenetic Development and Modulation in Juveniles by Nodavirus. Dev. Comp. Immunol. 2020, 103, 103516. [Google Scholar] [CrossRef] [PubMed]
- Cervera, L.; González-Fernández, C.; Arizcun, M.; Cuesta, A.; Chaves-Pozo, E. Severe Natural Outbreak of Cryptocaryon Irritans in Gilthead Seabream Produces Leukocyte Mobilization and Innate Immunity at the Gill Tissue. Int. J. Mol. Sci. 2022, 23, 937. [Google Scholar] [CrossRef] [PubMed]
- Rakers, S.; Niklasson, L.; Steinhagen, D.; Kruse, C.; Schauber, J.; Sundell, K.; Paus, R. Antimicrobial Peptides (AMPs) from Fish Epidermis: Perspectives for Investigative Dermatology. J. Investig. Dermatol. 2013, 133, 1140–1149. [Google Scholar] [CrossRef]
- Buonocore, F.; Picchietti, S.; Porcelli, F.; Della Pelle, G.; Olivieri, C.; Poerio, E.; Bugli, F.; Menchinelli, G.; Sanguinetti, M.; Bresciani, A.; et al. Fish-Derived Antimicrobial Peptides: Activity of a Chionodracine Mutant against Bacterial Models and Human Bacterial Pathogens. Dev. Comp. Immunol. 2019, 96, 9–17. [Google Scholar] [CrossRef]
- Asensio-Calavia, P.; González-Acosta, S.; Otazo-Pérez, A.; López, M.R.; Morales-delaNuez, A.; Pérez de la Lastra, J.M. Teleost Piscidins—In Silico Perspective of Natural Peptide Antibiotics from Marine Sources. Antibiotics 2023, 12, 855. [Google Scholar] [CrossRef]
- García-Beltrán, J.M.; Arizcun, M.; Chaves-Pozo, E. Antimicrobial Peptides from Photosynthetic Marine Organisms with Potential Application in Aquaculture. Mar. Drugs 2023, 21, 290. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.A.; Acosta, F.; Montero, D.; Guzmán, F.; Torres, E.; Vega, B.; Mercado, L. Synthetic Hepcidin from Fish: Uptake and Protection against Vibrio anguillarum in Sea Bass (Dicentrarchus labrax). Fish Shellfish Immunol. 2016, 55, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Juretić, D.; Simunić, J. Design of α-Helical Antimicrobial Peptides with a High Selectivity Index. Expert Opin. Drug Discov. 2019, 14, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Freitas, E.D.; Bataglioli, A.; Oshodi, J.; Beppu, M.M. Antimicrobial Peptides and Their Potential Application in Antiviral Coating Agents. Colloids Surf. B Biointerfaces 2020, 217, 112693. [Google Scholar] [CrossRef]
- Personne, H.; Paschoud, T.; Fulgencio, S.; Baeriswyl, S.; Köhler, T.; Van Delden, C.; Stocker, A.; Javor, S.; Reymond, J.L. To Fold or Not to Fold: Diastereomeric Optimization of an α-Helical Antimicrobial Peptide. J. Med. Chem. 2023, 66, 7570–7583. [Google Scholar] [CrossRef]
- Lin, W.; Liu, S.; Hu, L.; Zhang, S. Characterization and Bioactivity of Hepcidin-2 in Zebrafish: Dependence of Antibacterial Activity upon Disulfide Bridges. Peptides 2014, 57, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Hocquellet, A.; Le Senechal, C.; Garbay, B. Importance of the Disulfide Bridges in the Antibacterial Activity of Human Hepcidin. Peptides 2012, 36, 303–307. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M.Á. The Antimicrobial Peptide Hepcidin Exerts an Important Role in the Innate Immunity against Bacteria in the Bony Fish Gilthead Seabream. Mol. Immunol. 2008, 45, 2333–2342. [Google Scholar] [CrossRef]
- Neves, J.V.; Barroso, C.; Carvalho, P.; Nunes, M.; Gonçalves, J.F.M.; Rodrigues, P.N.S. Characterization of Erythroferrone in a Teleost Fish (Dicentrarchus labrax) with Two Functional Hepcidin Types: More Than an Erythroid Regulator. Front. Immunol. 2022, 13, 867630. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2019, 27, 4–16. [Google Scholar] [CrossRef]
- Chen, J.; Lv, Y.P.; Dai, Q.M.; Hu, Z.H.; Liu, Z.M.; Li, J.H. Host Defense Peptide LEAP-2 Contributes to Monocyte/Macrophage Polarization in Barbel Steed (Hemibarbus labeo). Fish Shellfish Immunol. 2019, 87, 184–192. [Google Scholar] [CrossRef]
- Hao, D.; Wang, G.; Liu, N.; Liu, H.; Wang, C.; Liu, W.; Yan, X.; Zhang, M. Antimicrobial and Immunoregulatory Activities of TS40, a Derived Peptide of a TFPI-2 Homologue from Black Rockfish (Sebastes schlegelii). Mar. Drugs 2022, 20, 353. [Google Scholar] [CrossRef]
- Chen, R.Y.; Chen, J.; Liu, Z.M.; Lin, Z.H.; Guo, Z.P. Barbel Steed (Hemibarbus labeo) NK-Lysin Protects against Aeromonas Hydrophila Infection via Immunomodulatory Activity. Dev. Comp. Immunol. 2021, 122, 104114. [Google Scholar] [CrossRef]
- Valero, Y.; Saraiva-Fraga, M.; Costas, B.; Guardiola, F.A. Antimicrobial Peptides from Fish: Beyond the Fight against Pathogens. Rev. Aquac. 2020, 12, 224–253. [Google Scholar] [CrossRef]
- Poisa-Beiro, L.; Dios, S.; Montes, A.; Aranguren, R.; Figueras, A.; Novoa, B. Nodavirus Increases the Expression of Mx and Inflammatory Cytokines in Fish Brain. Mol. Immunol. 2008, 45, 218–225. [Google Scholar] [CrossRef]
- Carla Piazzon, M.; Lutfall, G.; Forlenzaa, M. IL10, a Tale of an Evolutionarily Conserved Cytokine across Vertebrates. Crit. Rev. Immunol. 2016, 36, 99–129. [Google Scholar] [CrossRef]
- Neves, J.V.; Caldas, C.; Wilson, J.M.; Rodrigues, P.N.S. Molecular Mechanisms of Hepcidin Regulation in Sea Bass (Dicentrarchus labrax). Fish Shellfish Immunol. 2011, 31, 1154–1161. [Google Scholar] [CrossRef]
- Masso-Silva, J.A.; Diamond, G. Antimicrobial Peptides from Fish. Pharmaceuticals 2014, 7, 265–310. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M.C. Priming of the Neutrophil Respiratory Burst: Role in Host Defense and Inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Zhang, P.; Zhang, Q.; Zhang, J. Two Hepcidins from Spotted Scat (Scatophagus argus) Possess Antibacterial and Antiviral Functions In Vitro. Fish Shellfish Immunol. 2016, 50, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; García-Alcázar, A.; Esteban, M.Á.; Cuesta, A.; Chaves-Pozo, E. Antimicrobial Response Is Increased in the Testis of European Sea Bass, but Not in Gilthead Seabream, upon Nodavirus Infection. Fish Shellfish Immunol. 2015, 44, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.D.; Kung, C.W.; Chen, J.Y. Antiviral Activity by Fish Antimicrobial Peptides of Epinecidin-1 and Hepcidin 1-5 against Nervous Necrosis Virus in Medaka. Peptides 2010, 31, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.D.; Kung, C.W.; Chi, S.C.; Chen, J.Y. Inactivation of Nervous Necrosis Virus Infecting Grouper (Epinephelus coioides) by Epinecidin-1 and Hepcidin 1-5 Antimicrobial Peptides, and Downregulation of Mx2 and Mx3 Gene Expressions. Fish Shellfish Immunol. 2010, 28, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wei, J.; Huang, X.; Huang, Y.; Qin, Q. Antiviral Effects of β-Defensin Derived from Orange-Spotted Grouper (Epinephelus coioides). Fish Shellfish Immunol. 2012, 32, 828–838. [Google Scholar] [CrossRef]
- Falco, A.; Medina-Gali, R.M.; Antonio Poveda, J.; Bello-Perez, M.; Novoa, B.; Antonio Encinar, J. Antiviral Activity of a Turbot (Scophthalmus maximus) NK-Lysin Peptide by Inhibition of Low-PH Virus-Induced Membrane Fusion. Mar. Drugs 2019, 17, 87. [Google Scholar] [CrossRef]
- Hao, D.-F.; Wang, G.-H.; Li, N.-Q.; Liu, H.-M.; Wang, C.-B.; Liu, W.-Q.; Yan, X.; Zhang, M. Antimicrobial and Immunoregulatory Activities of the Derived Peptide of a Natural Killer Lysin from Black Rockfish (Sebastes schlegelii). Fish Shellfish Immunol. 2022, 123, 369–380. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A.; Cuesta, A. Nodavirus Infection Induces a Great Innate Cell-Mediated Cytotoxic Activity in Resistant, Gilthead Seabream, and Susceptible, European Sea Bass, Teleost Fish. Fish Shellfish Immunol. 2012, 33, 1159–1166. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Dabad, M.; Valero, Y.; Cuesta, A.; Esteban, M.Á.; Gómez-Garrido, J.; Meseguer, J.; Alioto, T.; Esteve-Codina, A. Innate Cell-Mediated Cytotoxic Activity of European Sea Bass Leucocytes against Nodavirus-Infected Cells: A Functional and RNA-Seq Study. Sci. Rep. 2017, 7, 15396. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Valero, Y.; Lozano, M.T.; Rodríguez-Cerezo, P.; Miao, L.; Campo, V.; Esteban, M.A.; Cuesta, A. Fish Granzyme a Shows a Greater Role than Granzyme b in Fish Innate Cell-Mediated Cytotoxicity. Front. Immunol. 2019, 10, 2579. [Google Scholar] [CrossRef]
- Valero, Y.; Boughlala, B.; Arizcun, M.; Patel, S.; Fiksdal, I.U.; Esteban, M.Á.; De Juan, J.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. Genes Related to Cell-Mediated Cytotoxicity and Interferon Response Are Induced in the Retina of European Sea Bass upon Intravitreal Infection with Nodavirus. Fish Shellfish Immunol. 2018, 74, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Avila, E.E. Functions of Antimicrobial Peptides in Vertebrates. Curr. Protein Pept. Sci. 2016, 18, 1098–1119. [Google Scholar] [CrossRef] [PubMed]
- Toubanaki, D.K.; Efstathiou, A.; Karagouni, E. Transcriptomic Analysis of Fish Hosts Responses to Nervous Necrosis Virus. Pathogens 2022, 11, 201. [Google Scholar] [CrossRef] [PubMed]
- García-Álvarez, M.Á.; Arizcun, M.; Chaves-Pozo, E.; Cuesta, A. Profile of Innate Immunity in Gilthead Seabream Larvae Reflects Mortality upon Betanodavirus Reassortant Infection and Replication. Int. J. Mol. Sci. 2022, 23, 5092. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; Arizcun, M.; Esteban, M.Á.; Cuesta, A.; Chaves-Pozo, E. Transcription of Histones H1 and H2B Is Regulated by Several Immune Stimuli in Gilthead Seabream and European Sea Bass. Fish Shellfish Immunol. 2016, 57, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, A.; Esteban, M.A.; Meseguer, J. Natural Cytotoxic Activity of Gilthead Seabream (Sparus aurata L.) Leucocytes Assessment by Flow Cytometry and Microscopy. Vet. Immunol. Immunopathol. 1999, 71, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Bayne, C.J.; Levy, S. Modulation of the Oxidative Burst in Trout Myeloid Cells by Adrenocorticotropic Hormone and Catecholamines: Mechanisms of Action. J. Leukoc. Biol. 1991, 50, 554–560. [Google Scholar] [CrossRef]
- Rodríguez, A.; Esteban, M.Á.; Meseguer, J. Phagocytosis and Peroxidase Release by Seabream (Sparus aurata L.) Leucocytes in Response to Yeast Cells. Anat. Rec. Part A 2003, 272, 415–423. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession Number | Sequence |
|---|---|---|
| NK-lysin | A0A218MG56 | KLLAVCDQIGLLKSLCRKFVKKH |
| Hepcidin 2.1 | KJ890397.1 | HSSPGGCRFCCNCCPNMSGCGVCCTF |
| Dicentracin | P59906 | DAFFHHIFRGIVHVGKSIHKLVTGGKAQQD |
| Protein Name | Gene Name | Accession Number | Sequence (5’→3’) | |
|---|---|---|---|---|
| Housekeeping | Elongation factor 1 alpha | ef1a | AJ866727 | F: CGTTGGCTTCAACATCAAGA R: GAAGTTGTCTGCTCCCTTGG |
| Antiviral response | Interferon-induced GTP-binding protein Mx | mx | AM228977 | F: GTATGAGGAGAAGGTGCGTCC R: CTCTTCCCCGAGCTTTGGTC |
| Interferon regulatory factor | irf3 | CBN81356 | F: AGAGGTGAGTGGCAATGGTC R: GAGCAGTTTGAAGCCTTTGG | |
| Leucocyte-type markers | Myeloperoxidase | mpo | DLAgn_0011834 | F: GAAGAGTGGGGCCTTTGTTT R: CTGGGCCTCAGTGAAGACTC |
| Immunoglobulin M heavy chain | ighm | FN908858 | F: AGGACAGGACTGCTGCTGTT R: CACCTGCTGTCTGCTGTTGT | |
| Major histocompatibility complex 2 | mhc2 | AM113466 | F: CAGAGACGGACAGGAAG R: CAAGATCAGACCCAGGA | |
| Inflammation-related molecules | Interleukin-1 beta | il1b | AJ269472 | F: CAGGACTCCGGTTTGAACAT R: GTCCATTCAAAAGGGGACAA |
| Interleukin-8 | il8 | AM490063 | F: GTCTGAGAAGCCTGGGAGTG R: GCAATGGGAGTTAGCAGGAA | |
| Interleukin-10 | il10 | DQ821114.1 | F: ACTCCTCGGTCTCTTCTCCT R: TCCACAAAACGACAGCACTG | |
| Transforming growth factor beta | tgfb | XM_051399565.1 | F: GCTACCATGCCAACTACTGC R: TGTTGCCTGCCCACATAGTA | |
| Cytotoxic response | Fas ligand | fasl | ENSDLAT00005004342 | F: GATGTGGGAGGAACCTGTGG R: GAACGGGTAGCTCTGGTCAC |
| Perforin | prf | KY801204 | F: CTGTACAACGGGCTTCTGGT R: ACTGGAGAACGTTGGACCAC | |
| Non-specific cytotoxic cell receptor protein 1 | nccrp1 | FM022070 | F: TGGGGTGAGATACGTCCACT R: TGGTTTTGGTTGCTCTGACA | |
| Granzyme B | gzmb | DLAgn_00151210 | F: AAGTTGAGCTCCAAGGCAAA R: TCCCCAGCCAGAGATGATAC | |
| Antimicrobial peptides | NK-lysin | nkl | KY801205 | F: GAAGAAACACCTCGGGGAAT R: GCAGGTCCAACATCTCCTTC |
| Hepcidin | hamp | DQ131605 | F: CCAGTCACTGAGGTGCAAGA R: GCTGTGACGCTTGTGTCTGT | |
| Dicentracin | dic | AY303949 | F: GGCAAGTCCATCCACAAACT R: ATATTGCTCCGCTTGCTGAT | |
| Lysozyme | lyz | KJ433681.1 | F: ATTTCCTGGCTGGAACACAG R: GAGCTCTGGCAACAACATCA | |
| Defensin beta 1 | defb1 | DLAgn_00041270 | F: CCTTTCCTTGGTCTTGCCCA R: ACACACAGCACAAGAAGCCT |
| Protein Name | Gene Name | Accession Number | Sequence (5’→3’) | |
|---|---|---|---|---|
| Housekeeping | Elongation factor 1 alpha | ef1a | AF184170 | F: CTTCAACGCTCAGGTCATCAT R: GCACAGCGAAACGACAAGGGGA |
| Antiviral response | Interferon-induced GTP-binding protein Mx | mx | FJ490556, FJ490555 | F: AAGAGGAGGACGAGGAGGAG R: CATCCCAGATCCTGGTCAGT |
| Interferon regulatory factor | irf3 | AM956899 | F: TCAGAATGCCCCAAGAGATT R: AGAGTCTCCGCCTTCAGATG | |
| Leucocyte-type markers | Myeloperoxidase | mpo | FM148574 | F: TTGGTCCAGACATCCTCG R: ATGGGCAAAGCGGTAG |
| Immunoglobulin M heavy chain | ighm | JQ811851 | F: CAACATGCCCAATTGATGAG R: GGCACGACACTCTAGCTTCC | |
| Major histocompatibility complex 2 | mhc2 | DQ019401 | F: CTGGACCAAGAACGGAAAGA R: CATCCCAGATCCTGGTCAGT | |
| Inflammation-related molecules | Interleukin-1 beta | il1b | AJ277166 | F: GGGCTGAACAACAGCACTCTC R: TTAACACTCTCCACCCTCCA |
| Interleukin-8 | il8 | AM765841 | F: GCCACTCTGAAGAGGACAGG R: TTTGGTTGTCTTTGGTCGAA | |
| Interleukin-10 | il10 | FG261948 | F: AGGCAGGAGTTTGAAGCTGA R: ATGCTGAAGTTGGTGGAAGG | |
| Transforming growth factor beta | tgfb | AF424703 | F: GCATGTGGCAGAGATGAAGA R: TTCAGCATGATACGGCAGAG | |
| Cytotoxic response | Fas ligand | fasl | ENSSAUT00010022087 | F: GCCACTTTGCCCGAACAATT R: GGTGGGGCAGTAAATCACCA |
| Perforin | prf | XM_030407187 | F: CTGGAGAAAGGCCTGTGGAG R: TCGGGCAACAGTCTTGGTTT | |
| Non-specific cytotoxic cell receptor protein 1 | nccrp1 | AY651258 | F: ACTTCCTGCACCGACTCAAG R: TAGGAGCTGGTTTTGGTTGG | |
| Granzyme B | gzmb | AM957224 | F: GAAACAAAGGAACGGGTCAA R: GAGCTGTCCATCTTTTGCTTG | |
| Antimicrobial peptides | NK-lysin | nkl | MN240490 | F: CGCACCTCGGAGAACTGATT R: TCCACGTCGCTTCGGTAAAA |
| Hepcidin | hamp | CB184616 | F: GCCATCGTGCTCACCTTTAT R: CTGTTGCCATACCCCATCTT | |
| Piscidin | pis | XM_030405214.1 | F: GTGGCAACCGCAATAACACA R: AATGTTTGGCTGCAATGCGT | |
| Lysozyme | lyz | AM749959 | F: CCAGGGCTGGAAATCAACTA R: CCAACATCAACACCTGCAAC | |
| Defensin beta 1 | defb1 | FM158209 | F: CCCCAGTCTGAGTGGAGTGT R: AATGAGACACGCAGCACAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cervera, L.; Chaves-Pozo, E.; Cuesta, A. Synthetic Antimicrobial Peptides Fail to Induce Leucocyte Innate Immune Functions but Elicit Opposing Transcriptomic Profiles in European Sea Bass and Gilthead Seabream. Mar. Drugs 2024, 22, 86. https://doi.org/10.3390/md22020086
Cervera L, Chaves-Pozo E, Cuesta A. Synthetic Antimicrobial Peptides Fail to Induce Leucocyte Innate Immune Functions but Elicit Opposing Transcriptomic Profiles in European Sea Bass and Gilthead Seabream. Marine Drugs. 2024; 22(2):86. https://doi.org/10.3390/md22020086
Chicago/Turabian StyleCervera, Laura, Elena Chaves-Pozo, and Alberto Cuesta. 2024. "Synthetic Antimicrobial Peptides Fail to Induce Leucocyte Innate Immune Functions but Elicit Opposing Transcriptomic Profiles in European Sea Bass and Gilthead Seabream" Marine Drugs 22, no. 2: 86. https://doi.org/10.3390/md22020086