
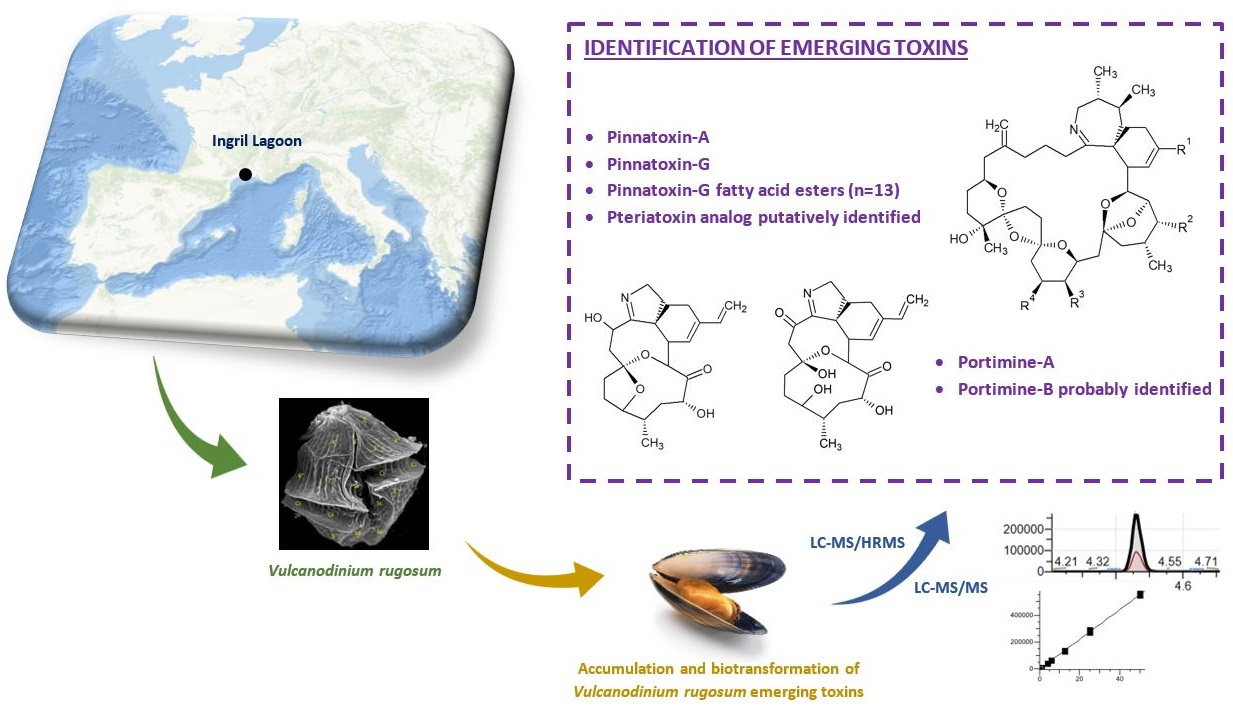
Exploration of Vulcanodinium rugosum Toxins and their Metabolism Products in Mussels from the Ingril Lagoon Hotspot in France


Abstract
:
1. Introduction
2. Results
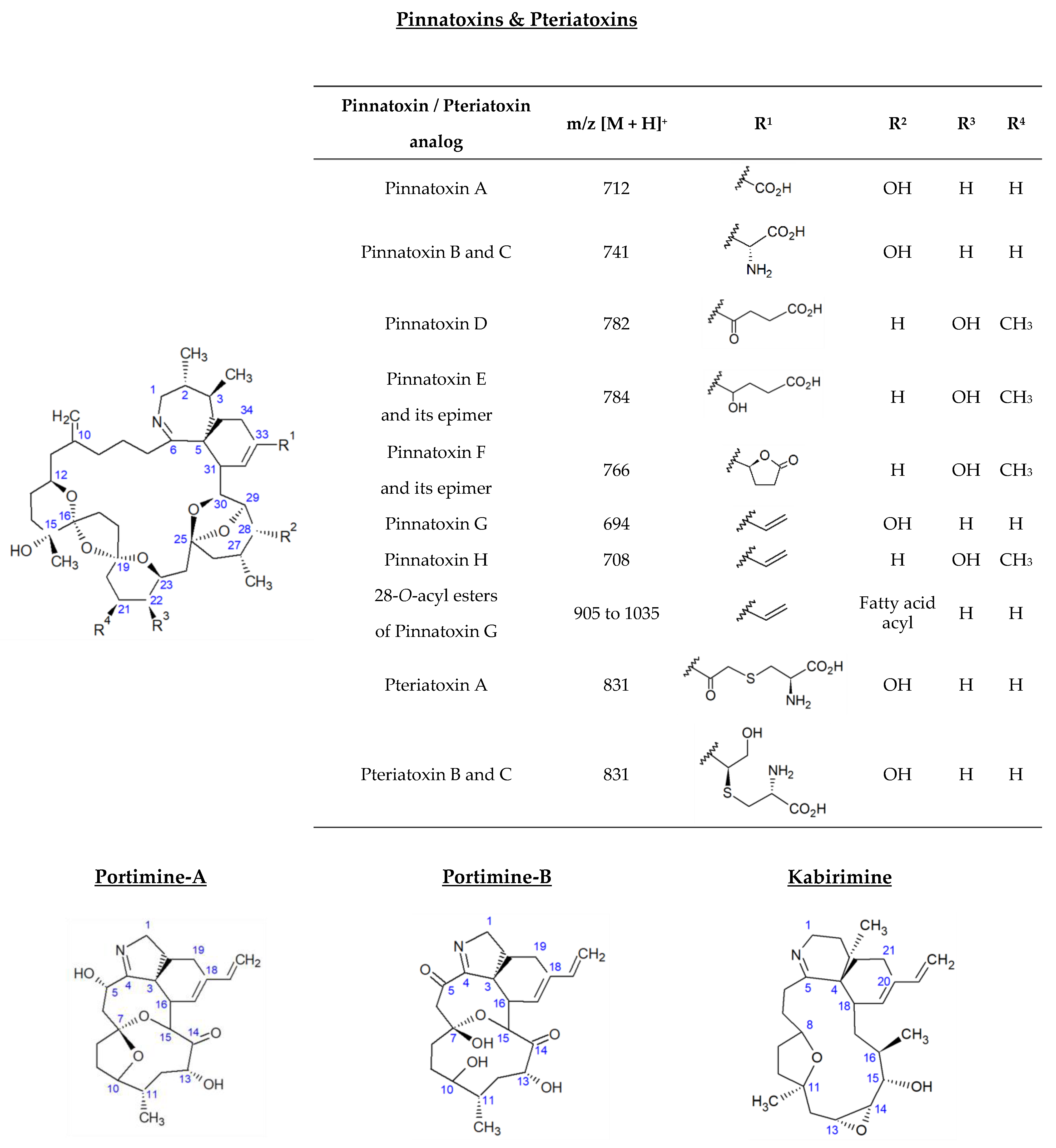
2.1. Pinnatoxins
2.1.1. LC-HRMS Screening of Pinnatoxin Analogs (Instrumental Method A.1)
2.1.2. LC-MS/MS Detection and Quantification of Pinnatoxins (Instrument Method B.1)
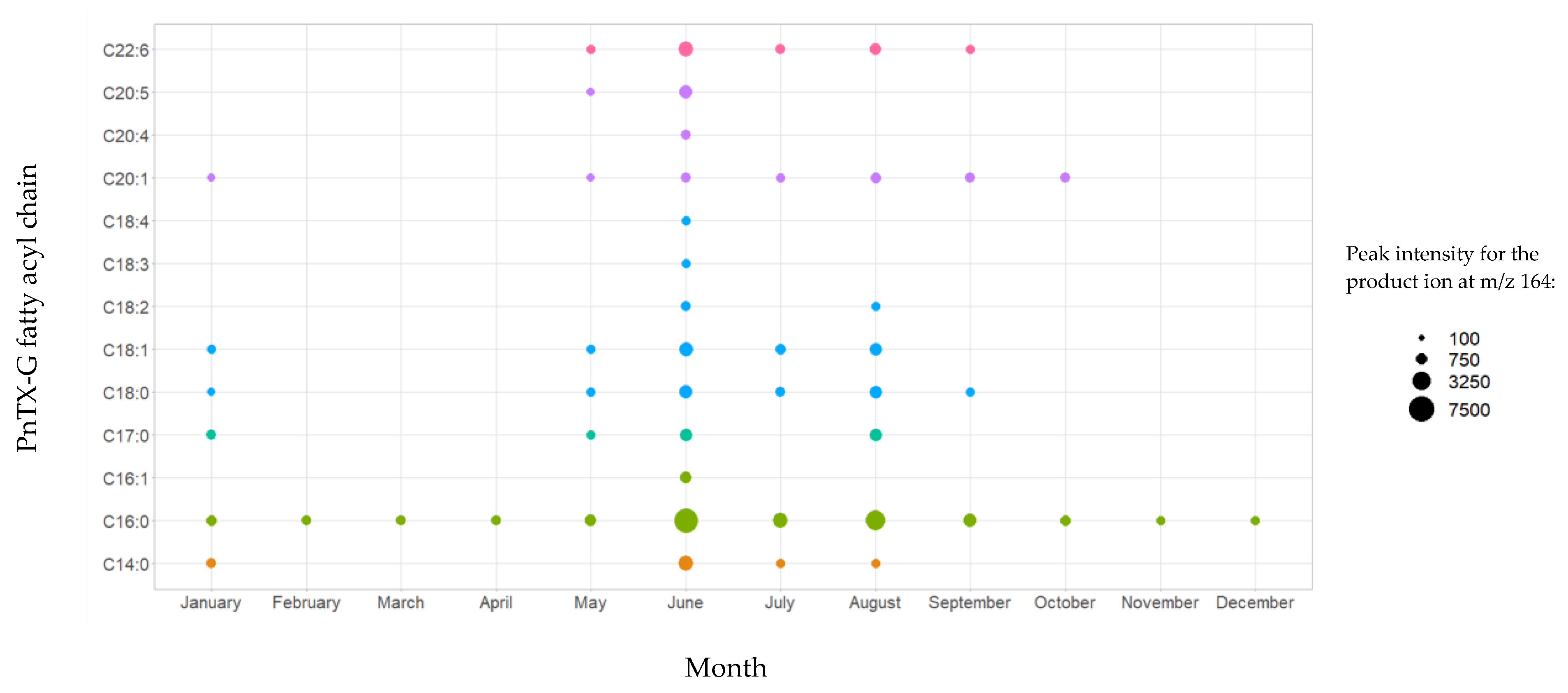
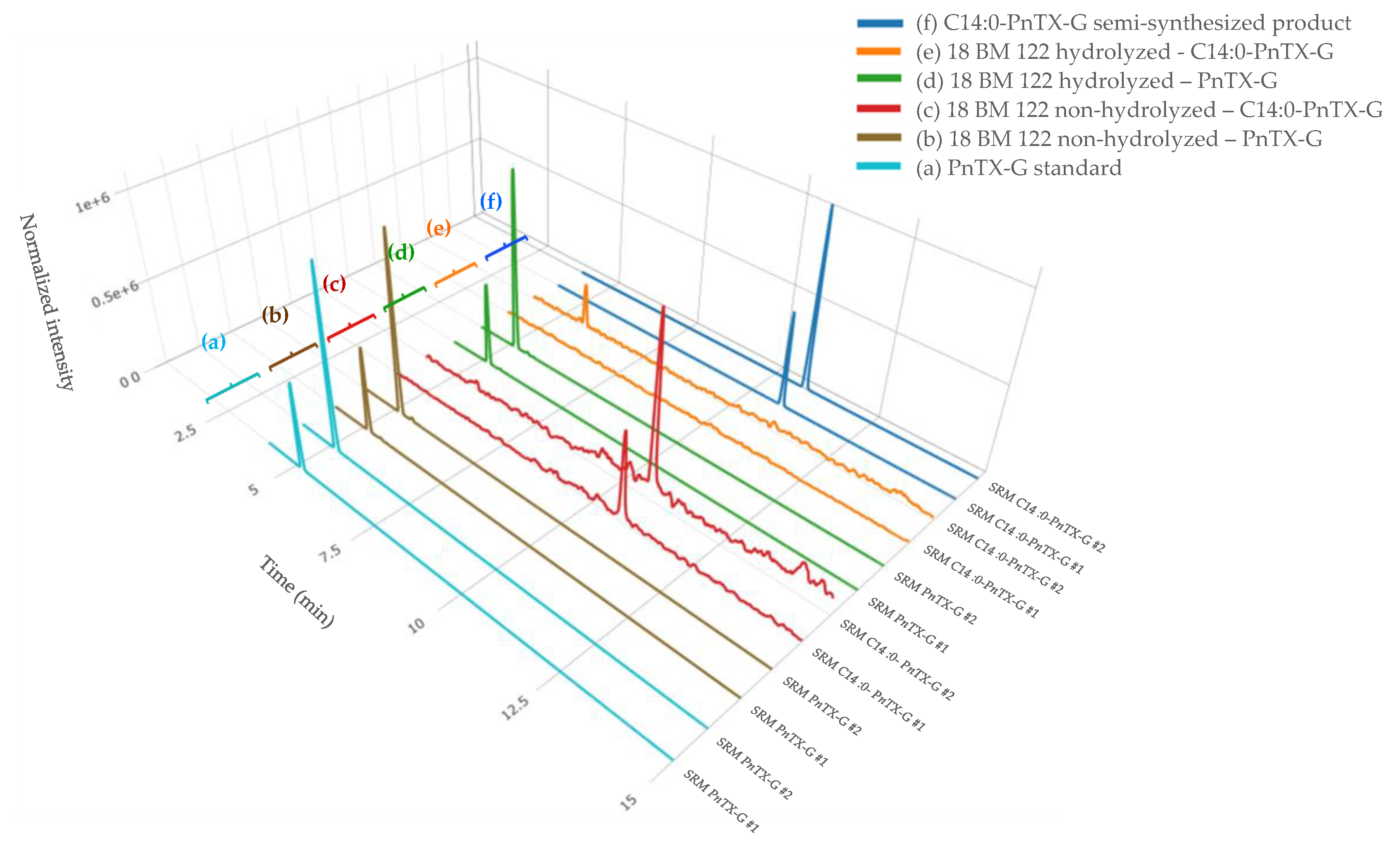
2.1.3. LC-MS/MS Identification of Fatty Acid Esters of PnTX-G (Instrumental Method B.2)
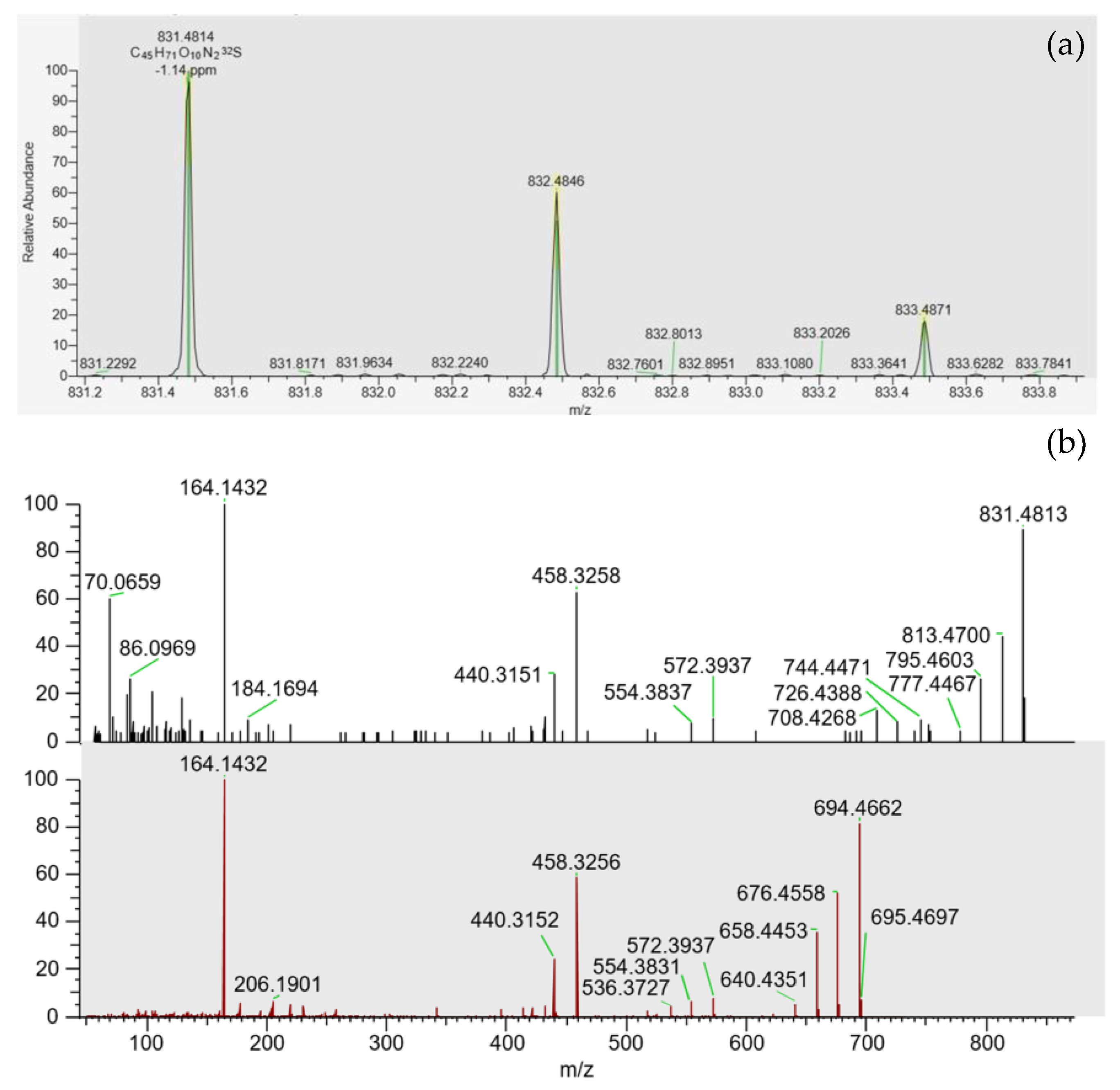
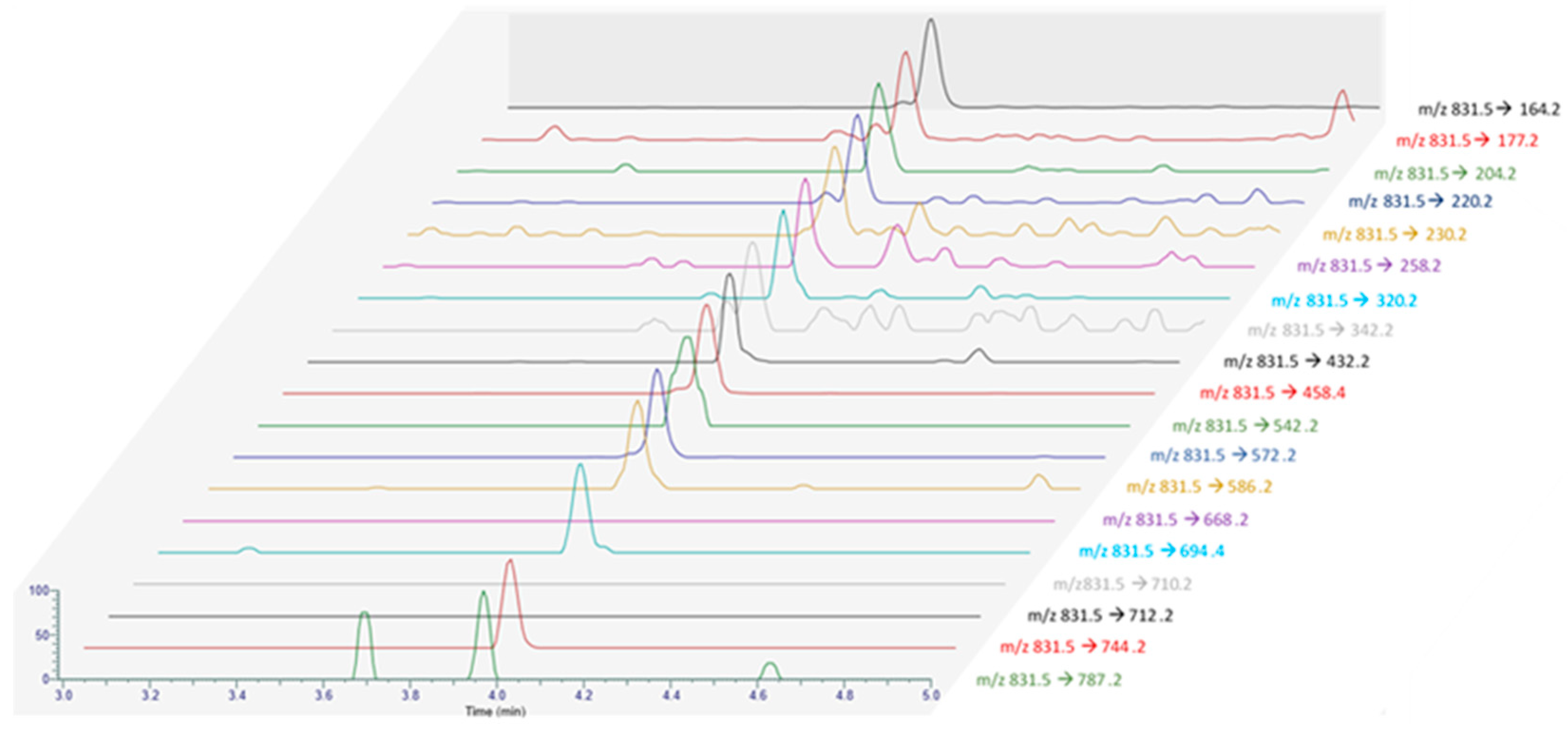
2.2. Pteriatoxins
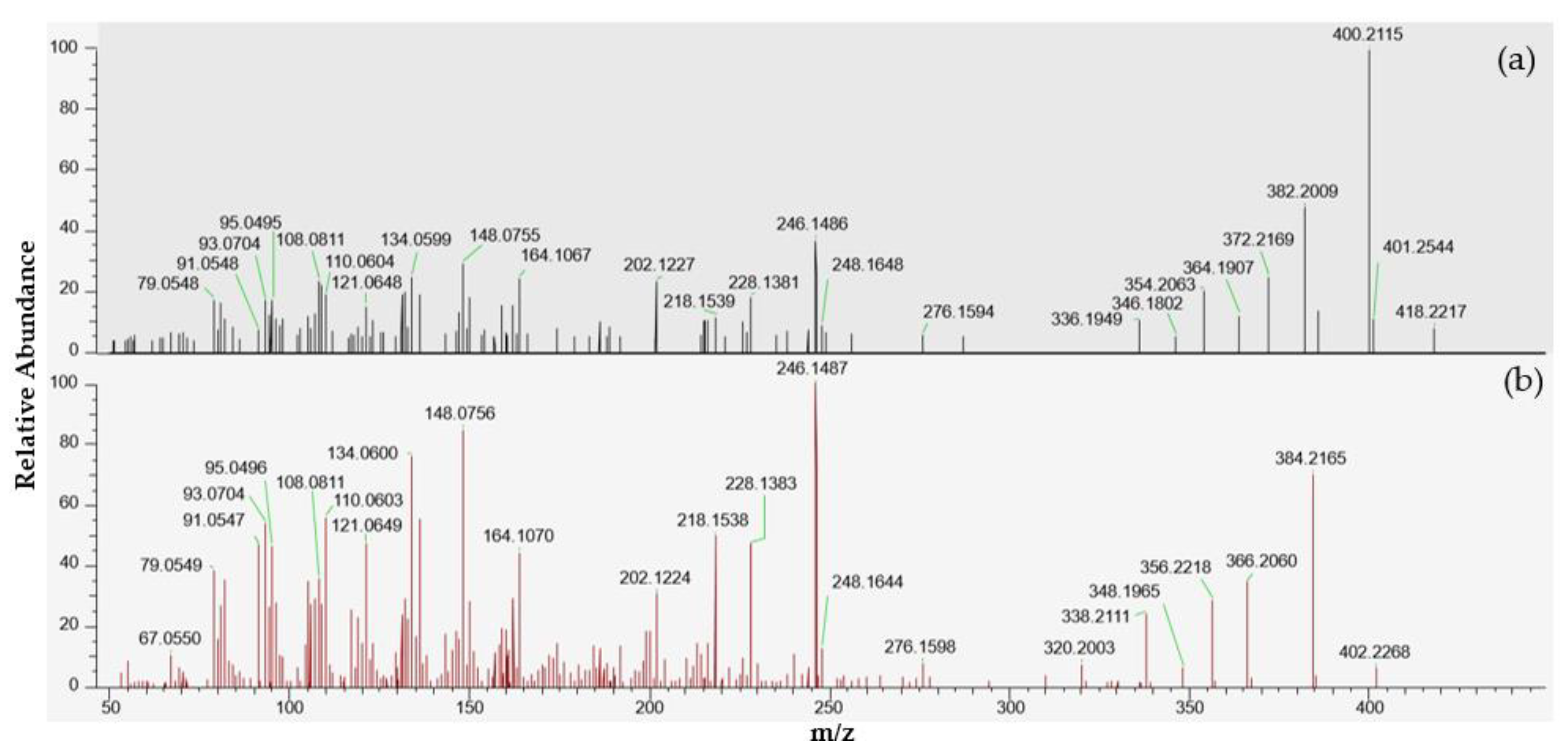
2.3. Portimines and Kabirimine
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sampling Design
4.3. Sample Preparation
4.3.1. Extraction of Toxins
4.3.2. Hydrolysis of Fatty Acid Esters of PnTX-G
4.3.3. Solid Phase Extraction Clean-Up and Concentration
4.4. Analysis by Liquid Chromatography Coupled to Mass Spectrometry
4.4.1. LC-MS/HRMS Analysis
4.4.2. LC-MS/MS Analysis
4.5. Synthesis of Fatty Acid Esters of Pinnatoxin-G
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nézan, E.; Siano, R.; Boulben, S.; Six, C.; Bilien, G.; Chèze, K.; Duval, A.; Le Panse, S.; Quéré, J.; Chomérat, N.; et al. Genetic Diversity of the Harmful Family Kareniaceae (Gymnodiniales, Dinophyceae) in France, with the Description of Karlodinium Gentienii Sp. Nov.: A New Potentially Toxic Dinoflagellate. Harmful Algae 2014, 40, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Huang, F.; Chen, S.; Tan, X.; Zuo, J.; Peng, J.; Xie, R. The Isolation and Bioactivities of Pinnatoxin. Chin. J. Mar. Drugs 1990, 9, 33–35. [Google Scholar]
- Uemura, D.; Chou, T.; Haino, T.; Nagatsu, A.; Fukuzawa, S.; Zheng, S.; Chen, H. Pinnatoxin A: A Toxic Amphoteric Macrocycle from the Okinawan Bivalve Pinna Muricata. J. Am. Chem. Soc. 1995, 117, 1155–1156. [Google Scholar] [CrossRef]
- Chou, T.; Osamu, K.; Uemura, D. Relative Stereochemistry of Pinnatoxin A, a Potent Shellfish Poison from Pinna Muricata. Tetrahedron Lett. 1996, 37, 4023–4026. [Google Scholar] [CrossRef]
- Chou, T.; Haino, T.; Kuramoto, M.; Uemura, D. Isolation and Structure of Pinnatoxin D, a New Shellfish Poison from the Okinawan Bivalve Pinna Muricata. Tetrahedron Lett. 1996, 37, 4027–4030. [Google Scholar] [CrossRef]
- McCauley, J.A.; Nagasawa, K.; Lander, P.A.; Mischke, S.G.; Semones, M.A.; Kishi, Y. Total Synthesis of Pinnatoxin A. J. Am. Chem. Soc. 1998, 120, 7647–7648. [Google Scholar] [CrossRef]
- Takada, N.; Umemura, N.; Suenaga, K.; Uemura, D. Structural Determination of Pteriatoxins A, B and C, Extremely Potent Toxins from the Bivalve Pteria Penguin. Tetrahedron Lett. 2001, 42, 3495–3497. [Google Scholar] [CrossRef]
- Takada, N.; Umemura, N.; Suenaga, K.; Chou, T.; Nagatsu, A.; Haino, T.; Yamada, K.; Uemura, D. Pinnatoxins B and C, the Most Toxic Components in the Pinnatoxin Series from the Okinawan Bivalve Pinna Muricata. Tetrahedron Lett. 2001, 42, 3491–3494. [Google Scholar] [CrossRef]
- Matsuura, F.; Hao, J.; Reents, R.; Kishi, Y. Total Synthesis and Stereochemistry of Pinnatoxins B and C. Org. Lett. 2006, 8, 3327–3330. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Matsuura, F.; Kishi, Y.; Kita, M.; Uemura, D.; Asai, N.; Iwashita, T. Stereochemistry of Pteriatoxins A, B, and C. J. Am. Chem. Soc. 2006, 128, 7742–7743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selwood, A.I.; Miles, C.O.; Wilkins, A.L.; van Ginkel, R.; Munday, R.; Rise, F.; McNabb, P. Isolation, Structural Determination and Acute Toxicity of Pinnatoxins E, F and G. J. Agric. Food Chem. 2010, 58, 6532–6542. [Google Scholar] [CrossRef] [PubMed]
- Selwood, A.I.; Wilkins, A.L.; Munday, R.; Gu, H.; Smith, K.F.; Rhodes, L.L.; Rise, F. Pinnatoxin H: A New Pinnatoxin Analogue from a South China Sea Vulcanodinium Rugosum Isolate. Tetrahedron Lett. 2014, 55, 5508–5510. [Google Scholar] [CrossRef]
- Smith, K.F.; Rhodes, L.L.; Suda, S.; Selwood, A.I. A Dinoflagellate Producer of Pinnatoxin G, Isolated from Sub-Tropical Japanese Waters. Harmful Algae 2011, 10, 702–705. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; Munday, R.; Suda, S.; Molenaar, S.; Hallegraeff, G. Dinoflagellate Vulcanodinium Rugosum Identified as the Causative Organism of Pinnatoxins in Australia, New Zealand and Japan. Phycologia 2011, 50, 624–628. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; Molenaar, S.; Munday, R.; Wilkinson, C.; Hallegraeff, G. Production of Pinnatoxins E, F and G by Scrippsielloid Dinoflagellates Isolated from Franklin Harbour, South Australia. N.Z. J. Mar. Freshw. Res. 2011, 45, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; van Ginkel, R.; Holland, P.; Munday, R. Production of Pinnatoxins by a Peridinoid Dinoflagellate Isolated from Northland, New Zealand. Harmful Algae 2010, 9, 384–389. [Google Scholar] [CrossRef]
- McCarron, P.; Rourke, W.A.; Hardstaff, W.; Pooley, B.; Quilliam, M.A. Identification of Pinnatoxins and Discovery of Their Fatty Acid Ester Metabolites in Mussels (Mytilus Edulis) from Eastern Canada. J. Agric. Food Chem. 2012, 60, 1437–1446. [Google Scholar] [CrossRef]
- Selwood, A.I. Portimine: A Bioactive Metabolite from the Benthic Dinoflagellate Vulcanodinium Rugosum. Tetrahedron Lett. 2013, 54, 4705–4707. [Google Scholar] [CrossRef]
- Aráoz, R.; Barnes, P.; Séchet, V.; Delepierre, M.; Zinn-Justin, S.; Molgó, J.; Zakarian, A.; Hess, P.; Servent, D. Cyclic Imine Toxins Survey in Coastal European Shellfish Samples: Bioaccumulation and Mode of Action of 28-O-Palmitoyl Ester of Pinnatoxin-G. First Report of Portimine-A Bioaccumulation. Harmful Algae 2020, 98, 101887. [Google Scholar] [CrossRef]
- Fribley, A.M.; Xi, Y.; Makris, C.; Alves-de-Souza, C.; York, R.; Tomas, C.; Wright, J.L.C.; Strangman, W.K. Identification of Portimine B, a New Cell Permeable Spiroimine That Induces Apoptosis in Oral Squamous Cell Carcinoma. ACS Med. Chem. Lett. 2019, 10, 175–179. [Google Scholar] [CrossRef]
- Hermawan, I.; Higa, M.; Hutabarat, P.U.B.; Fujiwara, T.; Akiyama, K.; Kanamoto, A.; Haruyama, T.; Kobayashi, N.; Higashi, M.; Suda, S.; et al. Kabirimine, a New Cyclic Imine from an Okinawan Dinoflagellate. Mar. Drugs 2019, 17, 353. [Google Scholar] [CrossRef] [Green Version]
- Arnich, N.; Abadie, E.; Delcourt, N.; Fessard, V.; Fremy, J.-M.; Hort, V.; Lagrange, E.; Maignien, T.; Molgó, J.; Peyrat, M.-B.; et al. Health Risk Assessment Related to Pinnatoxins in French Shellfish. Toxicon 2020, 180, 1–10. [Google Scholar] [CrossRef]
- Munday, R.; Selwood, A.I.; Rhodes, L. Acute Toxicity of Pinnatoxins E, F and G to Mice. Toxicon 2012, 60, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Mondeguer, F.; Abadie, E.; Herve, F.; Bardouil, M.; Sechet, V.; Raimbault, V.; Berteaux, T.; Zendong, S.Z.; Palvadeau, H.; Amzil, Z.; et al. Pinnatoxines En Lien Avec L’espèce Vulcanodinium Rugosum (II); Ifremer: Brest, France, 2015. [Google Scholar]
- Servent, D.; Malgorn, C.; Bernes, M.; Gil, S.; Simasotchi, C.; Hérard, A.-S.; Delzescaux, T.; Thai, R.; Barbe, P.; Keck, M.; et al. First Evidence That Emerging Pinnatoxin-G, a Contaminant of Shellfish, Reaches the Brain and Crosses the Placental Barrier. Sci. Total Environ. 2021, 790, 148125. [Google Scholar] [CrossRef]
- Clarke, M.R.; Jones, B.; Squires, C.L.M.; Imhoff, F.M.; Harwood, D.T.; Rhodes, L.; Selwood, A.I.; McNabb, P.S.; Baird, S.K. Cyclic Imine Pinnatoxin G Is Cytotoxic to Cancer Cell Lines via Nicotinic Acetylcholine Receptor-Driven Classical Apoptosis. J. Nat. Prod. 2021, 84, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Johansson, N.; Alexander, J.; Benford, D.; Cockburn, A.; Dogliotti, E.; Di Domenico, A.; Fernández-Cruz, M.L.; Galli, C.; Grandjean, P.; Gzyl, J.; et al. EFSA Panel on Contaminants in the Food Chain (CONTAM) Opinion of the Scientific Panel on Contaminants in the Food Chain on a Request from the European Commission on Marine Biotoxins in Shellfish—Okadaic Acid and Analogues. EFSA J. 2008, 589, 1–62. [Google Scholar] [CrossRef]
- Braga, A.C.; Alves, R.N.; Maulvault, A.L.; Barbosa, V.; Marques, A.; Costa, P.R. In Vitro Bioaccessibility of the Marine Biotoxin Okadaic Acid in Shellfish. Food Chem. Toxicol. 2016, 89, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Doucet, E.; Ross, N.N.; Quilliam, M.A. Enzymatic Hydrolysis of Esterified Diarrhetic Shellfish Poisoning Toxins and Pectenotoxins. Anal. Bioanal. Chem. 2007, 389, 335–342. [Google Scholar] [CrossRef]
- García, C.; Truan, D.; Lagos, M.; Santelices, J.P.; Díaz, J.C.; Lagos, N. Metabolic Transformation of Dinophysistoxin-3 into Dinophysistoxin-1 Causes Human Intoxication by Consumption of O-Acyl-Derivatives Dinophysistoxins Contaminated Shellfish. J. Toxicol. Sci. 2005, 30, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Cuddihy, S.L.; Drake, S.; Harwood, D.T.; Selwood, A.I.; McNabb, P.S.; Hampton, M.B. The Marine Cytotoxin Portimine Is a Potent and Selective Inducer of Apoptosis. Apoptosis 2016, 21, 1447–1452. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on Marine Biotoxins in Shellfish—Cyclic Imines (Spirolides, Gymnodimines, Pinnatoxins and Pteriatoxins). EFSA J. 2010, 8, 1628. [Google Scholar] [CrossRef]
- Moreira-González, A.R.; Comas-González, A.; Valle-Pombrol, A.; Seisdedo-Losa, M.; Hernández-Leyva, O.; Fernandes, L.F.; Chomérat, N.; Bilien, G.; Hervé, F.; Rovillon, G.A.; et al. Summer Bloom of Vulcanodinium Rugosum in Cienfuegos Bay (Cuba) Associated to Dermatitis in Swimmers. Sci. Total Environ. 2020, 757, 143782. [Google Scholar] [CrossRef]
- Hess, P.; Mertens, K.; Chomerat, N.; Sechet, V.; Herve, F.; Plessis, L.; Reveillon, D.; Brehmer, P. Vulcanodinium Rugosum—A Potent and Ubiquitous Genus Affecting Mice and Man; Université Louis-et-Maximilien: München, Germany, 2022; Available online: https://hal.science/hal-03955437 (accessed on 27 July 2023).
- MacKenzie, L.A.; Selwood, A.I.; McNabb, P.; Rhodes, L. Benthic Dinoflagellate Toxins in Two Warm-Temperate Estuaries: Rangaunu and Parengarenga Harbours, Northland, New Zealand. Harmful Algae 2011, 10, 559–566. [Google Scholar] [CrossRef]
- McNabb, P.S. New Perspectives on Biotoxin Detection in Rangaunu Harbour, New Zealand Arising from the Discovery of Pinnatoxins. Harmful Algae 2012, 13, 34–39. [Google Scholar] [CrossRef]
- Rundberget, T.; Aasen, J.A.B.; Selwood, A.I.; Miles, C.O. Pinnatoxins and Spirolides in Norwegian Blue Mussels and Seawater. Toxicon 2011, 58, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; Herve, F.; Abadie, E.; Sechet, V.; Molgo, J.; Amzil, Z.; Fessard, V. Pinnatoxines En Lien Avec L’espèce Vulcanodinium Rugosum; Ifreme: Brest, France, 2012. [Google Scholar]
- Hess, P.; Abadie, E.; Hervé, F.; Berteaux, T.; Séchet, V.; Aráoz, R.; Molgó, J.; Zakarian, A.; Sibat, M.; Rundberget, T.; et al. Pinnatoxin G Is Responsible for Atypical Toxicity in Mussels (Mytilus galloprovincialis) and Clams (Venerupis decussata) from Ingril, a French Mediterranean Lagoon. Toxicon 2013, 75, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambla-Alegre, M. Occurrence of Cyclic Imines in European Commercial Seafood and Consumers Risk Assessment. Environ. Res. 2018, 7, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Lamas, J.P.; Arévalo, F.; Moroño, Á.; Correa, J.; Muñíz, S.; Blanco, J. Detection and Spatio-Temporal Distribution of Pinnatoxins in Shellfish from the Atlantic and Cantabrian Coasts of Spain. Toxins 2019, 11, 340. [Google Scholar] [CrossRef] [Green Version]
- Otero, P.; Miguéns, N.; Rodríguez, I.; Botana, L.M. LC–MS/MS Analysis of the Emerging Toxin Pinnatoxin-G and High Levels of Esterified OA Group Toxins in Galician Commercial Mussels. Toxins 2019, 11, 394. [Google Scholar] [CrossRef] [Green Version]
- Otero, P.; Vale, C.; Boente-Juncal, A.; Costas, C.; Louzao, M.C.; Botana, L.M. Detection of Cyclic Imine Toxins in Dietary Supplements of Green Lipped Mussels (Perna canaliculus) and in Shellfish Mytilus chilensis. Toxins 2020, 12, 613. [Google Scholar] [CrossRef]
- Norambuena, L.; Mardones, J.I. Emerging Phycotoxins in the Chilean Coast: First Localized Detection of the Neurotoxic Cyclic Imine Pinnatoxin-G in Shellfish Banks. Mar. Pollut. Bull. 2023, 190, 114878. [Google Scholar] [CrossRef]
- García-Altares, M.; Casanova, A.; Bane, V.; Diogène, J.; Furey, A.; de la Iglesia, P. Confirmation of Pinnatoxins and Spirolides in Shellfish and Passive Samplers from Catalonia (Spain) by Liquid Chromatography Coupled with Triple Quadrupole and High-Resolution Hybrid Tandem Mass Spectrometry. Mar. Drugs 2014, 12, 3706–3732. [Google Scholar] [CrossRef]
- Tamele, I.J.; Timba, I.; Vasconcelos, V.; Costa, P.R. First Report of Pinnatoxins in Bivalve Molluscs from Inhaca Island (South of Mozambique)—South of the Indian Ocean. J. Mar. Sci. Eng. 2022, 10, 1215. [Google Scholar] [CrossRef]
- Otero, P.; Silva, M. Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges. Mar. Drugs 2022, 20, 199. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M. Recent Trends in Inter-Laboratory Precision at Ppb and Sub-Ppb Concentrations in Relation to Fitness for Purpose Criteria in Proficiency Testing. Analyst 2000, 125, 385–386. [Google Scholar] [CrossRef]
- R Core Team R. A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing; Scientific Research: Wuhan, China, 2016; Available online: https://www.R-project.org/ (accessed on 27 July 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Sievert, C. Interactive Web-Based Data Visualization with R, Plotly, and Shiny; Chapman and Hall/CRC: London, UK, 2020; ISBN 978-1-138-33145-7. [Google Scholar]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef]
- Mattei, C.; Abadie, E.; Delcourt, N.; Fessard, V.; Fremy, J.-M.; Hess, P.; Hort, V.; Lagrange-Guillon, E.; Vernoux, J.P. Rapport d’expertise collective. In Risques Liés Aux Pinnatoxines Dans Les Coquillages; Avis de l’Anses: Paris, France, 2019; pp. 1–136. [Google Scholar]
- Catherine, B.; Nadine, N.-M.; Maud, L.; Anne, D.; Patrick, L.; Jean-Paul, B.; Hubert, G.; Eric, A.; Jean-Pierre, A.; Bruno, A.; et al. REPHY dataset—French observation and monitoring program for phytoplankton and hydrology in coastal waters REPHY Dataset—French Observation and Monitoring Program for Phytoplankton and Hydrology in Coastal Waters. 1987-2018 Metropolitan Data. SEANOE 2019. [Google Scholar] [CrossRef]
- Abadie, E.; Muguet, A.; Berteaux, T.; Chomérat, N.; Hess, P.; Roque D’OrbCastel, E.; Masseret, E.; Laabir, M. Toxin and Growth Responses of the Neurotoxic Dinoflagellate Vulcanodinium Rugosum to Varying Temperature and Salinity. Toxins 2016, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Dernekbasi, S.; Oksuz, A.; Celik, M.Y.; Karay, I.; Karay, S. The Fatty Acid Composition of Cultured Mussels (Mytilus galloprovincialis Lamarck 1819) in Offshore Longline System in the Black Sea. J. Aquac. Mar. Biol. 2015, 2, 00049. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Ji, Y.; Qiu, J.; Wang, G.; Tang, Z.; Li, A. Comparative Study on the Esterification of Gymnodimine in Different Shellfish Exposed to the Dissolved Toxin in Seawater. Harmful Algae 2022, 115, 102233. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Ji, Y.; Fang, Y.; Zhao, M.; Wang, S.; Ai, Q.; Li, A. Response of Fatty Acids and Lipid Metabolism Enzymes during Accumulation, Depuration and Esterification of Diarrhetic Shellfish Toxins in Mussels (Mytilus galloprovincialis). Ecotoxicol. Environ. Saf. 2020, 206, 111223. [Google Scholar] [CrossRef] [PubMed]
- Rossignoli, A.E.; Fernández, D.; Regueiro, J.; Mariño, C.; Blanco, J. Esterification of Okadaic Acid in the Mussel Mytilus galloprovincialis. Toxicon 2011, 57, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Lyu, C.; Chen, T.; Qiang, B.; Liu, N.; Wang, H.; Zhang, L.; Liu, Z. CMNPD: A Comprehensive Marine Natural Products Database towards Facilitating Drug Discovery from the Ocean. Nucleic Acids Res. 2021, 49, D509–D515. [Google Scholar] [CrossRef] [PubMed]
- European Union Reference Laboratory for Marine Biotoxins (EURL MB). EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS—Version 5; European Union Reference Laboratory for Marine Biotoxines: Vigo, Spain, 2015; pp. 1–33. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sampling Month | Levels (µg/kg) | |||||
|---|---|---|---|---|---|---|---|
| PnTX-A | PnTX-G | Portimine-A | |||||
| Free | Total | Free | Total | Free | Total | ||
| 18 BM 012 | January | 4.2 | 5.9 | 56.7 | 55.4 | 0.7 | 1.9 |
| 18 BM 026 | February | 4.6 | 6.3 | 68.8 | 68.2 | 0.6 | 1.0 |
| 18 BM 042 | March | 2.0 | 2.8 | 38.4 | 37.6 | <LOQ * | 0.7 |
| 18 BM 065 | April | 2.1 | 2.8 | 59.3 | 59.3 | 0.4 | 0.7 |
| 18 BM 107 | May | 1.1 | 2.1 | 48.1 | 52.0 | 2.4 | 3.0 |
| 18 BM 122 | June | 4.0 | 4.5 | 230.0 | 235.5 | 75.6 | 78.7 |
| 18 BM 150 | July | 6.7 | 6.2 | 467.5 | 475.2 | 32.4 | 59.4 |
| 18 BM 161 | August | 6.3 | 10.1 | 367.3 | 390.5 | 8.9 | 10.2 |
| 18 BM 194 | September | 3.1 | 3.9 | 319.4 | 314.4 | 0.9 | 1.7 |
| 18 BM 257 | October | 4.6 | 4.6 | 321.6 | 316.9 | 0.5 | 0.8 |
| 18 BM 271 | November | 1.2 | 2.1 | 27.4 | 25.5 | <LOQ * | 0.6 |
| 18 BM 286 | December | 2.3 | 3.1 | 12.7 | 14.3 | 0.5 | 0.6 |
| m/z | Elemental Composition | Δ ppm |
|---|---|---|
| 79.0548 | No proposal | / |
| 91.0547 | No proposal | / |
| 93.0704 | No proposal | / |
| 95.0495 | C6H7O | 3.8 |
| 108.0811 | C7H10N | 3.0 |
| 110.0604 | C6H8NO | 3.2 |
| 121.0648 | C8H9O | 0.4 |
| 134.0755 | C8H8NO | −1.2 |
| 148.0755 | C9H10NO | −1.3 |
| 164.1067 | C10H14NO | −1.6 |
| 202.1224 | C13H16NO | 0.4 |
| 218.1539 | C14H20NO | −0.4 |
| 228.1381 | C15H18NO | −0.9 |
| 246.1486 | C15H20NO2 | −1.0 |
| 336.1949 | C22H26NO2; [M + H − 3H2O − CO]+ | −2.6 |
| 346.1802 | C23H24NO2; [M + H − 4H2O]+ | 0.1 |
| 354.2063 | C22H28NO3; [M + H − 2H2O − CO]+ | −0.1 |
| 364.1907 | C23H26NO3; [M + H − 3H2O]+ | −0.1 |
| 372.2169 | C22H30NO4; [M + H − H2O − CO]+ | 0.0 |
| 382.2009 | C23H28NO4; [M + H − 2H2O]+ | −0.9 |
| 400.2115 | C23H30NO5; [M + H − H2O]+ | −0.8 |
| 418.2217 | C23H32NO6; [M + H]+ | −1.6 |
| Compound | Precursor Ion (m/z) | Product Ion * (m/z) | Collision Energy (V) |
|---|---|---|---|
| PnTX-A | 712.5 | 164.2 (Q) | 77 |
| 458.3 (q) | 61 | ||
| PnTX-B/C | 741.4 | 164.2 (Q) | 77 |
| 458.3 (q) | 61 | ||
| PnTX-D | 782.4 | 164.2 (Q) | 77 |
| 488.3 (q) | 61 | ||
| PnTX-E | 784.4 | 164.2 (Q) | 77 |
| 488.3 (q) | 61 | ||
| PnTX-F | 766.4 | 164.2 (Q) | 77 |
| 488.3 (q) | 61 | ||
| PnTX-G | 694.5 | 164.2 (Q) | 77 |
| 458.3 (q) | 61 | ||
| PnTX-H | 708.5 | 164.2 (Q) | 77 |
| 488.3 (q) | 61 | ||
| PtTXs | 831.5 | 164.2 (Q) | 77 |
| 458.3 (q) | 61 | ||
| Portimine-A | 402.2 | 384.2 (Q) | 22 |
| 246.2 (q) | 28 | ||
| Portimine-B | 418.2 | 400.2 (Q) | 22 |
| 246.2 (q) | 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hort, V.; Bastardo-Fernández, I.; Nicolas, M. Exploration of Vulcanodinium rugosum Toxins and their Metabolism Products in Mussels from the Ingril Lagoon Hotspot in France. Mar. Drugs 2023, 21, 429. https://doi.org/10.3390/md21080429
Hort V, Bastardo-Fernández I, Nicolas M. Exploration of Vulcanodinium rugosum Toxins and their Metabolism Products in Mussels from the Ingril Lagoon Hotspot in France. Marine Drugs. 2023; 21(8):429. https://doi.org/10.3390/md21080429
Chicago/Turabian StyleHort, Vincent, Isabel Bastardo-Fernández, and Marina Nicolas. 2023. "Exploration of Vulcanodinium rugosum Toxins and their Metabolism Products in Mussels from the Ingril Lagoon Hotspot in France" Marine Drugs 21, no. 8: 429. https://doi.org/10.3390/md21080429