
Applications of Antioxidant Secondary Metabolites of Sargassum spp.
 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Antioxidant Secondary Metabolites
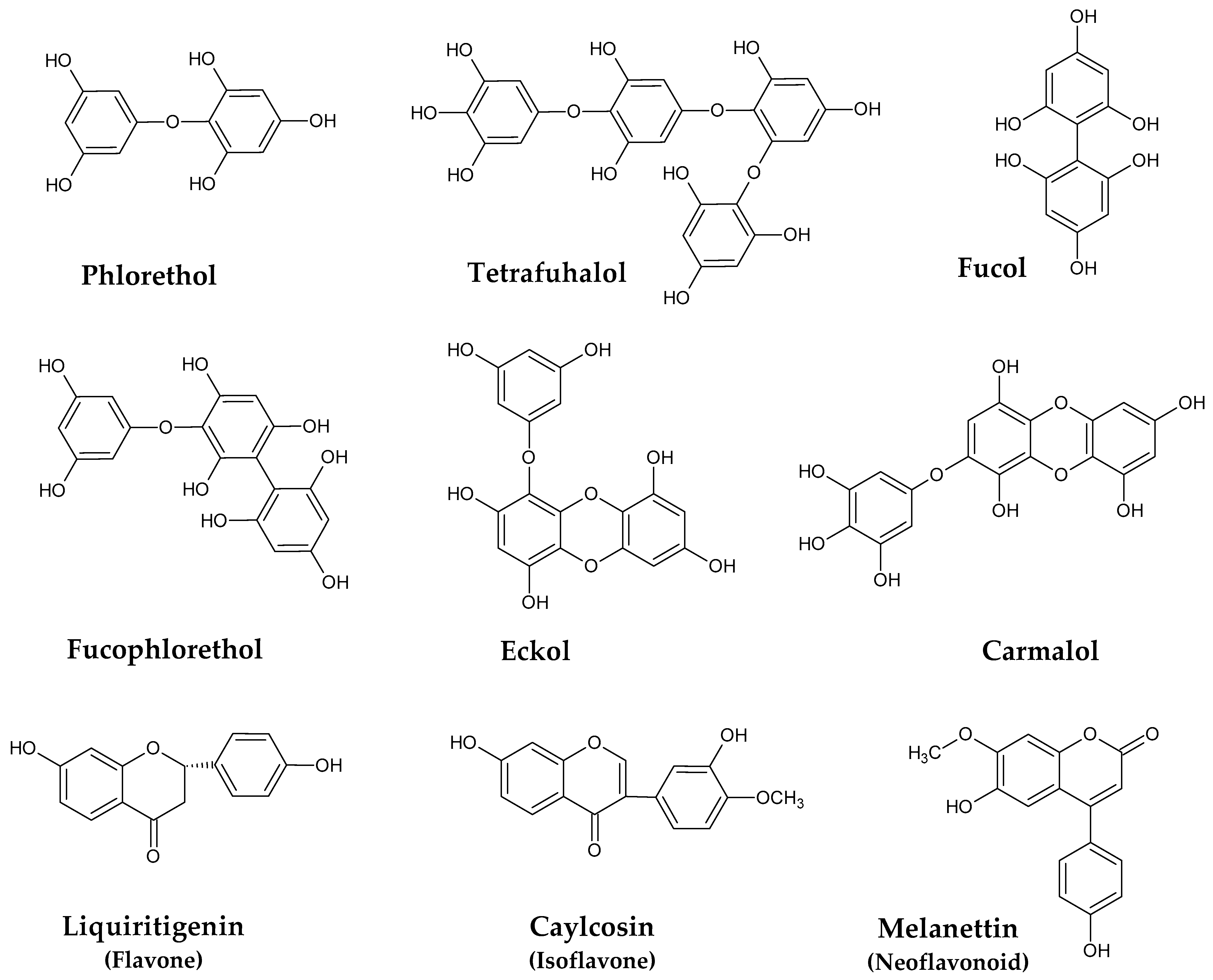
2.1. Phenolic Compounds
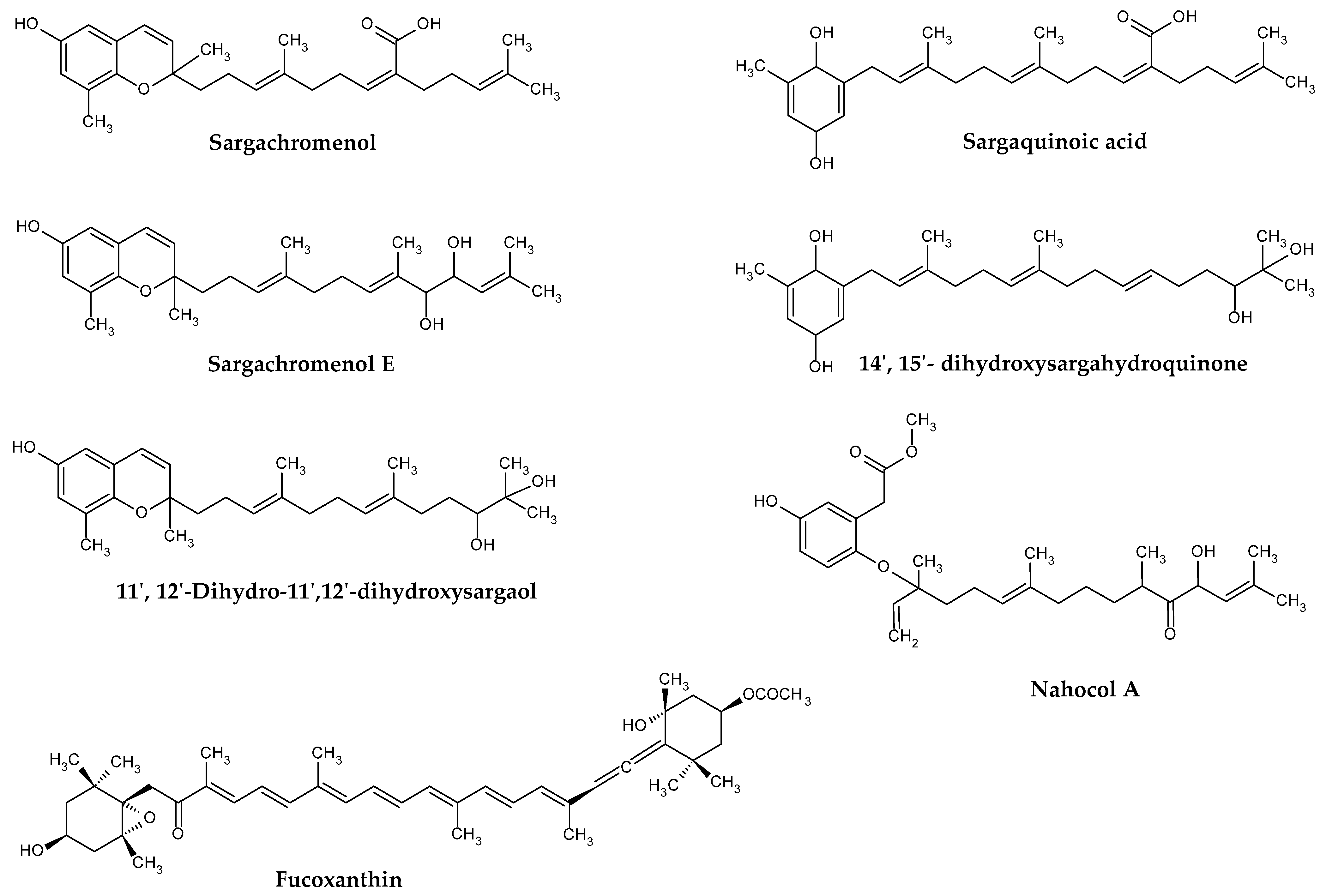
2.2. Terpenoids
2.3. Phytosterols
3. Bioactive Potential of Sargassum Antioxidant Secondary Metabolites
3.1. In Chemico Studies

{kind=link}
{kind=link}
| Sargassum spp. Extracts and Compounds | ||||
|---|---|---|---|---|
| Sargassum spp. | Extraction Conditions | Bioactive Compounds | In Chemico Antioxidant Properties | Ref. |
| S. acinarium | 80% MeOH; 80% EtOH; 80% Act; H2O | Alkaloids; phenolics, steroids, terpenoids | TAA MeOH; EtOH; Act; H2O: 2.1; 2.8; 1.7; 0.7 mg AA/g DW; RP MeOH; EtOH; Act; H2O: 0.4; 1.3; 0.4; 0.5 mg AA/g DW | [76] |
| S. angustifolium | EtOH; 50% EtOH; H2O | Cardiac glycosides, saponins, steroids, flavonoids | DPPH● EtOH; 50% EtOH; H2O: 1/EC50 = 14.3; 10.0; 0.7 mg/mL; FICA EtOH; 50% EtOH; H2O (% at 1 mg/mL): 15.6; 19.7; 88.3%; RP EtOH; 50% EtOH; H2O (OD 700 nm at 5 mg/mL): 1.0; 0.4; 0.1; TBARS EtOH; 50% EtOH; H2O (% at 1 mg/mL): 68.9; 23.3; 4.9% | [90] |
| S. aquifolium | EtOH; 50% EtOH; H2O | Cardiac glycosides, saponins, steroids, flavonoids | DPPH● EtOH; 50% EtOH; H2O: 1/EC50 = 2.9; 20.0; 0.7 mg/mL; FICA EtOH; 50% EtOH; H2O (% at 1 mg/mL): 7.3; 20.1; 52.0%; RP EtOH; 50% EtOH; H2O (OD 700 nm at 5 mg/mL): 0.4; 0.5; 0.7; TBARS EtOH; 50% EtOH; H2O (% at 1 mg/mL): 29.3; 10.3; 8.9% | [90] |
| EtOH | Phenolics | DPPH●: IC50 = 828.2 µg/mL | [77] | |
| S. asperifolium | EtOH; 50% EtOH; H2O | Cardiac glycosides, flavonoids, saponins, steroids, condensed tannins | DPPH● EtOH; 50% EtOH; H2O: 1/EC50 = 13.3; 40.0; 0.7 mg/mL; FICA EtOH; 50% EtOH; H2O (% at 1 mg/mL): 4.3; 10.9; 42.9%; RP EtOH; 50% EtOH; H2O (OD 700 nm at 5 mg/mL): 1.0; 0.7; 0.2; TBARS EtOH; 50% EtOH; H2O (% at 1 mg/mL): 53.7; 11.3; 3.3% | [90] |
| S. boveanum | EtOH; 50% EtOH; H2O | Cardiac glycosides, alkaloids, saponins, steroids, flavonoids | DPPH● EtOH; 50% EtOH; H2O: 1/EC50 = 100.0; 20.0; 1.2 mg/mL; FICA EtOH; 50% EtOH; H2O (% at 1 mg/mL): 3.9; 23.4; 78.8%; RP EtOH; 50% EtOH; H2O (OD 700 nm at 5 mg/mL): 1.5; 1.1; 0.6; TBARS EtOH; 50% EtOH; H2O (% at 1 mg/mL): 58.9; 20.1; 1.3% | [90] |
| Ext: MeOH; Fract: DCM → EtOAc → BuOH → H2O | Phenolics | DPPH● MeOH; DCM; EtOAc; BuOH; H2O: EC50 = 1091.7; 245.9; 171.4; 779.9; 1987.1 ppm; ABTS●+ MeOH; DCM; EtOAc; BuOH; H2O: EC50 = 1204.6; 247.6; 219.5;487.9; 1556.4 ppm; FRAP MeOH; DCM; EtOAc; BuOH; H2O: EC50 = 535.8; 157.9; 129.2; 243.9; 1381.7 ppm | [91] | |
| S. cinctum | H2O; MeOH | Phenolics | TAA H2O; MeOH: 38.3; 21.9 mg AA/g DW; DPPH● H2O; MeOH: IC50 = 1.1; 1.2 mg/mL; FRAP H2O; MeOH: 12.3; 3.7 mg AA/g DW | [91] |
| S. coriifolium | MeOH; EtOH; H2O | Terpenoids; saponins; phlorotannins; cardiac glycosides; flavonoids; phenols | DPPH● MeOH; EtOH; H2O: IC50 = 1.0; 1.4; 2.7 mg/mL; ABTS●+ MeOH; EtOH; H2O: IC50 = 1.6; 2.2; 3.7 mg/mL; H2O2 MeOH; EtOH; H2O: IC50 = 2.0; 2.6; 4.7 mg/mL | [92] |
| S. crassifolium | EtOH | Phenolics | DPPH●: IC50 = 767.0 µg/mL | [75] |
| S. cristaefolium | EtOH | Phenolics | DPPH●: IC50 = 737.3 µg/mL | [77] |
| Ext: CHCl3:MeOH:H2O (1:2:0.8) Fract: DCM → Act → MeOH | Fucoxanthin, porphyrin derivatives, galactosyldiacylglycerols | TAA Ext; DCM; Act; MeOH: 39.2; 46.2; 66.1; 7.8 ìmol TE/g DPPH● Ext; DCM; Act; MeOH (at 0.5 mg/mL): 41.3; 48.6; 67.2; 11.9% FRAP Ext; DCM; Act; MeOH: 688.1; 368.1; 679.2; 132.1 ìmol FE/g | [79] | |
| S. duplicatum | EtOH; MeOH; EtOAc | Fucoxanthin | DPPH● EtOH; MeOH; EtOAc: IC50 = 93.8; 78.5; 112.3 ìg/mL | [83] |
| S. furcatum | DCM:MeOH (2:1) | Phenolics | DPPH●: EC50 = 0.5 mg/L; ABTS●+: EC50 = 0.3 mg/L | [93] |
| S. horneri | 70% MeOH | Phenolics | DPPH●: EC50 = 06 mg/mL; H2O2: EC50 = 83.9 mg/mL; O2●-: EC50 = 0.5 mg/mL; OH●: EC50 = 1.4 mg/mL; RP: EC50 = 0.2 mg/mL; FICA: 0.4 mg/mL | [94] |
| MeOH | Fucoxanthin, phenolics | DPPH● (at 2 mg/mL): 46.5%; RP (OD 700 nm at 2 mg/mL): 0.8 | [95] | |
| Ext: 80% MeOH Fract: CHCl3 | Phenolics, polysaccharides, sterols, Apo-9 fucoxanthinone | DPPH●Ext; CHCl3: IC50 = 1.1; 2.7 mg/mL; Alkyl | [96] | |
| S. linearifolium | 80% MeOH → hot H2O | Phenolics, alkaloids, terpenoids, sterols | DPPH● hot H2O: IC50 = 124.5 µg/mL; ABTS●+ hot H2O: IC50 = 257.1 µg/mL | [97] |
| 70% EtOH | Fucoxanthin, Phenolics | ABTS●+ (at 1 mg/mL): 34.5 mg TE/g extract; DPPH● (at 1 mg/mL): 15.6 mg TE/g DW; FRAP (at 1 mg/mL): 12.5 mg TE/g DW | [84] | |
| S. miyabei | 70% EtOH | Phenolics | DPPH● (% at 0.1 mg/mL): 42.8%; OH● (% at 0.1 mg/mL): 43.8%; LP (% at 0.1 mg/mL): 52.4% | [98] |
| 70% EtOH | SHQA, SCM, Phenolics | ABTS●+: 186.2 mg VCE/g; DPPH●: 193.7 mg VCE/g; FRAP: 1.0 mM FE/g | [52] | |
| S. muticum | 80% MeOH; 80% EtOH; 80% Act; H2O | Alkaloids; phenolics, steroids, terpenoids | TAA MeOH; EtOH; Act; H2O: 0.9; 3.1; 2.3; 0.7 mg AA/g DW; RP MeOH; EtOH; Act; H2O: 0.2; 1.4; 1.0; 0.7 mg AA/g DW | [76] |
| EtOH; Act; EtOAc; CHCl3; Hex | Phenolics | DPPH● EtOH; Act; EtOAc; CHCl3; Hex: 4.0; 5.4; 16.2; 6.1;4.7 mg TE/g DW; FRAP EtOH; Act; EtOAc; CHCl3; Hex: 4.2; 17.5; 18.3; 6.0; 6.2 mg AA/g DW | [99] | |
| CHCl3:MeOH (1:1); SC CO2 | Glycolipids | DPPH● CHCl3:MeOH; SC CO2: EC50 = 4.1; 0.9 mg/mL | [100] | |
| CHCl3:MeOH (1:1); SC CO2 | Phospholipids | DPPH● CHCl3:MeOH; SC CO2: EC50 = 4.8; 1.0 mg/mL | [100] | |
| 50% MeOH | Phenolics | DPPH●: IC50 = 1.64 mg/mL | [101] | |
| EAE-PLE 4h with alcalase or vicozyme | Phenolics; phlorotannins | TEAC alcalase; viscozyme: 0.5; 0.6 mmol TE/g | [102] | |
| S. naozhouense | 75% EtOH → Column chromatography | (+)-epiloliolide | DPPH●: IC50 = 17 mM | [54] |
| S. oligocystum | EtOH; 50% EtOH; H2O | Cardiac glycosides, alkaloids, saponins, steroids, flavonoids, condensed tannins | DPPH● EtOH; 50% EtOH; H2O: 1/EC50 = 12.5; 12.5; 5.0 mg/mL; FICA EtOH; 50% EtOH; H2O (% at 1 mg/mL): 7.3; 20.1; 52.0% RP EtOH; 50% EtOH; H2O (OD 700 nm at 5 mg/mL): 1.1; 0.6; 0.5 TBARS EtOH; 50% EtOH; H2O (% at 1bmg/mL): 59.3; 22.0; 2.7% | [54] |
| S. podocanthum | 70% EtOH | Fucoxanthin, Phenolics | ABTS●+ (at 1 mg/mL): 147.1 mg TE/g extract; DPPH● (at 1 mg/mL): 136.6 mg TE/g DW; FRAP (at 1 mg/mL): 29.6 mg TE/g DW | [84] |
| S. polycystum | CHCl3; Hex; MeOH; Act; 70% EtOH | Steroids, phenolics, saponins, terpenoids | TAA CHCl3; Hex; MeOH; Act; 70% EtOH (at 1.25 mg/mL): 68.0; 61.4; 121.0; 46.0; 120.0 mmol AA/g DW | [103] |
| EtOH | Phenolics | DPPH●: IC50 = 804.3 µg/mL | [77] | |
| H2O; MeOH | Phenolics | TAA H2O; MeOH: 43.3; 36.1 mg AA/g DW; DPPH● H2O; MeOH: IC50 = 1.0; 1.2 mg/mL; FRAP H2O; MeOH: 11.6; 3.0 mg AA/g DW | [104] | |
| EtOH | Carotenoids | DPPH● (at 1 mg/mL): 20.4%; ORAC: 42.1 mmol TE/100 g | [81] | |
| S. serrifolium | 70% EtOH | SHQA, SCM, Phenolics | ABTS●+: 99.5 mg VCE/g; DPPH●: 75.4 mg VCE/g; FRAP: 0.3 mM FE/g | [52] |
| 70% EtOH → HPLC purification | SHQA, SCM, SQA | ABTS●+ SHQA, SCM, SQA: IC50 = 13.8; 13.1; 47.3 µg/mL; DPPH● SHQA, SCM, SQA: IC50 = 5.9; 8.0; 15.3 µg/mL; O2●- SHQA, SCM, SQA: IC50 = 16.9; 14.5; 20.5 µg/mL; ROS: IC50 = 0.2; 0.2; 0.3 µg/mL | [56] | |
| S. tenerrium | H2O; MeOH | Phenolics | TAA H2O; MeOH: 52.0; 40.1 mg AA/g DW; DPPH● H2O; MeOH: IC50 = 0.8; 0.8 mg/mL; FRAP H2O; MeOH: 13.4; 4.0 mg AA/g DW | [104] |
| S. thunbergii | 70% EtOH | Phenolics | DPPH● (% at 0.1 mg/mL): 39.3 %; OH● (% at 0.1 mg/mL): 34.3 %; LP (% at 0.1 mg/mL): 31.0% | [98] |
| MeOH | Fucoxanthin, phenolics | DPPH● (at 2 mg/mL): 90.4%; RP (OD 700 nm at 2 mg/mL): 0.5 | [95] | |
| S. vestitum | 70% EtOH | Phenolics | ABTS●+: 40.3 mg TE/g DW; DPPH●: 111.8 mg TE/g DW; FRAP: 46.2 mg TE/g DW | [105] |
| MAE 70% EtOH | Phenolics | ABTS●+: 149.8 mg TE/g DW; DPPH●: 116.5 mg TE/g DW; FRAP: 68.0 mg TE/g DW | [105] | |
| UAE 70% EtOH | Phenolics | ABTS●+: 147.2 mg TE/g DW; DPPH●: 86.1 mg TE/g DW; FRAP: 60.1 mg TE/g DW | [105] | |
| 70% EtOH | Fucoxanthin, Phenolics | ABTS●+ (at 1 mg/mL): 183.4 mg TE/g extract; DPPH● (at 1 mg/mL): 209.5 mg TE/g DW; FRAP (at 1 mg/mL): 283.7 mg TE/g DW | [84] | |
| S. vulgare | Act | Phenolics | DPPH●: IC50 = 0.8 mg/mL; O2●-: IC50 = 1.0 mg/mL; RP (OD 700 nm at 1 mg/mL): 0.3 | [106] |
| MeOH; H2O; hot H2O | Phenolics | ABTS●+ MeOH; H2O; hot H2O: EC50 = 165.8; 73.3; 46.5 µg/mL; β-CBT MeOH; H2O; hot H2O: EC50 = 18.2; 142.7; 66.5 µg/mL; FRAP: 216.4; 102.1; 198.3µg TE/mL | [107] | |
| Ext: 70% Act Fract: Hex → EtOAc → H2O | Phlorotannins | ABTS●+Ext; Hex; EtOAc; H2O: IC50 = 72.9; 93.8; 25.1; 74.9 µg/mL; DPPH● Ext; Hex; EtOAc; H2O: IC50 = 97.4; 29.8; 25.8; 96.6 µg/mL O2●- Hex; EtOAc: IC50 = 37.1; 27.0 µg/mL; β-CBT Ext; Hex; EtOAc: IC50 = 65.2; 41.3; 72.1 µg/mL; | [108] | |
| S. wightii | SFE CO2:EtOH (94:6); EtOH; 60% EtOH; 40% EtOH; H2O | Phenolics, protein | TAA SFE; EtOH; 60% EtOH; 40% EtOH; H2O: 67.9; 21.6; 23.0; 31.1; 33.3 mg AA/g extract; FRAP SFE; EtOH; 60% EtOH; 40% EtOH; H2O: 57.3; 45.4; 55.7; 62.3; 68.7 mg AA/g extract | [109] |
| Hex → EtOAc → MeOH | Phlorotannins | DPPH● EtOAc: IC50 = 59.9 µg/mL; ABTS●+ EtOAc: IC50 = 51.0 µg/mL; FRAP EtOAc: 55.2 µg/mL | [110] | |
| EtOAc → Column chromatography → TLC | Fucoxanthin | DPPH●: IC50 = 79.6 µM; ABTS●+: IC50 = 76.0 µM | [85] | |
3.2. Cellular and In Vivo Studies
3.2.1. Oxidative Stress Protective Effects
3.2.2. Effects on Oxidative-Stress Related Disorders
| Sargassum spp. | Solvents | Secondary Antioxidants | Cells | Oxidative Stress Inductors | Effects | Ref. |
|---|---|---|---|---|---|---|
| S. angustifolium | 80% EtOH | PPS | SH-SY5Y | Methamphetamine | ↓ ROS | [127] |
| S. fusiformis | Ext: 80% MeOH Frac: Chl and water | fucoxanthin (↑%) | HaCaT | Particulate matter exposure | ↓ ROS | [125] |
| S. glaucescens | Ext: 70% Ace in EtOH Frac: 90% EtOH, EtOH:water:Hex (9:1:10) | fucoxanthin (↑%) | RAW 264.7 | LPS-stimulation | ↓ ROS/O2- | [122] |
| S. horneri | 80% MeOH | PPS (↑%) | HaCaT | UV-B | ↓ ROS | [118] |
| 95% EtOH | PPS (↑%) | MLE-12 | Particulate matter | ↓ ROS/LPO; ↑ SOD/CAT | [123] | |
| 70% EtOH | PPS (↑%), | HaCaT | Fine dust | ↓ ROS | [124] | |
| Ext: 70% EtOH Frac: Hex, DCM, EtOAc, BuOH | Fucosterol | BV2 HT22 | LPS (BV2 cells); Glu (HT22 cells) | BV2 cells: ↑ HO-1/Nrf2 (DCM fraction) HT22 cells: ↓ ROS (DCM fraction) | [141] | |
| S. muticum | Ext: MeOH:DCM Frac: MeOH:DCM, MeOH | PPS | MCF-7 | H2O2 | ↓ H2O2 | [111] |
| Ext: 80% EtOH Frac: Hex, DCM, EtOAc, BuOH, H2O | PPS | HaCaT | UV-B | ↓ ROS/LPO; ↑ SOD/CAT | [117] | |
| S. plagiophyllum | Distilled water | PPS | Human normal colon | H2O2 | ↓ ROS | [142] |
| S. polycystum and S. natans | Enzymes: Viscozyme, Celluclast, AMG, Termamyl, and Ultraflo | PPS | Chang | Extraction with different enzymes | ↑ ROS scavenging effects in Celluclast Ext | [112] |
| S. sagamanum | 80% EtOH | PPS | INS-1 | Glucose | ↓ ROS/LPO | [133] |
| S. serratiflium | 70% EtOH | SHQA, SC, SQA (↑%) | HepG2 | t-BHP | ↓ ROS/ LPO; Prevention of GSH oxidation; ↓ SOD/CAT; ↑ GST; ↑ Nrf2 Independent of t-BHP: ↑ HO-1/Nrf2 | [57] |
| Ext: EtOAc, MeOH, EtOH, Ace, Hex, Chl, H2O | SHQA, SC, SQA (↑%) | RAW 264.7 | LPS | ↑ TPC (EtOAc, MeOH and EtOH Ext) ↓ ROS (EtOAc, MeOH, and EtOH Ext) | [56] | |
| S. thunbergii | 40% EthOH | PPS (↑%) | L929 | UV-B | ↓ ROS/LPO; ↑ SOD and CAT | [119] |
| S. binderi | Ext: 70% EtOH Frac: Hex, Chl, EtOAc | Fucosterol | A549 | Fine dust | ↓ ROS; ↑ HO-1, SOD, CAT, and Nrf2 | [126] |
| S. carpophyllum | Ext: 80% MeOH Frac: Chl and H2O | 3 phlorotannins | RBL-2H3 | DNP-HAS | ↓ ROS | [138] |
| S. horneri | Ext: 80% MeOH Frac: Chl | (−)-Loliolide | Vero | AAPH | ↓ ROS | [44] |
| Ext: 80% MeOH Frac: Hex, EtOAc, MeOH, H2O | (−)-loliolide | HaCaT | UV-B | ↓ ROS | [45] | |
| Ext: MeOH Frac: Hex, 85% Aq MeOH, BuOH, H2O | SC E | HDF | UV-A | ↓ ROS, LPO and membrane protein oxidation | [47] | |
| Ext: 80% MeOH Frac: Hex, Chl, EtOAc | (−)-loliolide | HaCaT | Fine dust | ↓ ROS | [46] | |
| EtOH | Fucoxanthin | PC12 | Methamphetamine | ↓ ROS; ↑ SOD and CAT; ↑ HO-1 and Nrf2 | [128] | |
| Ext: 70% EtOH Frac: Hex, EtOAc | SCM | RAW 264.7 | LPS | ↓ ROS; ↑ HO-1/Nrf2 | [48] | |
| Ext: 70% EtOH Frac: Hex | Fucosterol | HDF | TNF-á/IFN-ã | ↓ ROS; ↑ HO-1/Nrf2 | [143] | |
| S. macrocarpum | 80% EtOH | SHQA | RAW 264.7 | LPS | ↓ ROS; ↑ HO-1 | [49] |
| S. muticum | Ext: MeOH Frac: Hex, DCM, EtOAc, BuOH | TPM | HDF | UV-A | ↓ ROS | [53] |
| S. serratifolium | Ext: 95% EtOH, H2O/ EtOH Frac: Hex, EtOAc, BuOH, H2O | SHQA | KU812F | PMACI | ↓ ROS | [144] |
| S. siliquastrum | Ext: 80% MeOH Frac: Chl | Fucoxanthin | Vero | H2O2 | ↓ ROS | [116] |
| Ext: MeOH Frac: Hex, 85% Aq MeOH, BuOH, H2O | SCM D, E, and K, 3 chromanols | HT1080 | H2O2 | ↓ ROS/LPO; ↑ GSH | [59] | |
| Ext: 80% MeOH Frac: Chl | Fucoxanthin | Human fibroblasts | UV-B | ↓ ROS | [120] | |
| S. thunbergii | Ext: 80% MeOH; Frac: Chl | I6CA | V79-4 | H2O2 | ↓ ROS; ↑ HO-1 and Nrf2 | [62,63] |
| Ext: MeOH Frac: Hex, 85% aq MeOH, BuOH, H2O | SC E, SC D, SHQA | HT1080 | Fe(II)/H2O2; AAPH | ↓ ROS; ↓ LPO | [61] |
| Sargassum spp. | Solvents | Secondary Antioxidants | Animal Model | Oxidative Stress Inductors | Effects | Ref. |
|---|---|---|---|---|---|---|
| ND | H2O | PPS (↑%) | Wistar rats | CCl4 | ↑ GPx | [114] |
| S. angustifolium | 70% EtOH | PPS (↑%) | Wistar rats | Dexamethasone | ↓ LPO | [140] |
| 70% EtOH | PPS | Wistar rats | CdCl2 | ↑ TAOC | [139] | |
| S. fusiformis | Ext: 80% MeOH Frac: Chl, H2O | fucoxanthin (↑%) | Zebrafish embryos | Particulate matter | ↓ ROS | [125] |
| 95% EtOH | Flv, fucoxanthin, fucosterol | ICR mice | HFD/STZ | ↓ LPO; ↑ SOD and CAT | [134] | |
| S. glaucescens | Ext: 70% Ace in EtOH Frac: 90% EtOH, EtOH: H2O:Hex | Fucoxanthin (↑%) | Syrian hamsters | Cisplatin chemotherapy | ↑ SOD (plasma); ↑ GPx and CAT (testicles) | [122] |
| S. hemiphyllum | Ext: 70% EtOH Frac: EtOAc | Phlorotannins | Kunming mice | CCl4 | ↑ SOD (serum, kidney); ↑ TAOC (serum, kidney, and liver); ↑ CAT (kidney, brain, and liver); ↑ GPx (brain and liver); ↓ LPO (kidney) | [115] |
| S. longifollium | EtOH | Terpenoids, PPS, and Flv (encapsulated in Na-Caseinate matrix) | Fingerling Tilapia | A. salmonicida | SOD and LPO levels returned to normal | [145] |
| S. pallidum | 70% aq Ace | PPS (↑%) | Wistar rats | CCl4 | ↓ LPO; ↑ SOD and GSH | [113] |
| S. policystum | Enzymes: Celluclast | PPS | Zebrafish embryos | H2O2 | ↓ ROS | [112] |
| H2O | PPS (↑%) | Sprague–Dawley rats | HCF diet | ↓ LPO; ↑ SOD | [146] | |
| S. thunbergii | Ext: 40% EtOH Frac: EtOAc | PPS (↑%) | Zebrafish embryos | UV-B | ↓ ROS | [119] |
| S. virgatum | EtOH | PPS | Wistar rats | ã-irradiation | ↑ SOD, CAT, GPx, and GSH | [121] |
| S. vulgare | MeOH | ↑ Total phenolics and Flv | Wistar Rats | ASP | ↓ LPO; ↑ SOD and CAT | [147] |
| S. horneri | Ext: 80% MeOH Frac: Chl | (−)-Loliolide | Zebrafish | AAPH | ↓ LPO and ROS | [44] |
| Ext: 80% MeOH Frac: Hex, EtOAc, MeOH, H2O | (−)-Loliolide | Zebrafish | UV-B | ↓ LPO an ROS | [45] |
4. Applications in Non-Patented and in Patented Products
4.1. Food Applications
4.2. Cosmetic Applications
4.3. Applications as Biostimulants
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mattio, L.; Payri, C.E. 190 Years of Sargassum Taxonomy, Facing the Advent of DNA Phylogenies. Bot Rev. 2011, 77, 31–70. [Google Scholar] [CrossRef]
- Amador-Castro, F.; Garcia-Cayuela, T.; Alper, H.S.; Rodriguez-Martinez, V.; Carrillo-Nieves, D. Valorization of pelagic sargassum biomass into sustainable applications: Current trends and challenges. J. Environ. Manag. 2021, 283, 112013. [Google Scholar] [CrossRef]
- Sargasso Sea Commission About the Sargasso Sea. Available online: http://www.sargassoseacommission.org/sargasso-sea/about-the-sargasso-sea (accessed on 5 January 2023).
- Rodriguez-Martinez, R.E.; Jordan-Dahlgren, E.; Hu, C.M. Spatio-temporal variability of pelagic Sargassum landings on the northern Mexican Caribbean. Remote Sens. Appl. Soc. Environ. 2022, 27, 100767. [Google Scholar] [CrossRef]
- Monteiro, C.A.; Engelen, A.H.; Santos, R. Habitat-related differences in recruitment and survival of early recruits of the invasive Sargassum muticum (Phaeophyta, Sargassaceae) in northern Portugal. Hydrobiologia 2012, 683, 287–296. [Google Scholar] [CrossRef]
- Raoux, A.; Pezy, J.-P.; Sporniak, T.; Dauvin, J.-C. Does the invasive macro-algae Sargassum muticum (Yendo) Fensholt, 1955 offer an appropriate temporary habitat for mobile fauna including non indigenous species? Ecological. Indic. 2021, 126, 107624. [Google Scholar] [CrossRef]
- Fernandez, C. Boom-bust of Sargassum muticum in northern Spain: 30 years of invasion. Eur. J. Phycol. 2020, 55, 285–295. [Google Scholar] [CrossRef]
- Engelen, A.H.; Primo, A.L.; Cruz, T.; Santos, R. Faunal differences between the invasive brown macroalga Sargassum muticum and competing native macroalgae. Biol. Invasions 2013, 15, 171–183. [Google Scholar] [CrossRef]
- Rushdi, M.I.; Abdel-Rahman, I.A.M.; Saber, H.; Attia, E.Z.; Abdelraheem, W.M.; Madkour, H.A.; Hassan, H.M.; Elmaidomy, A.H.; Abdelmohsen, U.R. Pharmacological and natural products diversity of the brown algae genus Sargassum. RSC Adv. 2020, 10, 24951–24972. [Google Scholar] [CrossRef]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef]
- Baweja, P.; Kumar, S.; Sahoo, D.; Levine, I. Chapter 3—Biology of Seaweeds. In Seaweed in Health and Disease Prevention; Fleurence, J., Levine, I., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 41–106. [Google Scholar]
- Desrochers, A.; Cox, S.-A.; Oxenford, H.A.; Tussenbroek, B. Sargassum Uses Guide: A Resource for Caribbean Researchers, Entrepreneurs and Policy Makers; FAO: Rome, Italy; University of the West Indies, Cave Hill Campus: Bridgetown, Barbados, 2020; p. 172. [Google Scholar]
- Aaron-Amper, J.; Largo, D.B.; Handugan, E.R.B.; Nini, J.L.; Alingasa, K.M.A.; Gulayan, S.J. Culture of the tropical brown seaweed Sargassum aquifolium: From hatchery to field out-planting. Aquacult. Rep. 2020, 16, 102591. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Jiang, X.D.; Li, H.Y.; Liu, T.H.; Ji, L.; Sun, Y.Q. Utilization of different seaweeds (Sargassum polycystum, Sargassum thunbergii, Sargassum horneri, Enteromorpha prolifera, Macrocystis pyrifera, and the residue of M. pyrifera) in the diets of sea cucumber Apostichopus japonicus (Selenka, 1867). Algal. Res. 2022, 61, 102591. [Google Scholar] [CrossRef]
- Singh, B.K.; Chopra, R.C.; Rai, S.N.; Verma, M.P.; Mohanta, R.K. Nutritional Evaluation of Seaweed on Nutrient Digestibility, Nitrogen Balance, Milk Production and Composition in Sahiwal Cows. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 437–443. [Google Scholar] [CrossRef]
- Casas-Valdez, M.; Portillo-Clark, G.; Aguila-Ramírez, N.; Rodríguez-Astudillo, S.; Sánchez-Rodríguez, I.; Carrillo-Domínguez, S. Efecto del alga marina Sargassum spp. sobre las variables productivas y la concentración de colesterol en el camarón café, Farfantepenaeus californiensis (Holmes, 1900). Rev. Biol. Mar. Oceanogr. 2006, 41, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Resources, Government of Bermuda—Department of Environmental and Natural Resources Sargassum spp. Seaweeds. Available online: https://environment.bm/sargassum-seaweed (accessed on 5 January 2023).
- Rosa, G.P.; Tavares, W.R.; Sousa, P.M.C.; Pages, A.K.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed Secondary Metabolites with Beneficial Health Effects: An Overview of Successes in In Vivo Studies and Clinical Trials. Mar. Drugs 2020, 18, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Fucaceae: A Source of Bioactive Phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catarino, M.D.; Pires, S.M.G.; Silva, S.; Costa, F.; Braga, S.S.; Pinto, D.C.G.A.; Silva, A.M.S.; Cardoso, S.M. Overview of Phlorotannins' Constituents in Fucales. Mar. Drugs 2022, 20, 754. [Google Scholar] [CrossRef]
- Bedoux, G.; Hardouin, K.; Burlot, A.S.; Bourgougnon, N. Chapter Twelve—Bioactive Components from Seaweeds: Cosmetic Applications and Future Development. In Advances in Botanical Research; Bourgougnon, N., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 71, pp. 345–378. [Google Scholar]
- Li, Y.X.; Wijesekara, I.; Li, Y.; Kim, S.K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Imbs, T.I.; Zvyagintseva, T.N. Phlorotannins are Polyphenolic Metabolites of Brown Algae. Russ. J. Mar. Biol. 2018, 44, 263–273. [Google Scholar] [CrossRef]
- Catarino, M.D.; Amarante, S.J.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Brown Algae Phlorotannins: A Marine Alternative to Break the Oxidative Stress, Inflammation and Cancer Network. Foods 2021, 10, 1478. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.; Cruz, M.T.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Phlorotannins from Fucus vesiculosus: Modulation of Inflammatory Response by Blocking NF-κB Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 6897. [Google Scholar] [CrossRef]
- Catarino, M.D.; Fernandes, I.; Oliveira, H.; Carrascal, M.; Ferreira, R.; Silva, A.M.S.; Cruz, M.T.; Mateus, N.; Cardoso, S.M. Antitumor Activity of Fucus vesiculosus-Derived Phlorotannins through Activation of Apoptotic Signals in Gastric and Colorectal Tumor Cell Lines. Int. J. Mol. Sci. 2021, 22, 7604. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Circuncisao, A.R.; Neves, B.; Marcal, C.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Impact of Gastrointestinal Digestion on the Anti-Inflammatory Properties of Phlorotannins from Himanthalia elongata. Antioxidants 2022, 11, 1518. [Google Scholar] [CrossRef]
- Catarino, M.D.; Marcal, C.; Bonifacio-Lopes, T.; Campos, D.; Mateus, N.; Silva, A.M.S.; Pintado, M.M.; Cardoso, S.M. Impact of Phlorotannin Extracts from Fucus vesiculosus on Human Gut Microbiota. Mar. Drugs 2021, 19, 375. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of Phlorotannins Extraction from Fucus vesiculosus and Evaluation of Their Potential to Prevent Metabolic Disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trench, C.; Thomas, S.L.; Thorney, D.; Maddix, G.M.; Francis, P.; Small, H.; Machado, C.B.; Webber, D.; Tonon, T.; Webber, M. Application of Stranded Pelagic Sargassum Biomass as Compost for Seedling Production in the Context of Mangrove Restoration. Front Environ. Sci. 2022, 10, 932293. [Google Scholar] [CrossRef]
- Martinez, J.H.I.; Castaneda, H.G.T. Preparation and Chromatographic Analysis of Phlorotannins. J. Chromatogr. Sci. 2013, 51, 825–838. [Google Scholar] [CrossRef] [Green Version]
- Montero, L.; Sanchez-Camargo, A.P.; Garcia-Canas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibanez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef]
- Liu, X.; Wang, C.Y.; Shao, C.L.; Wei, Y.X.; Wang, B.G.; Sun, L.L.; Zheng, C.J.; Guan, H.S. Chemical constituents from Sargassum pallidum (Turn.) C. Agardh. Biochem. Syst. Ecol. 2009, 37, 127–129. [Google Scholar] [CrossRef]
- Klejdus, B.; Lojkova, L.; Plaza, M.; Snoblova, M.; Sterbova, D. Hyphenated technique for the extraction and determination of isoflavones in algae: Ultrasound-assisted supercritical fluid extraction followed by fast chromatography with tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 7956–7965. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Sashima, T.; Hosokawa, M.; Miyashita, K. Comparative evaluation of growth inhibitory effect of stereoisomers of fucoxanthin in human cancer cell lines. J. Funct. Foods 2009, 1, 88–97. [Google Scholar] [CrossRef]
- Jaswir, I.; Noviendri, D.; Salleh, H.M.; Taher, M.; Miyashita, K.; Ramli, N. Analysis of Fucoxanthin Content and Purification of All-Trans-Fucoxanthin from Turbinaria turbinata and Sargassum plagyophyllum by SiO2 Open Column Chromatography and Reversed Phase-HPLC. J. Liq. Chromatogr. Relat. Technol. 2013, 36, 1340–1354. [Google Scholar] [CrossRef]
- Terasaki, M.; Hirose, A.; Narayan, B.; Baba, Y.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Evaluation of Recoverable Functional Lipid Components of Several Brown Seaweeds (Phaeophyta) from Japan with Special Reference to Fucoxanthin and Fucosterol Contents. J. Phycol. 2009, 45, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Kamogawa, H.; Susanto, E.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Seasonal variations of total lipids, fatty acid composition, and fucoxanthin contents of Sargassum horneri (Turner) and Cystoseira hakodatensis (Yendo) from the northern seashore of Japan. J. Appl. Phycol. 2013, 25, 1159–1169. [Google Scholar] [CrossRef]
- Kato, T.; Kumanireng, A.S.; Ichinose, I.; Kitahara, Y.; Kakinuma, Y.; Kato, Y. Structure and synthesis of active component from a marine alga, Sargassum tortile, which induces the settling of swimming larvae of Coryne uchidai. Chem. Lett. 1975, 4, 335–338. [Google Scholar] [CrossRef]
- Tsuchiya, N.; Sato, A.; Haruyama, H.; Watanabe, T.; Iijima, Y. Nahocols and isonahocols, endothelin antagonists from the brown alga, Sargassum autumnale. Phytochemistry 1998, 48, 1003–1011. [Google Scholar] [CrossRef]
- Reddy, P.; Urban, S. Meroditerpenoids from the southern Australian marine brown alga Sargassum fallax. Phytochemistry 2009, 70, 250–255. [Google Scholar] [CrossRef]
- Herath, K.H.I.N.M.; Kim, H.J.; Jang, J.H.; Kim, H.S.; Kim, H.J.; Jeon, Y.J.; Jee, Y. Mojabanchromanol Isolated from Sargassum horneri Attenuates Particulate Matter Induced Inflammatory Responses via Suppressing TLR2/4/7-MAPK Signaling in MLE-12 Cells. Mar. Drugs 2020, 18, 355. [Google Scholar] [CrossRef]
- Kim, H.S.; Wang, L.; Fernando, I.P.S.; Je, J.G.; Ko, S.C.; Kang, M.C.; Lee, J.M.; Yim, M.J.; Jeon, Y.J.; Lee, D.S. Antioxidant efficacy of (-)-loliolide isolated from Sargassum horneri against AAPH-induced oxidative damage in Vero cells and zebrafish models in vivo. J. Appl. Phycol. 2020, 32, 3341–3348. [Google Scholar] [CrossRef]
- Wang, L.; Kim, H.S.; Je, J.G.; Fu, X.T.; Huang, C.X.; Ahn, G.; Oh, J.Y.; Sanjeewa, K.K.A.; Xu, J.C.; Gao, X.; et al. In Vitro and In Vivo Photoprotective Effects of (-)-Loliode Isolated from the Brown Seaweed, Sargassum horneri. Molecules 2021, 26, 6898. [Google Scholar] [CrossRef]
- Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Jeon, Y.J.; Kim, H.S.; Fernando, I.P.S.; Ahn, G. (-)-Loliolide Isolated fromSargassum horneriProtects against Fine Dust-Induced Oxidative Stress in Human Keratinocytes. Antioxidants 2020, 9, 474. [Google Scholar] [CrossRef]
- Kim, J.A.; Ahn, B.N.; Kong, C.S.; Kim, S.K. The chromene sargachromanol E inhibits ultraviolet A-induced ageing of skin in human dermal fibroblasts. Brit. J. Dermatol. 2013, 168, 968–976. [Google Scholar] [CrossRef]
- Han, E.J.; Jayawardena, T.U.; Jang, J.H.; Fernando, I.P.S.; Jee, Y.; Jeon, Y.J.; Lee, D.S.; Lee, J.M.; Yim, M.J.; Wang, L.; et al. Sargachromenol Purified from Sargassum horneri Inhibits Inflammatory Responses via Activation of Nrf2/HO-1 Signaling in LPS-Stimulated Macrophages. Mar. Drugs 2021, 19, 497. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.J.; Cao, L.; Lee, B.; Gwon, W.G.; Park, S.H.; Kim, H.R. Sargahydroquinoic Acid, a Cyclooxygenase-2 Inhibitor, Attenuates Inflammatory Responses by Regulating NF-κB Inactivation and Nrf2 Activation in Lipopolysaccharide-Stimulated Cells. Inflammation 2021, 44, 2120–2131. [Google Scholar] [CrossRef]
- Iwashima, M.; Mori, J.; Ting, X.; Matsunaga, T.; Hayashi, K.; Shinoda, D.; Saito, H.; Sankawa, U.; Hayashi, T. Antioxidant and antiviral activities of plastoquinones from the brown alga Sargassum micracanthum, and a new chromene derivative converted from the plastoquinones. Biol. Pharm. Bull. 2005, 28, 374–377. [Google Scholar] [CrossRef] [Green Version]
- Mori, J.; Iwashima, M.; Wakasugi, H.; Saito, H.; Matsunaga, T.; Ogasawara, M.; Takahashi, S.; Suzuki, H.; Hayashi, T. New plastoquinones isolated from the brown alga, Sargassum micracanthum. Chem. Pharm. Bull. 2005, 53, 1159–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, S.H.; Cao, L.; Jeong, S.J.; Kim, H.R.; Nam, T.J.; Lee, S.G. The Comparison of Total Phenolics, Total Antioxidant, and Anti-Tyrosinase Activities of Korean Sargassum Species. J. Food Qual. 2021, 2021, 6640789. [Google Scholar] [CrossRef]
- Balboa, E.M.; Li, Y.X.; Ahn, B.N.; Eom, S.H.; Dominguez, H.; Jimenez, C.; Rodriguez, J. Photodamage attenuation effect by a tetraprenyltoluquinol chromane meroterpenoid isolated from Sargassum muticum. J. Photoch. Photobiol. B 2015, 148, 51–58. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, R.M.; Lin, X.P.; Liu, Y.H. Norisoprenoids from the Brown Alga Sargassum naozhouense Tseng et Lu. Molecules 2018, 23, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, S.; Tsutsumi, S.; Takada, Y.; Kimura, J. Antibacterial Quinone Metabolites from the Brown Alga, Sargassum sagamianum. B Chem. Soc. Jpn. 2008, 81, 1125–1130. [Google Scholar] [CrossRef]
- Lim, S.; Choi, A.H.; Kwon, M.; Joung, E.J.; Shin, T.; Lee, S.G.; Kim, N.G.; Kim, H.R. Evaluation of antioxidant activities of various solvent extract from Sargassum serratifolium and its major antioxidant components. Food Chem. 2019, 278, 178–184. [Google Scholar] [CrossRef]
- Lim, S.; Kwon, M.; Joung, E.J.; Shin, T.; Oh, C.W.; Choi, J.S.; Kim, H.R. Meroterpenoid-Rich Fraction of the Ethanolic Extract from Sargassum serratifolium Suppressed Oxidative Stress Induced by Tert-Butyl Hydroperoxide in HepG2 Cells. Mar. Drugs 2018, 16, 374. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Jang, K.H.; Kim, B.; Lee, B.H.; Choi, B.W.; Oh, K.B.; Shin, J. Meroditerpenoids from the Brown Alga Sargassum siliquastrum. J. Nat. Prod. 2008, 71, 1714–1719. [Google Scholar] [CrossRef]
- Lee, J.I.; Seo, Y. Chromanols from Sargassum siliquastrum and Their Antioxidant Activity in HT 1080 Cells. Chem. Pharm. Bull. 2011, 59, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.; Park, K.E.; Kim, Y.A.; Lee, H.J.; Yoo, J.S.; Ahn, J.W.; Lee, B.J. Isolation of tetraprenyltoluquinols from the brown alga Sargassum thunbergii. Chem. Pharm. Bull. 2006, 54, 1730–1733. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Kong, C.S.; Seo, Y.W.; Kim, S.K. Sargassum thunbergii extract inhibits MMP-2 and-9 expressions related with ROS scavenging in HT1080 cells. Food Chem. 2010, 120, 418–425. [Google Scholar] [CrossRef]
- Kim, S.O.; Choi, Y.H. Indole-6-Carboxaldehyde Isolated from Sargassum thunbergii (Mertens) Kuntze Prevents Oxidative Stress-Induced Cellular Damage in V79-4 Chinese Hamster Lung Fibroblasts through the Activation of the Nrf2/HO-1 Signaling Pathway. Cell Physiol. Biochem. 2020, 54, 959–974. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Heo, S.J.; Ko, S.C.; Park, W.S.; Choi, I.W.; Yi, M.; Jung, W.K. Indole-6-carboxaldehyde isolated from Sargassum thunbergii inhibits the expression and secretion of matrix metalloproteinase-9. Int. J. Mol. Med. 2019, 44, 1979–1987. [Google Scholar] [CrossRef]
- Maneesh, A.; Chakraborty, K. Previously undescribed fridooleanenes and oxygenated labdanes from the brown seaweed Sargassum wightii and their protein tyrosine phosphatase-1B inhibitory activity. Phytochemistry 2017, 144, 19–32. [Google Scholar] [CrossRef]
- Terasaki, M.; Kawagoe, C.; Ito, A.; Kumon, H.; Narayan, B.; Hosokawa, M.; Miyashita, K. Spatial and seasonal variations in the biofunctional lipid substances (fucoxanthin and fucosterol) of the laboratory-grown edible Japanese seaweed (Sargassum horneri Turner) cultured in the open sea. Saudi J. Biol. Sci. 2017, 24, 1475–1482. [Google Scholar] [CrossRef]
- Magura, J.; Moodley, R.; Jonnalagadda, S.B. Toxic metals (As and Pb) in Sargassum elegans Suhr (1840) and its bioactive compounds. Int. J. Environ. Health Res. 2019, 29, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Mulder, M.; Bogie, J.; Hoeks, C.; Schepers, M.; Tuabe, A.; Leijten, F.; Chintapakorn, Y.; Struik, D.; Liu, H.B.; Hellings, N.; et al. Dietary Sargassum fusiforme Improves Memory and Reduces Amyloid Plaque Load in an Alzheimer's Disease Mouse Model. Atherosclerosis 2019, 287, E57. [Google Scholar] [CrossRef]
- Ito, M.; Koba, K.; Hikihara, R.; Ishimaru, M.; Shibata, T.; Hatate, H.; Tanaka, R. Analysis of functional components and radical scavenging activity of 21 algae species collected from the Japanese coast. Food Chem. 2018, 255, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.L.; Liu, C.P.; Gao, L.; Lu, Y.B. One-Step Preparative Separation of Phytosterols from Edible Brown Seaweed Sargassum horneri by High-Speed Countercurrent Chromatography. Mar. Drugs 2019, 17, 691. [Google Scholar] [CrossRef] [Green Version]
- Malinin, N.L.; West, X.Z.; Byzova, T.V. Oxidation as "The Stress of Life". Aging-Us 2011, 3, 906–910. [Google Scholar] [CrossRef]
- Decker, E.A.; Elias, R.J.; McClements, D.J. Oxidation in Foods and Beverages and Antioxidant Applications: Management in Different Industry Sectors; Elsevier: Amsterdam, The Netherlands, 2010; Volume 2. [Google Scholar]
- Sadeer, N.B.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety—Chemistry, Applications, Strengths, and Limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- Chakraborty, K.; Maneesh, A.; Makkar, F. Antioxidant Activity of Brown Seaweeds. J. Aquat. Food Prod. Technol. 2017, 26, 406–419. [Google Scholar] [CrossRef]
- Wu, Y.J.; Gao, H.Q.; Wang, Y.X.; Peng, Z.T.; Guo, Z.Q.; Ma, Y.X.; Zhang, R.F.; Zhang, M.W.; Wu, Q.; Xiao, J.; et al. Effects of different extraction methods on contents, profiles, and antioxidant abilities of free and bound phenolics of Sargassum polycystum from the South China Sea. J. Food Sci. 2022, 87, 968–981. [Google Scholar] [CrossRef]
- Ismail, G.A.; Gheda, S.F.; Abo-shady, A.M.; Abdel-karim, O.H. In vitro potential activity of some seaweeds as antioxidants and inhibitors of diabetic enzymes. Food Sci. Technol. 2020, 40, 681–691. [Google Scholar] [CrossRef] [Green Version]
- Prasedya, E.S.; Martyasari, N.W.R.; Abidin, A.S.; Ilhami, B.T.K.; Padmi, H.; Widyastuti, S.; Sunarwidhi, A.L.; Sunarpi, H. Antioxidant activity of brown macroalgae Sargassum ethanol extract from Lombok coast, Indonesia. IOP Conf. Series Earth Environ. Sci. 2021, 712, 012038. [Google Scholar] [CrossRef]
- Taniguchi, R.; Ito, C.; Keitoku, S.; Miyake, Y.; Itoigawa, M.; Matsui, T.; Shibata, T. Analysis on the Structure of Phlorethols Isolated From the Warm-Temperate Brown Seaweed Sargassum carpophyllum and Their Antioxidant Properties. Nat. Prod. Commun. 2022, 17. [Google Scholar] [CrossRef]
- Saraswati; Giriwono, P.E.; Iskandriati, D.; Tan, C.P.; Andarwulan, N. In-vitro anti-inflammatory activity, free radical (DPPH) scavenging, and ferric reducing ability (FRAP) of Sargassum cristaefolium lipid-soluble fraction and putative identification of bioactive compounds using UHPLC-ESI-ORBITRAP-MS/MS. Food Res. Int. 2020, 137, 109702. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.W.S.; Tan, K.M.; Chew, L.Y.; Kong, K.W.; Yan, S.W. Application of Two-Level Full Factorial Design for the Extraction of Fucoxanthin and Antioxidant Activities from Sargassum siliquosum and Sargassum polycystum. J. Aquat. Food Prod. Technol. 2018, 27, 446–463. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Chelyn, L.J.; Vimala, S.; Fairulnizal, M.N.M.; Brownlee, I.A.; Amin, I. Carotenoid composition and antioxidant potential of Eucheuma denticulatum, Sargassum polycystum and Caulerpa lentillifera. Heliyon 2020, 6, e04654. [Google Scholar] [CrossRef]
- Yan, X.J.; Chuda, Y.; Suzuki, M.; Nagata, T. Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci. Biotechnol. Biochem. 1999, 63, 605–607. [Google Scholar] [CrossRef]
- Savira, A.D.R.; Amin, M.N.G.; Alamsjah, M.A. The effect of different type of solvents on the antioxidant activity of fucoxanthin extract from brown seaweed Sargassum duplicatum. IOP Conf. Series Earth Environ. Sci. 2021, 718, 012010. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Raji, V.; Loganathan, C.; Sadhasivam, G.; Kandasamy, S.; Poomani, K.; Thayumanavan, P. Purification of fucoxanthin from Sargassum wightii Greville and understanding the inhibition of angiotensin 1-converting enzyme: An in vitro and in silico studies. Int. J. Biol. Macromol. 2020, 148, 696–703. [Google Scholar] [CrossRef]
- Ham, Y.M.; Kim, K.N.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Chemical Constituents from Sargassum micracanthum and Antioxidant Activity. Int. J. Pharmacol. 2010, 6, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Lee, H.J.; Park, K.E.; Kim, Y.A.; Ahn, J.W.; Jong, S.Y.; Lee, B.J. Peroxynitrite-scavenging constituents from the brown alga Sargassum thunbergii. Biotechnol. Bioprocess Eng. 2004, 9, 212–216. [Google Scholar] [CrossRef]
- Jang, K.H.; Lee, B.H.; Choi, B.W.; Lee, H.S.; Shin, J. Chromenes from the brown alga Sargassum siliquastrum. J. Nat. Prod. 2005, 68, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Cho, J.Y.; Kang, S.E.; Hong, Y.K.; Ahn, D.H. Antioxidant activity of mojabanchromanol, a novel chromene, isolated from brown alga Sargassum siliquastrum. J. Environ. Biol. 2008, 29, 479–484. [Google Scholar] [PubMed]
- Farvin, K.H.S.; Surendraraj, A.; Al-Ghunaim, A.; Al-Yamani, F. Chemical profile and antioxidant activities of 26 selected species of seaweeds from Kuwait coast. J. Appl. Phycol. 2019, 31, 2653–2668. [Google Scholar] [CrossRef]
- Savaghebi, D.; Barzegar, M.; Mozafari, M.R. Manufacturing of nanoliposomal extract from Sargassum boveanum algae and investigating its release behavior and antioxidant activity. Food Sci. Nutr. 2020, 8, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Sobuj, M.K.A.; Islam, M.A.; Haque, M.A.; Islam, M.M.; Alam, M.J.; Rafiquzzaman, S.M. Evaluation of bioactive chemical composition, phenolic, and antioxidant profiling of different crude extracts of Sargassum coriifolium and Hypnea pannosa seaweeds. J. Food Meas. Charact. 2021, 15, 1653–1665. [Google Scholar] [CrossRef]
- Vasconcelos, J.B.; de Vasconcelos, E.R.T.P.P.; Urrea-Victoria, V.; Bezerra, P.S.; Reis, T.N.V.; Cocentino, A.L.M.; Navarro, D.M.A.F.; Chow, F.; Areces, A.J.; Fujii, M.T. Antioxidant activity of three seaweeds from tropical reefs of Brazil: Potential sources for bioprospecting. J. Appl. Phycol. 2019, 31, 835–846. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, H.J.; Jee, Y.; Jeon, Y.J.; Kim, H.J. Antioxidant potential of Sargassum horneri extract against urban particulate matter-induced oxidation. Food Sci. Biotechnol. 2020, 29, 855–865. [Google Scholar] [CrossRef]
- Chen, C.J.; Han, L.S.; Yao, M.K.; Shi, L.; Zhang, M.S.; Shi, Y.P.; Liu, C.H.; Bai, X.F.; Liu, X.; Liu, X.; et al. Comparative Studies on Antioxidant, Angiotensin-Converting Enzyme Inhibitory and Anticoagulant Activities of the Methanol Extracts from Two Brown Algae (Sargassum horneri and Sargassum thunbergii). Russ. J. Mar. Biol. 2021, 47, 380–387. [Google Scholar] [CrossRef]
- Kim, H.S.; Sanjeewa, K.K.A.; Fernando, I.P.S.; Ryu, B.; Yang, H.W.; Ahn, G.; Kang, M.C.; Heo, S.T.; Je, J.G.; Jeon, Y.J. A comparative study of Sargassum horneri Korea and China strains collected along the coast of Jeju Island South Korea: Its components and bioactive properties. Algae 2018, 33, 341–349. [Google Scholar] [CrossRef]
- Abu-Khudir, R.; Ismail, G.A.; Diab, T. Antimicrobial, Antioxidant, and Anti-Tumor Activities of Sargassum linearifolium and Cystoseira crinita from Egyptian Mediterranean Coast. Nutr. Cancer 2021, 73, 829–844. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhang, Y.; Yuan, Y.; Farooq, M.A.; Fayyaz, M.S.; Su, D.X.; Zeng, Q.Z.; Rahaman, A. Process optimization and antioxidative activity of polyphenols derived from different seaweed species Sargassum miyabei, Undaria pinnatifida Suringar, and Sargassum thunbergii. Food Sci. Nutr. 2022, 10, 2021–2028. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Rodrigues, C.; Garcia-Oliveira, P.; Lourenco-Lopes, C.; Silva, S.A.; Garcia-Perez, P.; Carvalho, A.P.; Domingues, V.F.; Barroso, M.F.; Delerue-Matos, C.; et al. Screening of Bioactive Properties in Brown Algae from the Northwest Iberian Peninsula. Foods 2021, 10, 1915. [Google Scholar] [CrossRef]
- Terme, N.; Boulho, R.; Kucma, J.P.; Bourgougnon, N.; Bedoux, G. Radical scavenging activity of lipids from seaweeds isolated by solid-liquid extraction and supercritical fluids. Ocl 2018, 25, D505. [Google Scholar] [CrossRef]
- Le Lann, K.; Jegou, C.; Stiger-Pouvreau, V. Effect of different conditioning treatments on total phenolic content and antioxidant activities in two Sargassacean species: Comparison of the frondose Sargassum muticum (Yendo) Fensholt and the cylindrical Bifurcaria bifurcata R. Ross. Phycol. Res. 2008, 56, 238–245. [Google Scholar] [CrossRef]
- Sanchez-Camargo, A.D.; Montero, L.; Stiger-Pouvreau, V.; Tanniou, A.; Cifuentes, A.; Herrero, M.; Ibanez, E. Considerations on the use of enzyme-assisted extraction in combination with pressurized liquids to recover bioactive compounds from algae. Food Chem. 2016, 192, 67–74. [Google Scholar] [CrossRef]
- Nazarudin, M.F.; Paramisparam, A.; Khalid, N.A.; Albaz, M.N.; Shahidan, M.S.; Yasin, I.S.M.; Isha, A.; Abu Zarin, M.; Aliyu-Paiko, M. Metabolic variations in seaweed, Sargassum polycystum samples subjected to different drying methods via 1H NMR-based metabolomics and their bioactivity in diverse solvent extracts. Arab. J. Chem. 2020, 13, 7652–7664. [Google Scholar] [CrossRef]
- Vasanthi, C.; Rao, V.A.; Babu, R.N.; Sriram, P.; Karunakaran, R. In-vitro antioxidant activities of aqueous and alcoholic extracts of Sargassum species-Indian brown seaweed. J. Food Process. Preserv. 2020, 44, e14877. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Optimum conditions of microwave-assisted extraction for phenolic compounds and antioxidant capacity of the brown alga Sargassum vestitum. Sep. Sci. Technol. 2018, 53, 1711–1723. [Google Scholar] [CrossRef]
- Kosanic, M.; Rankovic, B.; Stanojkovic, T. Brown macroalgae from the Adriatic Sea as a promising source of bioactive nutrients. J. Food Meas. Charact. 2019, 13, 330–338. [Google Scholar] [CrossRef]
- Santos, J.P.; Torres, P.B.; dos Santos, D.Y.A.C.; Motta, L.B.; Chow, F. Seasonal effects on antioxidant and anti-HIV activities of Brazilian seaweeds. J. Appl. Phycol. 2019, 31, 1333–1341. [Google Scholar] [CrossRef]
- Chouh, A.; Nouadri, T.; Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phlorotannins of the Brown Algae Sargassum vulgare from the Mediterranean Sea Coast. Antioxidants 2022, 11, 1055. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.R.G.; Paul, P.T.; Anas, K.K.; Tejpal, C.S.; Chatterjee, N.S.; Anupama, T.K.; Geethalakshmi, V.; Anandan, R.; Jayarani, R.; Mathew, S. Screening of effective solvents for obtaining antioxidant-rich seaweed extracts using principal component analysis. J. Food Process. Preserv. 2020, 44, e14716. [Google Scholar] [CrossRef]
- Vijayan, R.; Chitra, L.; Penislusshiyan, S.; Palvannan, T. Exploring bioactive fraction of Sargassum wightii: In vitro elucidation of angiotensin-I-converting enzyme inhibition and antioxidant potential. Int. J. Food Prop. 2018, 21, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Pinteus, S.; Lemos, M.F.L.; Silva, J.; Alves, C.; Neugebauer, A.; Freitas, R.; Duarte, A.; Pedrosa, R. An Insight into Sargassum muticum Cytoprotective Mechanisms against Oxidative Stress on a Human Cell In Vitro Model. Mar. Drugs 2017, 15, 353. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.S.; Ranasinghe, P.; Gunasekara, U.K.D.S.S.; Jeon, Y.J. Antioxidant and anti-inflammatory functionality of ten Sri Lankan seaweed extracts obtained by carbohydrase assisted extraction. Food Sci. Biotechnol. 2018, 27, 1761–1769. [Google Scholar] [CrossRef]
- Sprygin, V.G.; Kushnerova, N.F.; Fomenko, S.E.; Drugova, E.S.; Lesnikova, L.N.; Merzlyakov, V.Y.; Momot, T.V. The Influence of an Extract from the Marine Brown Alga Sargassum pallidum on the Metabolic Reactions in the Liver under Experimental Toxic Hepatitis. Russ. J. Mar. Biol. 2017, 43, 479–484. [Google Scholar] [CrossRef]
- Altinok-Yipel, F.; Tekeli, I.O.; Ozsoy, S.Y.; Guvenc, M.; Sayin, S.; Yipel, M. Investigation of hepatoprotective effect of some algae species on carbon tetrachloride-induced liver injury in rats. Arch. Physiol. Biochem. 2020, 126, 463–467. [Google Scholar] [CrossRef]
- Zhao, Z.L.; Yang, X.Q.; Gong, Z.Q.; Pan, M.Z.; Han, Y.L.; Liu, Y. Antioxidant activities of crude phlorotannins from Sargassum hemiphyllum. J. Huazhong Univ. Sci. Technol. 2016, 36, 449–455. [Google Scholar] [CrossRef]
- Heo, S.J.; Ko, S.C.; Kang, S.M.; Kang, H.S.; Kim, J.P.; Kim, S.H.; Lee, K.W.; Cho, M.G.; Jeon, Y.J. Cytoprotective effect of fucoxanthin isolated from brown algae Sargassum siliquastrum against H2O2-induced cell damage. Eur. Food Res. Technol. 2008, 228, 145–151. [Google Scholar] [CrossRef]
- Piao, M.J.; Yoon, W.J.; Kang, H.K.; Yoo, E.S.; Koh, Y.S.; Kim, D.S.; Lee, N.H.; Hyun, J.W. Protective Effect of the Ethyl Acetate Fraction of Sargassum muticum Against Ultraviolet B-Irradiated Damage in Human Keratinocytes. Int. J. Mol. Sci. 2011, 12, 8146–8160. [Google Scholar] [CrossRef]
- Han, E.J.; Kim, S.Y.; Han, H.J.; Kim, H.S.; Kim, K.N.; Fernando, I.P.S.; Madusanka, D.M.D.; Dias, M.K.H.M.; Cheong, S.H.; Park, S.R.; et al. UVB protective effects of Sargassum horneri through the regulation of Nrf2 mediated antioxidant mechanism. Sci. Rep. 2021, 11, 9963. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Chen, H.H.; Qu, H.D.; Qiao, K.; Xu, M.; Wu, J.N.; Su, Y.C.; Shi, Y.; Liu, Z.Y.; Wang, Q. Photoprotective effects of Sargassum thunbergii on ultraviolet B-induced mouse L929 fibroblasts and zebrafish. BMC Complement. Med. 2022, 22, 144. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Jeon, Y.J. Protective effect of fucoxanthin isolated from Sargassum siliquastrum on UV-B induced cell damage. J. Photochem. Photobiol. B Biol. 2009, 95, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Semaida, A.I.; El-Khashab, M.A.; Saber, A.A.; Hassan, A.I.; Elfouly, S.A. Effects of Sargassum virgatum extracts on the testicular measurements, genomic DNA and antioxidant enzymes in irradiated rats. Int. J. Radiat. Biol. 2022, 98, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.T.; Sudirman, S.; Hsieh, M.C.; Hu, J.Y.; Kong, Z.L. Oral supplementation of fucoxanthin-rich brown algae extract ameliorates cisplatin-induced testicular damage in hamsters. Biomed. Pharmacother. 2020, 125, 109992. [Google Scholar] [CrossRef]
- Kim, H.J.; Herath, K.H.I.N.M.; Dinh, D.T.T.; Kim, H.S.; Jeon, Y.J.; Kim, H.J.; Jee, Y. Sargassum horneri ethanol extract containing polyphenols attenuates PM-induced oxidative stress via ROS scavenging and transition metal chelation. J. Funct. Foods 2021, 79, 104401. [Google Scholar] [CrossRef]
- Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, H.S.; Jeon, Y.J.; Jee, Y.; Kim, K.N.; Lee, K.; Fernando, I.P.S.; Ahn, G. Sargassum horneri (Turner) C. Agardh ethanol extract attenuates fine dust-induced inflammatory responses and impaired skin barrier functions in HaCaT keratinocytes. J. Ethnopharmacol. 2021, 273, 114003. [Google Scholar] [CrossRef]
- Dai, Y.L.; Jiang, Y.F.; Lu, Y.A.; Yu, J.B.; Kang, M.C.; Jeon, Y.J. Fucoxanthin-rich fraction from Sargassum fusiformis alleviates particulate matter-induced inflammation in vitro and in vivo. Toxicol. Rep. 2021, 8, 349–358. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Kim, H.S.; Lee, W.W.; Vaas, A.P.J.P.; De Silva, H.I.C.; Abayaweera, G.S.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, D.S.; et al. Beijing urban particulate matter-induced injury and inflammation in human lung epithelial cells and the protective effects of fucosterol from Sargassum binderi (Sonder ex J. Agardh). Environ. Res. 2019, 172, 150–158. [Google Scholar] [CrossRef]
- Salari, Z.; Abbasnejad, M.; Hesni, M.A.; Esmaeili-Mahani, S. Effect of the Sargassum angustifolium Extract on Methamphetamine-Induced Cytotoxicity in SH-SY5Y Cells. Altern. Med. 2022, 2022, 9978235. [Google Scholar] [CrossRef]
- Wei, J.X.; Mou, C.Y.; Bao, Y.J.; Xie, Y.F.; Jin, H.X.; Shen, H.W.; Zhou, W.H.; Zhang, J.R.; He, S.; Chen, B.J.; et al. Fucoxanthin alleviates methamphetamine-induced neurotoxicity possibly via the inhibition of interaction between Keap1 and Nrf2. J. Funct. Foods 2021, 86, 104713. [Google Scholar] [CrossRef]
- Shahriari, A.G.; Mohkami, A.; Niazi, A.; Parizipour, M.H.G.; Habibi-Pirkoohi, M. Application of Brown Algae (Sargassum angustifolium) Extract for Improvement of Drought Tolerance in Canola (Brassica napus L.). Iran. J. Biotechnol. 2021, 19, 22–29. [Google Scholar] [CrossRef]
- Latique, S.; Elouaer, M.; Souguir, M.; Aloui, H.; Hannachi, C.; Chernane, H.; Mansori, M.; Elkaoua, M. Effect of seaweed extract of Sargassum vulgare on germination behavior of two bean cultivars (Phaseolus vulgaris L) under salt stress. IOSR J. Agric. Vet. Sci. 2014, 7, 116–120. [Google Scholar] [CrossRef]
- Han, S.; Park, J.S.; Umanzor, S.; Yarish, C.; Kim, J.K. Effects of extraction methods for a new source of biostimulant from Sargassum horneri on the growth of economically important red algae, Neopyropia yezoensis. Sci. Rep. 2022, 12, 11878. [Google Scholar] [CrossRef] [PubMed]
- Khedia, J.; Dangariya, M.; Nakum, A.K.; Agarwal, P.; Panda, A.; Parida, A.K.; Gangapur, D.R.; Meena, R.; Agarwal, P.K. Sargassum seaweed extract enhances Macrophomina phaseolina resistance in tomato by regulating phytohormones and antioxidative activity. J. Appl. Phycol. 2020, 32, 4373–4384. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, H.A.; Han, J.S. Sargassum sagamianum extract protects INS-1 pancreatic β cells against high glucose-induced apoptosis. Cytotechnology 2019, 71, 389–399. [Google Scholar] [CrossRef]
- Wu, S.Y.; Zuo, J.H.; Cheng, Y.; Zhang, Y.; Zhang, Z.S.; Wu, M.J.; Yang, Y.; Tong, H.B. Ethanol extract of Sargarsum fusiforme alleviates HFD/STZ-induced hyperglycemia in association with modulation of gut microbiota and intestinal metabolites in type 2 diabetic mice. Food Res. Int. 2021, 147, 110550. [Google Scholar] [CrossRef]
- Oliyaei, N.; Moosavi-Nasab, M.; Tamaddon, A.M.; Tanideh, N. Antidiabetic effect of fucoxanthin extracted from Sargassum angustifolium on streptozotocin-nicotinamide-induced type 2 diabetic mice. Food Sci. Nutr. 2021, 9, 3521–3529. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef]
- Guadalajara, U.O. Effect of Fucoxanthin on the Metabolic Syndrome, Insulin Sensitivity and Insulin Secretion. Available online: https://ClinicalTrials.gov/show/NCT03613740 (accessed on 30 September 2022).
- Matsui, T.; Ito, C.; Itoigawa, M.; Shibata, T. Three phlorotannins from Sargassum carpophyllum are effective against the secretion of allergic mediators from antigen-stimulated rat basophilic leukemia cells. Food Chem. 2022, 377, 131992. [Google Scholar] [CrossRef] [PubMed]
- Safaeian, L.; Yegdaneh, A.; Mobasherian, M. The Effect of Sargassum angustifolium Ethanol Extract on Cadmium Chloride-Induced Hypertension in Rat. Res. J. Pharmacogn. 2021, 8, 81–89. [Google Scholar] [CrossRef]
- Yegdaneh, A.; Safaeian, L.; Halvaei-Varnousfaderani, M.; Bazvand, S. Antihyperlipidemic and Antioxidant Effects of Ethanol Fraction of Sargassum angustifolium in Dexamethasone-Induced Dyslipidemic Rats. Res. J. Pharmacogn. 2022, 9, 39–49. [Google Scholar] [CrossRef]
- Ko, W.; Lee, H.; Kim, N.; Jo, H.G.; Woo, E.R.; Lee, K.; Han, Y.S.; Park, S.R.; Ahn, G.; Cheong, S.H.; et al. The Anti-Oxidative and Anti-Neuroinflammatory Effects of Sargassum horneri by Heme Oxygenase-1 Induction in BV2 and HT22 Cells. Antioxidants 2021, 10, 859. [Google Scholar] [CrossRef] [PubMed]
- Sengkhim, R.; Peerakietkhajorn, S.; Jeanmard, N.; Pongparadon, S.; Khuituan, P.; Thitiphatphuvanon, T.; Surinlert, P.; Tipbunjong, C. Effects of Sargassum plagiophyllum extract pretreatment on tissue histology of constipated mice. Trop. J. Pharm. Res. 2021, 20, 2339–2346. [Google Scholar] [CrossRef]
- Kirindage, K.G.I.S.; Jayasinghe, A.M.K.; Han, E.J.; Jee, Y.; Kim, H.J.; Do, S.G.; Fernando, I.P.S.; Ahn, G. Fucosterol Isolated from Dietary Brown Alga Sargassum horneri Protects TNF-α/IFN-γ-Stimulated Human Dermal Fibroblasts Via Regulating Nrf2/HO-1 and NF-κB/MAPK Pathways. Antioxidants 2022, 11, 1429. [Google Scholar] [CrossRef]
- Choi, K.S.; Shin, T.S.; Chun, J.; Ahn, G.; Han, E.J.; Kim, M.J.; Kim, J.B.; Kim, S.H.; Kho, K.H.; Kim, D.H.; et al. Sargahydroquinoic acid isolated from Sargassum serratifolium as inhibitor of cellular basophils activation and passive cutaneous anaphylaxis in mice. Int. Immunopharmacol. 2022, 105, 108567. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Chandrasekaran, N.; Mukherjee, A.; Thomas, J. Protective efficacy of microencapsulated seaweed extracts for preventing Aeromonas infections in Oreochromis mossambicus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 218, 36–45. [Google Scholar] [CrossRef]
- Dousip, A.; Matanjun, P.; Sulaiman, M.R.; Tan, T.S.; Ooi, Y.B.H.; Lim, T.P. Effect of seaweed mixture intake on plasma lipid and antioxidant profile of hyperholesterolaemic rats. J. Appl. Phycol. 2014, 26, 999–1008. [Google Scholar] [CrossRef]
- Ibrahim, R.Y.M.; Hammad, H.B.I.; Gaafar, A.A.; Saber, A.A. The possible role of the seaweed Sargassum vulgare as a promising functional food ingredient minimizing aspartame-associated toxicity in rats. Int. J. Environ. Health Res. 2022, 32, 752–771. [Google Scholar] [CrossRef]
- He, W.; Sun, D. A Kind of Health Care Complex Tea and Preparation Method Thereof. China Patent CN104222378B, 17.08.2016, 17 August 2016. [Google Scholar]
- Practical and Creative Uses of Sargassum Seaweed in the Caribbean. Available online: https://www.villapalmarcancun.com/blog/destination/practical-and-creative-uses-of-sargassum-seaweed-in-the-caribbean (accessed on 5 January 2023).
- Prabhasankar, P.; Ganesan, P.; Bhaskar, N. Influence of Indian Brown Seaweed (Sargassum marginatum) as an Ingredient on Quality, Biofunctional, and Microstructure Characteristics of Pasta. Food Sci. Technol. Int. 2009, 15, 471–479. [Google Scholar] [CrossRef]
- Kim, M.-J.; Kim, K.-B.-W.-R.; Lee, C.-J.; Kwak, J.-H.; Kim, D.-H.; SunWoo, C.; Jung, S.-A.; Kang, J.-Y.; Kim, H.-J.; Choi, J.-S. Effect of Sargassum sagamianum extract on shelf-life and improved quality of morning bread. Korean J. Food Sci. Technol. 2011, 43, 723–728. [Google Scholar] [CrossRef]
- Kim, M.-J.; Song, E.-J.; Kim, K.-B.-W.-R.; Lee, C.-J.; Jung, J.-Y.; Kwak, J.-H.; Choi, M.-K.; Kim, D.-H.; SunWoo, C.; Choi, J.-S. Effect of Sargassum fulvellum extracts on shelf-life and quality improvement of bread. J. Korean Soc. Food Sci. Nutr. 2011, 40, 867–874. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Kim, K.-B.-W.-R.; Song, E.-J.; Kim, J.-H.; Kim, A.-R.; Kim, M.-J.; Moon, J.-H.; Kang, H.-M.; Lee, H.-D.; Hong, Y.-K. Effect of extracts from Sargassum siliquastrum on shelf-life and quality of bread. J. Korean Soc. Food Sci. Nutr. 2008, 37, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Gan, L.J.; You, Q.; Luo, Y.M.; Ye, Y.T.; Lei, L.; Deng, Z.Y.; Rong, H. Effect of superfine grinding Sargassum fusiforme residue powder on sponge cakes properties. LWT Food Sci. Technol. 2022, 165, 113735. [Google Scholar] [CrossRef]
- Miyashita, K.; Abe, M. Fucoxanthin-Enriched Algae Product and Method for Producing the Same. Japan Patent JP2013031369A, 14 February 2013. [Google Scholar]
- Zhang, T.; Fu, S.; Wang, Y.; Xue, C.; Mao, X.; Xue, Y.; Wang, J.; Chang, Y.; Tang, Q.; Jiang, X. Application of Fucoxanthin and/or Fucoxanthin in Preparation for Improving DHA Level of Human Body. China Patent CN112674337A, 20 April 2021. [Google Scholar]
- Aditya, N.W.; Amin, M.N.G.; Alamsjah, M.A. The effect of fucoxanthin as coloring agent on the quality of catfish sausage. IOP Conf. Ser. Earth Environ. Sci. 2020, 441, 012080. [Google Scholar] [CrossRef]
- Redmond, S.; Kim, J.K.; Yarish, C.; Pietrak, M.; Bricknell, I. Culture of Sargassum in Korea: Techniques and Potential for Culture in the US; The University of Maine: Orono, ME, USA, 2014. [Google Scholar]
- Lee, G.S.; Lee, D.H.; Ko, H.J.; Kim, K.B.; Kwon, S.W.; Pyo, H.B.; Lee, W.J.; Hyun, C.G. Sargassum sp. Extract that Have Low Photo-Induced Cytotoxicity, Preparation Method Thereof and Cosmetic Composition Containing the Same. Republic of Korea Patent KR1020110006444B1, 19 May 2011. [Google Scholar]
- He, M. Face Mask made of Seawead. China Patent CN1147374A, 16 April 1997. [Google Scholar]
- Cai, C.; Yang, Y.; Zheng, Y.; Hu, Y.; He, P.; Jia, R.; Wu, W.; Wu, H.; Zhou, L.; Zhang, Z. Sargassum horneri Boiled Alga Clay Mask and Preparation Method Thereof. China Patent CN103720619B, 16 April 2014. [Google Scholar]
- Li, X. Seaweed Mud Non-Woven Mask and Preparation Method Thereof. China Patent CN103876980A, 25 June 2014. [Google Scholar]
- Yang, L. Nano-Collagen Moistening Skin-Care Facial Mask. China Patent CN104323975A, 4 February 2015. [Google Scholar]
- Ltd., Q.A.M.D.C. Seaweed Paste Mask. China Patent CN103690464A, 2 April 2014. [Google Scholar]
- Mei, B.C.; Lyga, J.W. Sargassum muticum Extracts and Methods of Use. WO2014158854A1, 2 October 2014. [Google Scholar]
- Kim, H.R.; Im, K.T.; Im, S.J. Methods for the Preparation of Nano-emulsion Containing Skin-Whitening, Anti-Wrinkle and Anti-Inflammatory Cosmetic Composition and Use Thereof. Republic of Korea Patent KR1020210073174B1, 18 January 2022. [Google Scholar]
- Breton, L.; Girerd, F.; Renault, B. Use of Phloroglucinol in a Composition. Canada Patent CA2255222C, 16 December 2018. [Google Scholar]
- Decoder, I. Phloroglucinol Trimethyl Ether. Available online: https://incidecoder.com/ingredients/phloroglucinol-trimethyl-ether (accessed on 6 October 2022).
- Ismay, D. Some Colour Reactions of Phloroglucinol. J. Soc. Chem. Ind. 1950, 69, 58–60. [Google Scholar] [CrossRef]
- Andersen, F.A. Final report on the safety assessment of phloroglucinol. J. Am. Coll. Toxicol. 1995, 14, 468–475. [Google Scholar]
- Sipahutar, Y.H.; Albaar, N.; Purnamasari, H.B.; Kristiany, M.G.; Prabowo, D.H.G. Seaweed extract (Sargassum polycystum) as a preservative on sunscreen cream with the addition of seaweed porridge. IOP Conf. Ser. Earth Environ. Sci. 2019, 278, 012072. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, M.; Andre, G.; Pellegrini, L. Cosmetic or Dermatological Compositions, e.g. for Combating Skin Aging, Containing Extract of Sargassum Muticum Brown Algae Having e.g. Antiradical and DNA Protecting Action. France Patent FR2838342A1, 17 June 2005. [Google Scholar]
- Shin, Y.-S.; Shin, C.-W.; Shin, Y.-W.; Park, Y.-H. Cosmetic Composition Including Extracts of Sargassum Horneri and Enteromorpha Prolifera. Republic of Korea Patent KR20190087199A, 24 July 2019. [Google Scholar]
- Mansauda Karlah Lifie, R.; Effionora, A.; Tati, N. Antioxidant and Anti-Collagenase Activity of Sargassum Plagyophyllum Extract as an Anti-Wrinkle Cosmetic Ingredient. Pharmacogn. J. 2018, 10, 932–936. [Google Scholar] [CrossRef] [Green Version]
- Gazali, M. Aktivitas Inhibitor Tirosinase Pada Ekstrak Alga Cokelat Sargassum Sp. Agardh Asal Pesisir Lhok Bubon, Kabu-paten Aceh Barat. J. Perikan. Terpadu 2018, 1, 26–40. [Google Scholar] [CrossRef]
- Biocogent. DermalRx. Available online: https://www.biocogent.com/dermalrx (accessed on 5 January 2023).
- Decoder, I. Fucoxanthin. Available online: https://incidecoder.com/ingredients/fucoxanthin (accessed on 6 October 2022).
- Hwang, Y. Anti-Obesity Cream Composition Containing Fucoxanthin. WO2011152692A2, 8 December 2011. [Google Scholar]
- Sembera, J.A.; Meier, E.J.; Waliczek, T.M. Composting as an Alternative Management Strategy for Sargassum Drifts on Coastlines. Horttechnology 2018, 28, 80–84. [Google Scholar] [CrossRef]
- Mendoza-Morales, L.T.; Mendoza-González, A.C.; Mateo-Cid, L.E.; Rodríguez-Dorantes, A. Analysis of the effect as biostimulants of Sargassum vulgare and Ulva fasciata extracts on Lens esculenta growth. Mex. J. Biotechnol. 2019, 4, 15–28. [Google Scholar] [CrossRef]
- Latique, S.; Elouaer, M.A.; Chernane, H.; Hannachi, C.; Elkaoua, M. Effect of seaweed liquid extract of Sargassum vulgare on growth of durum wheat seedlings (Triticum durum L.) under salt stress. Int. J. Innov. Appl. Stud. 2014, 7, 1430. [Google Scholar]
- Sbaihat, L.; Takeyama, K.; Koga, T.; Takemoto, D.; Kawakita, K. Induced Resistance in Solanum lycopersicum by Algal Elicitor Extracted from Sargassum fusiforme. Sci. World J. 2015, 2015, 870520. [Google Scholar] [CrossRef] [Green Version]


| Species | Meroterpenoids | Ref. |
|---|---|---|
| S. autumnale | Nahocol (Isomers: A; A1; B; C; D1; D2) Isonahocol (Isomers D1; D2) | [41] |
| S. fallax | Fallahydroquinone Fallaquinone Fallachromenoic acid | [42] |
| S. horneri | Mojabanchromanol | [43] |
| (−)-Loliolide | [44,45,46] | |
| Sargachromanol E | [47] | |
| Sargachromenol | [48] | |
| S. macrcarpum | Sargahydroquinoic acid | [49] |
| S. micracathum | 2-geranylgeranyl-6-methylbenzoquinone | [50] |
| 2-geranylgeranyl-6-methyl-1,4-benzohydroquinone | [51] | |
| S. miyabei | Sargahydroquinoic acid Sargachromanol | [52] |
| S. muticum | Tetraprenyltoluquinol chromane meroterpenoid | [53] |
| S. naozhouense | Sargassumone (2R,6S,8S,9S)-hexahydro-2,9-dihydroxy-4,4,8-trimethyl-6-acetyloxy-3(2H)-benzofuranone (6S,8S,9R)-hexahydro-6,9-dihydroxy-4,4,8-trimethyl-2(2H)-benzofuranone Loliolide (+)-Epiloliolide Spheciospongones A (+)-Kjellmanianone | [54] |
| S. sagamianum | Chromequinolide 11′-Hydroxysargachromelide 15′-Hydroxysargaquinolide 15′- Methylenesargaquinolide (2′E,5′E)-2-Methyl-6-(7′-oxo-3′-methylocta-2′,5′-dienyl)-1,4-benzoquinone | [55] |
| S. serrifolium | Sargahydroquinoic acid Sargachromanol | [52,56,57] |
| Sargahydroquinoic acid | [49] | |
| Sargaquinoic acid | [56] | |
| S. siliquastrum | Sargahydroquinoic acid Isonahocol (Isomers D1; D2) Nahocol (Isomers A; A1; D1; D2) | [58] |
| Sargachromenol (Isomer D; E; K) 13-(3,4-dihydro-6-hydroxy-2,8-dimethy2H-1-benzopyran-2-yl)-2,6,10-trimethyl-trideca-(2E,6E)- diene-4,5,10-triol 9-(3,4-dihydro-6-hydroxy-2,8-dimethy-2H-1-benzopyran-2- yl)-2,6-dimethyl-(6E)-nonenoic acid | [59] | |
| S. thunbergii | Thumbergol (Isomer A; B) | [60] |
| Sargachromanol (Isomer E; D) Sargahydroquinoic acid | [61] | |
| Indole-6-Carboxaldehyde | [62,63] | |
| S. tortile | δ-Tocotrienol, δ -Tocotrienol; 11′,12′ (+)-Epoxide | [40] |
| S. wightii | 2(α)-hydroxy-(28,29)-frido-olean-12(13),21(22)-dien-20-propyl-21- hex-4′ (Z)-enoate; 2(α)-hydroxy-(28, 29)-frido-olean-12(13), 21(22)-dien-20-prop-2(E)-en-21-butanoate; 2α-hydroxy-8(17), 12 E, 14- labdatriene; 3β, 6β, 13α-tri hydroxy 8(17), 12E, 14-labdatriene | [64] |
| Species | Phytosterol | Reference |
| S. elegans | β-Sitosterol;Fucosterol | [66] |
| S. fusiforme | 24(S)-Saringosterol; Ergosterol; Fucosterol; Cholesterol; β-Sitosterol | [67,68] |
| S. horneri | Fucosterol; Saringosterol | [65,69] |
| S. lacerifolium | β-Sitosterol | |
| S. piluliferum | Ergosterol; Fucosterol; Cholesterol; | [68] |
| S. thunbergii | Ergosterol; Fucosterol; Cholesterol; | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catarino, M.D.; Silva-Reis, R.; Chouh, A.; Silva, S.; Braga, S.S.; Silva, A.M.S.; Cardoso, S.M. Applications of Antioxidant Secondary Metabolites of Sargassum spp. Mar. Drugs 2023, 21, 172. https://doi.org/10.3390/md21030172
Catarino MD, Silva-Reis R, Chouh A, Silva S, Braga SS, Silva AMS, Cardoso SM. Applications of Antioxidant Secondary Metabolites of Sargassum spp. Marine Drugs. 2023; 21(3):172. https://doi.org/10.3390/md21030172
Chicago/Turabian StyleCatarino, Marcelo D., Rita Silva-Reis, Amina Chouh, Sónia Silva, Susana S. Braga, Artur M. S. Silva, and Susana M. Cardoso. 2023. "Applications of Antioxidant Secondary Metabolites of Sargassum spp." Marine Drugs 21, no. 3: 172. https://doi.org/10.3390/md21030172