
Morphological and Physiological Aspects of Mutable Collagenous Tissue at the Autotomy Plane of the Starfish Asterias rubens L. (Echinodermata, Asteroidea): An Echinoderm Paradigm

Abstract
:1. Introduction
Taxonomic Note
2. Morphological Aspects
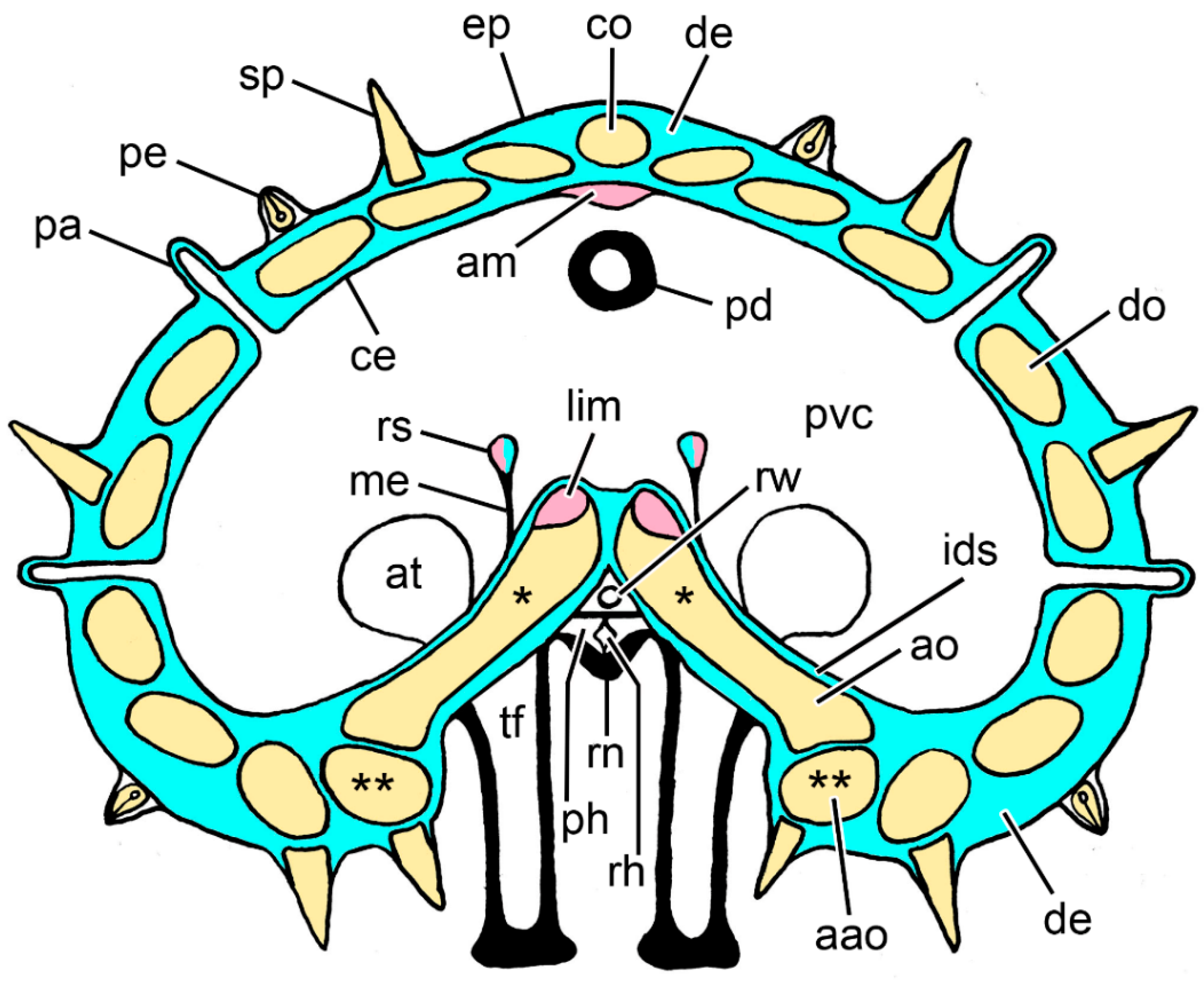
2.1. Anatomical Background
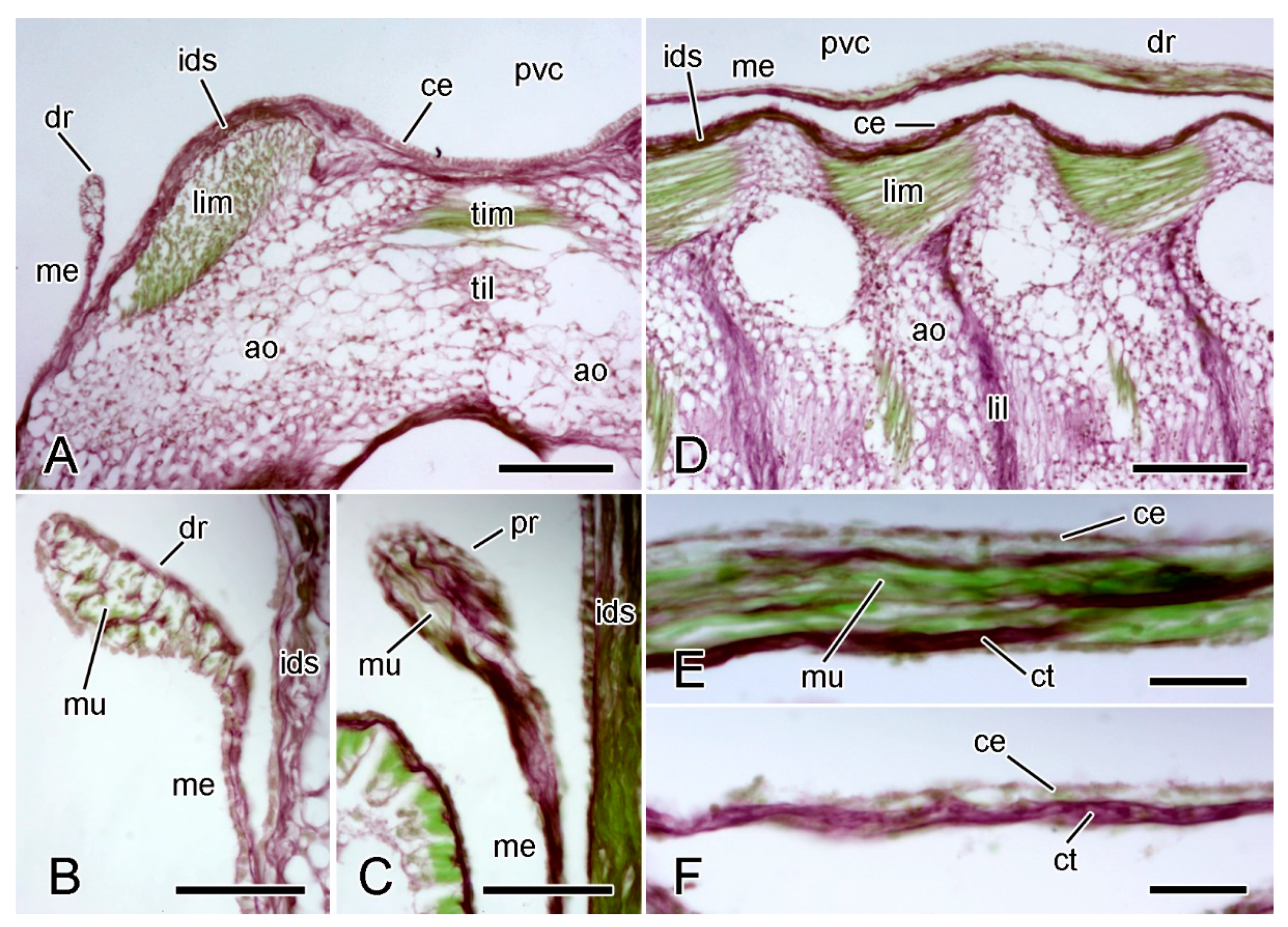
2.2. Dorsolateral Body Wall
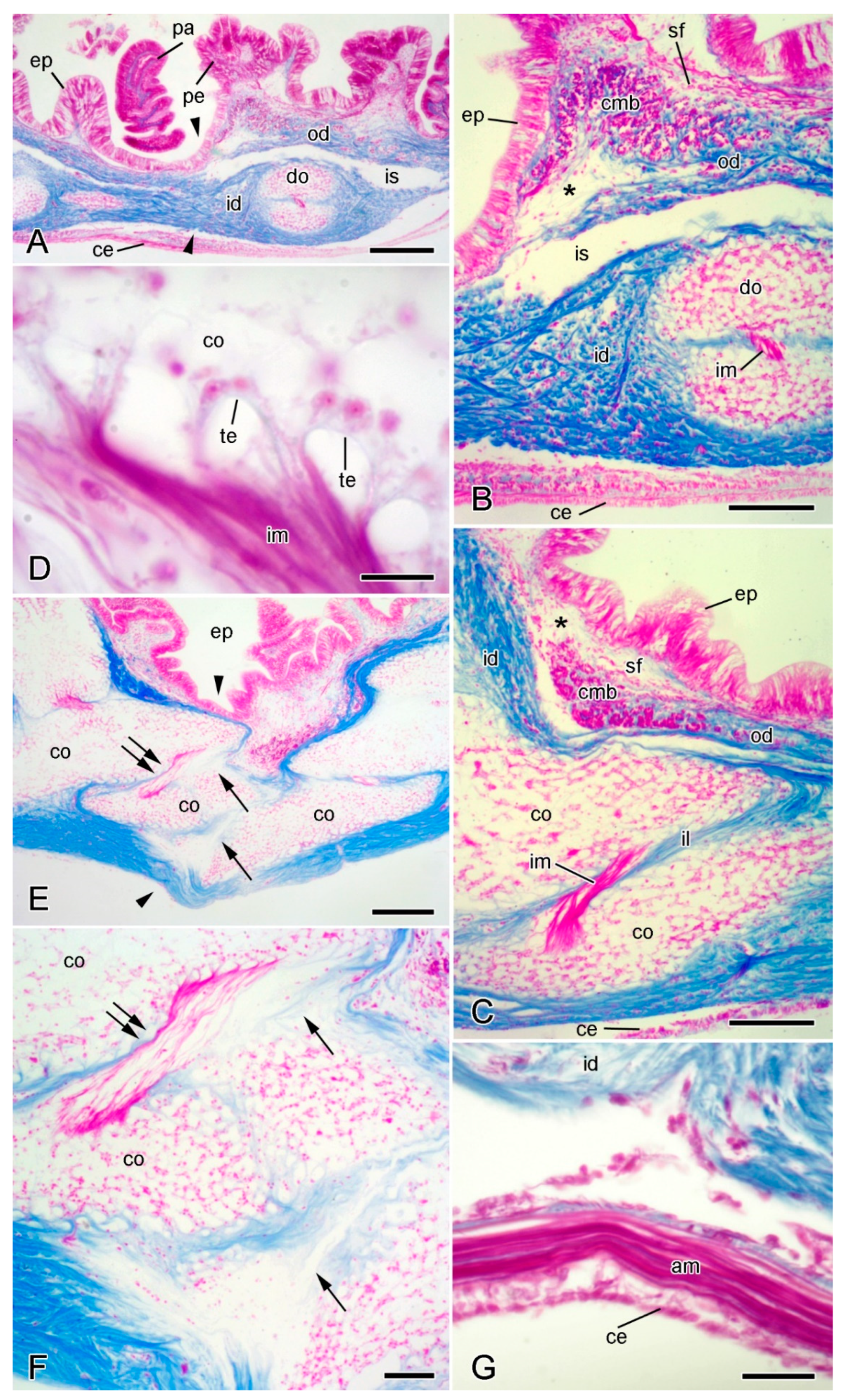
2.2.1. Organisation of Intact Dorsolateral Body Wall

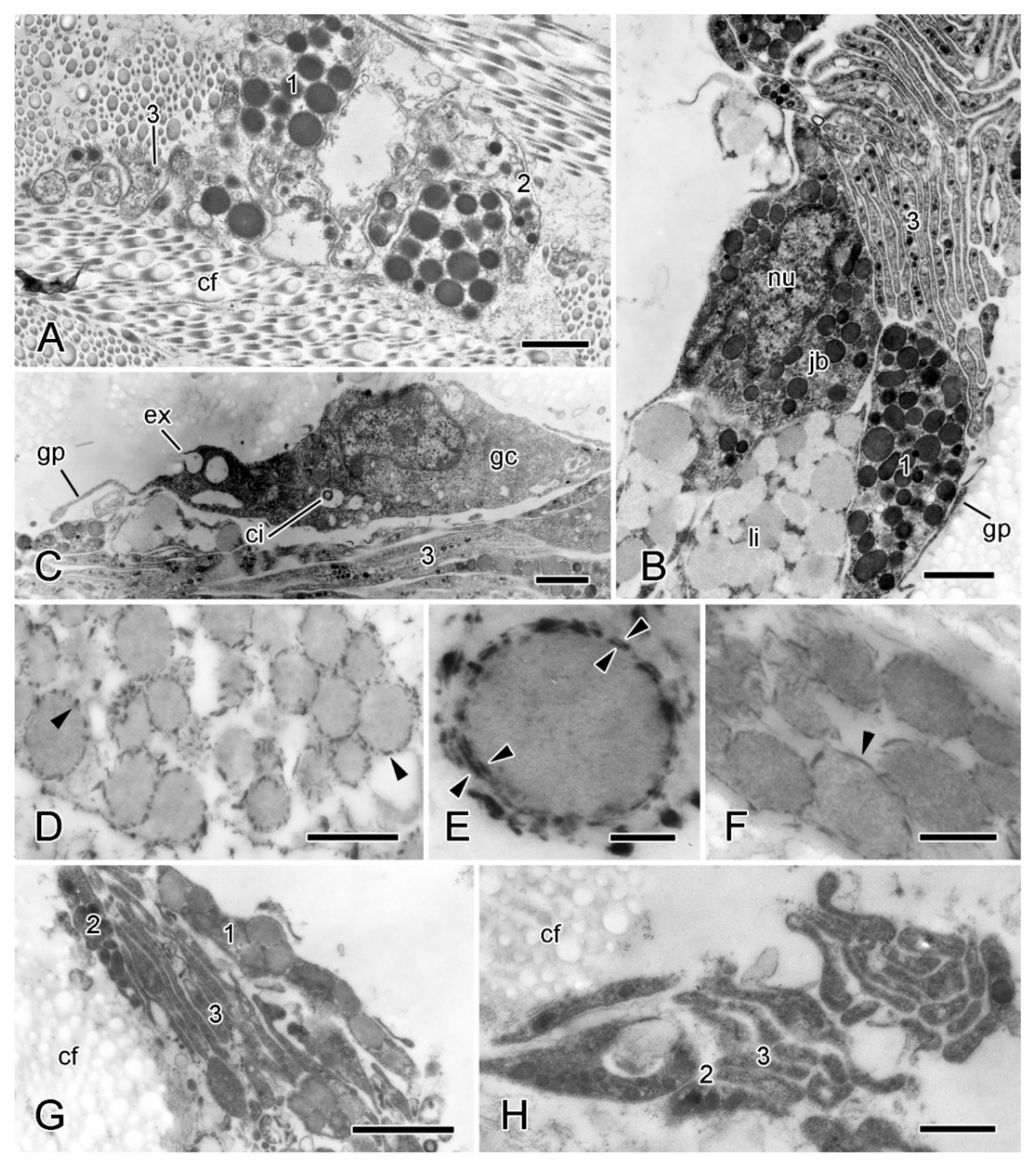
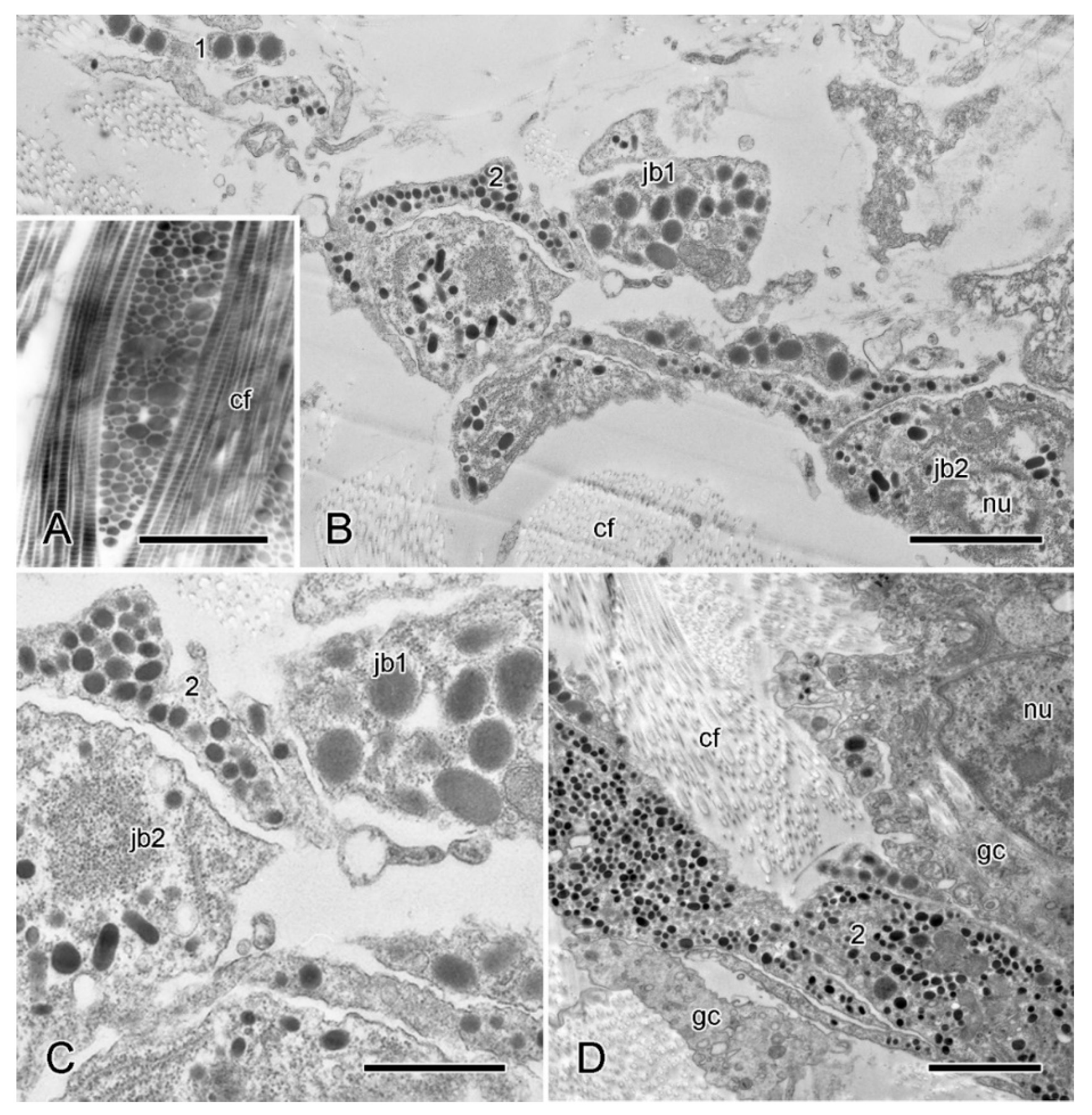
2.2.2. Organisation of Autotomising Dorsolateral Body Wall
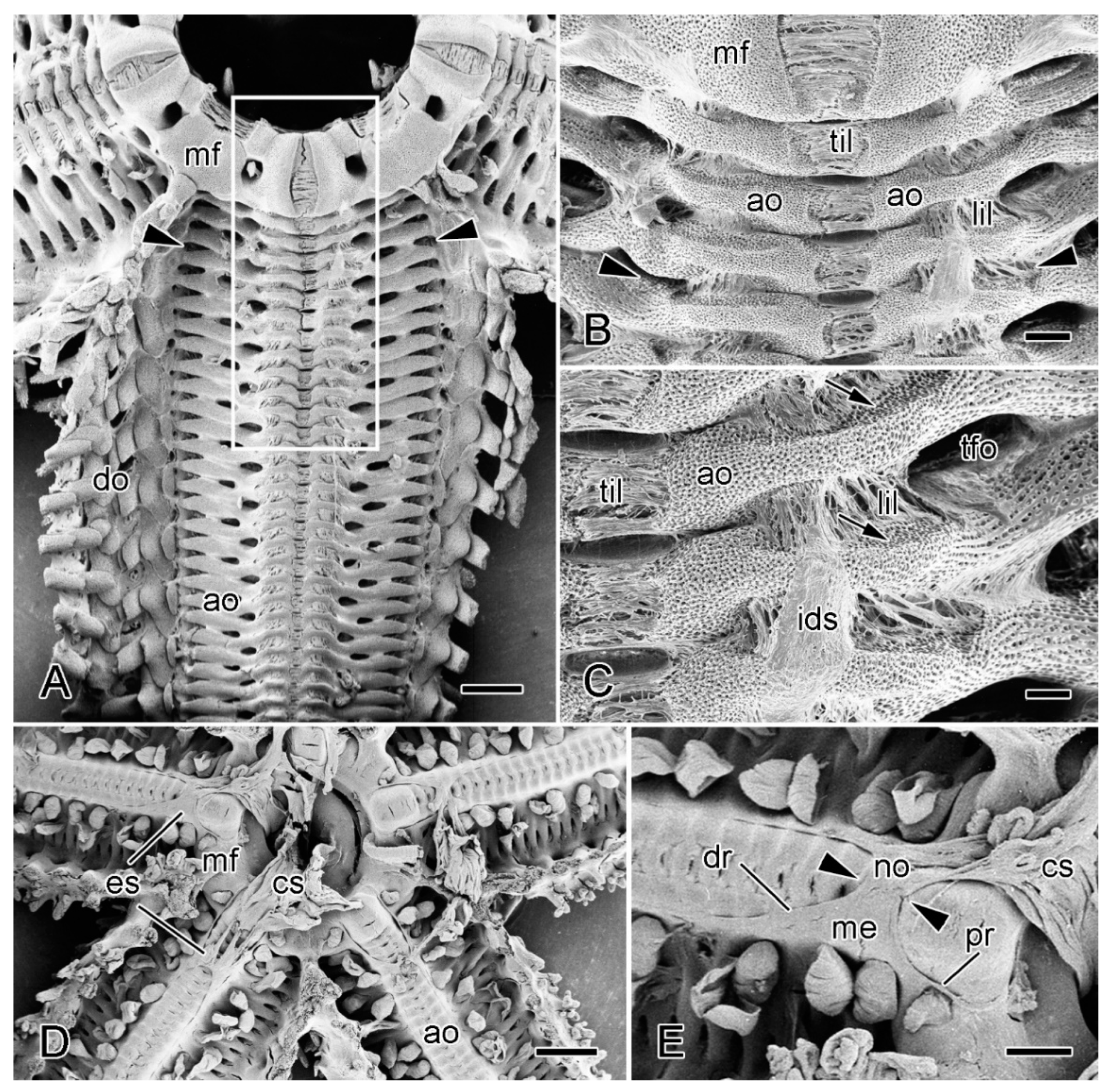
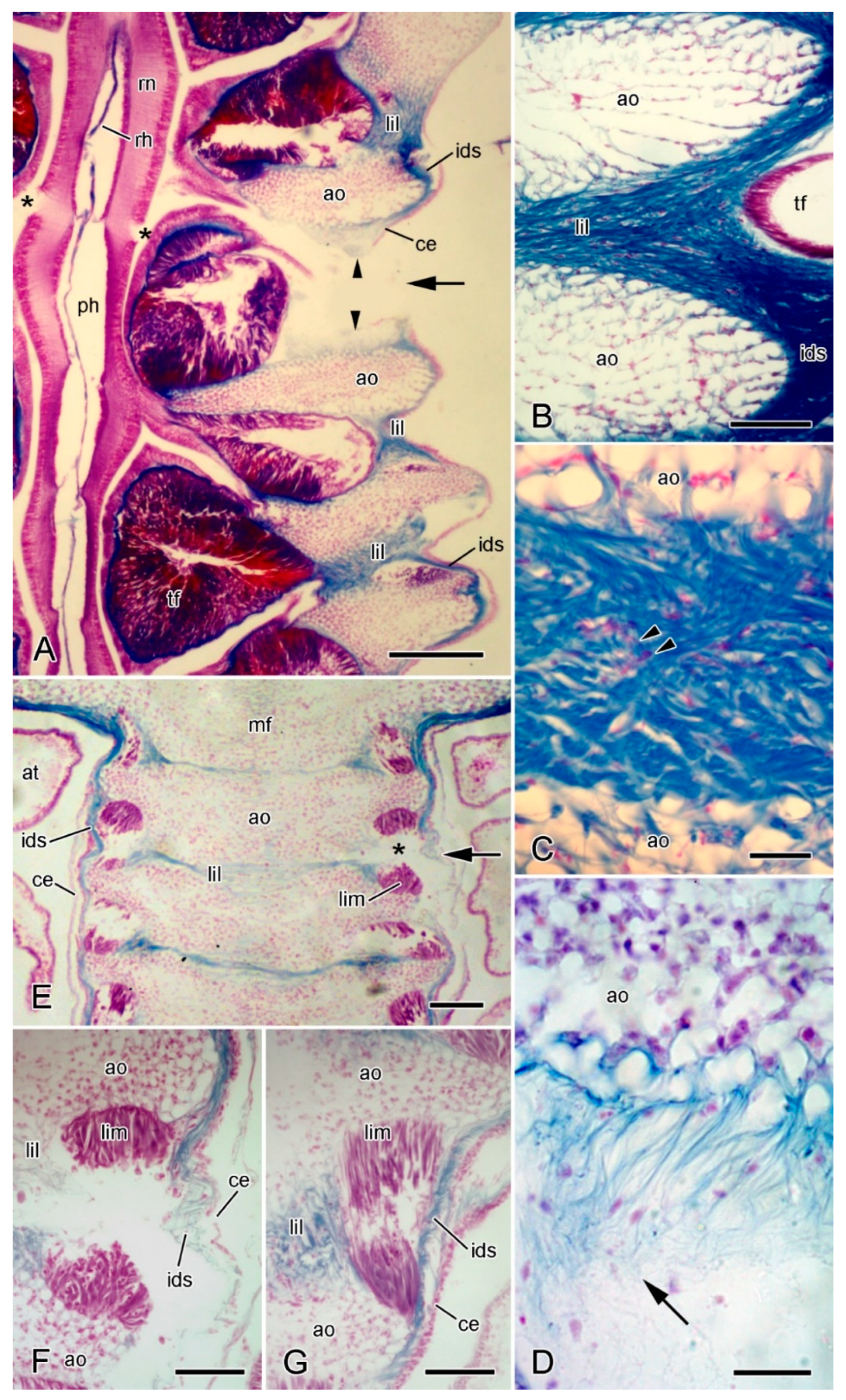
2.3. Ambulacral Body Wall
2.3.1. Organisation of Intact Ambulacral Body Wall
2.3.2. Organisation of Autotomising Ambulacral Body Wall
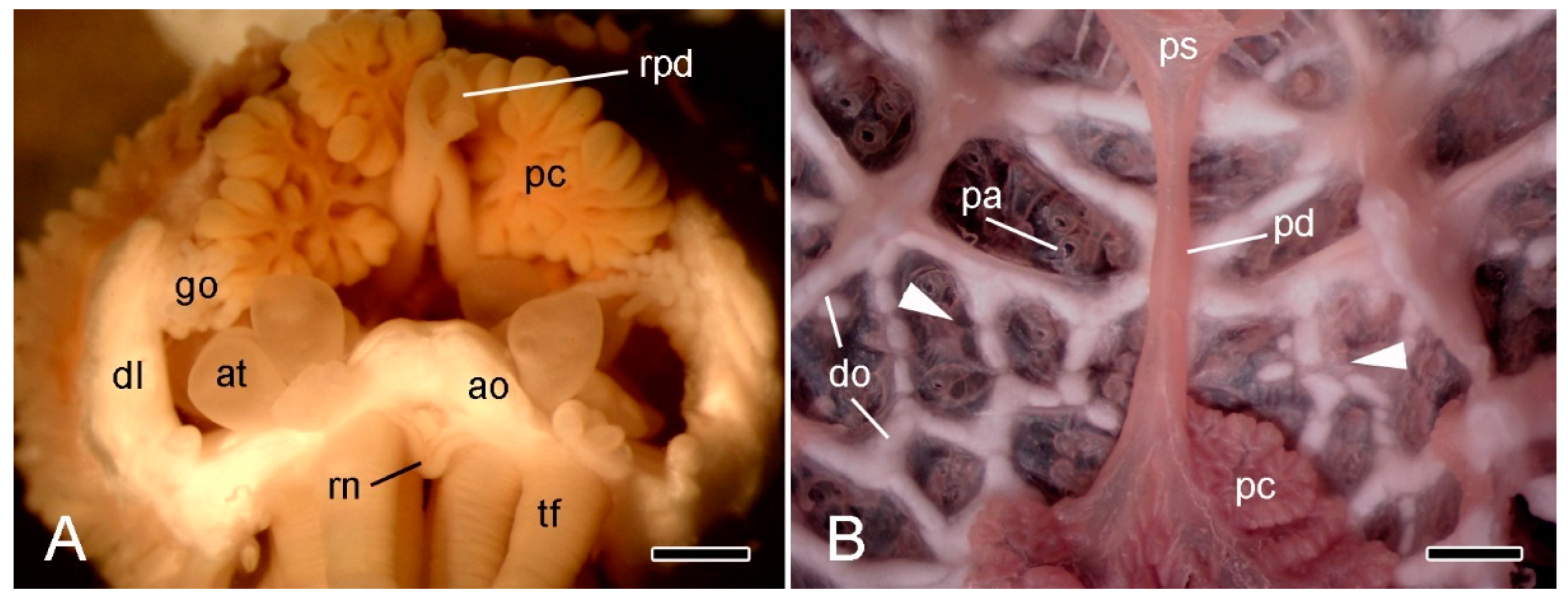
2.4. Extrinsic Stomach Retractor Apparatus
2.5. Comments on Morphological Aspects
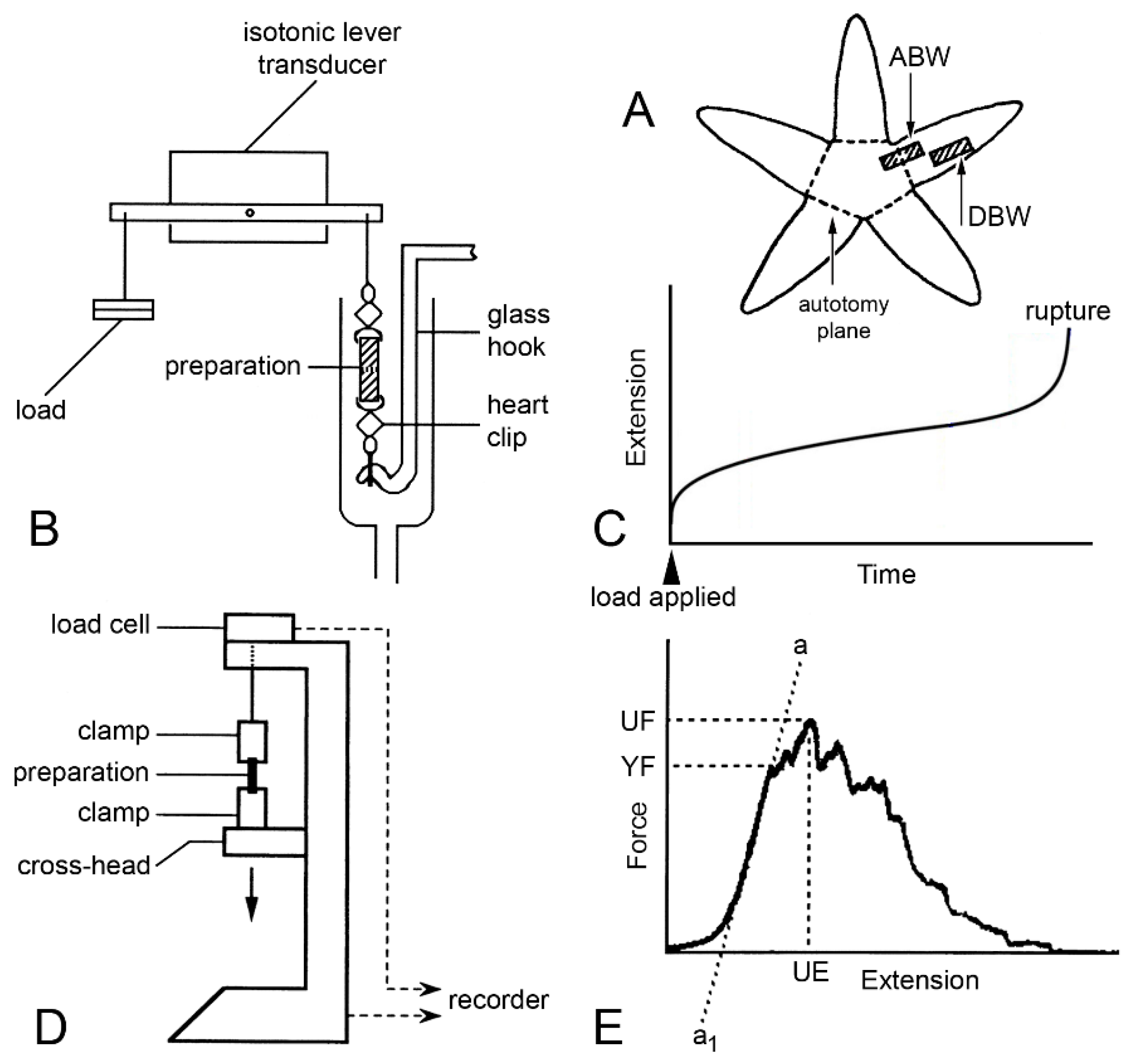
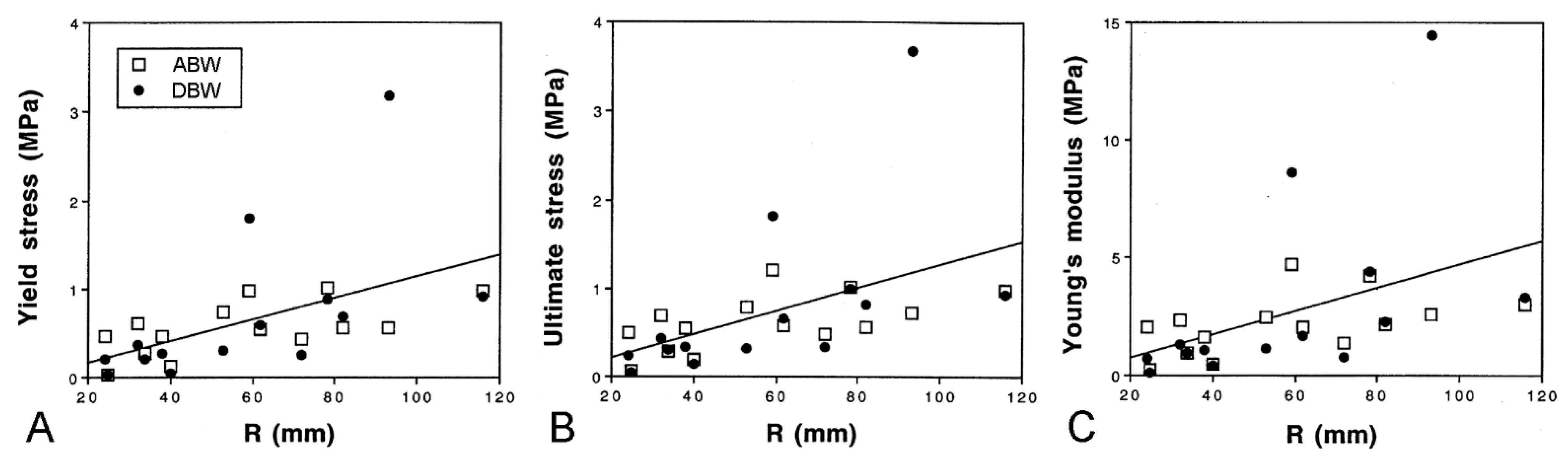
2.5.1. Morphological Insights into Breakage Mechanisms
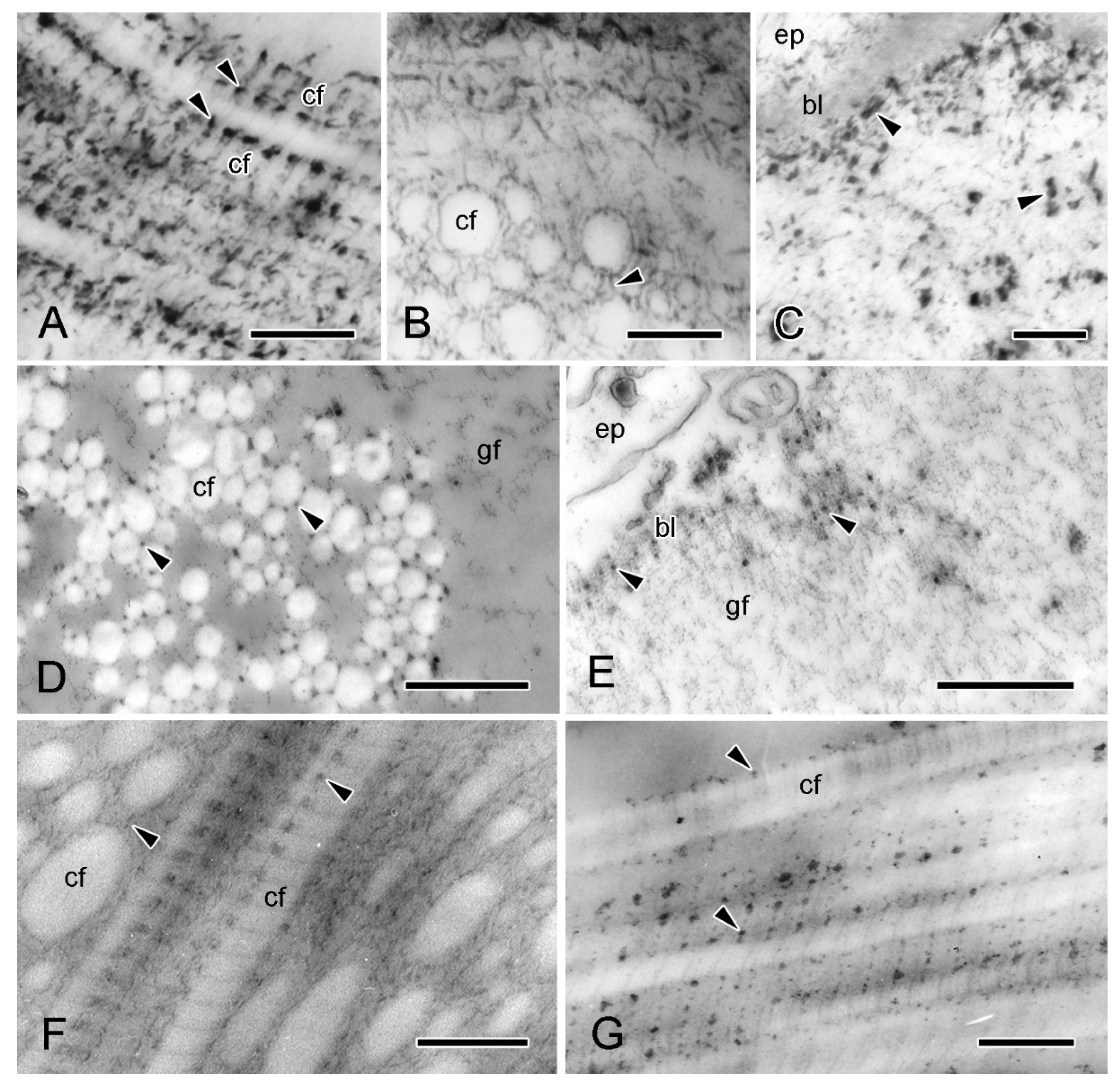
2.5.2. Juxtaligamental Components and Their Significance
3. Physiological Aspects
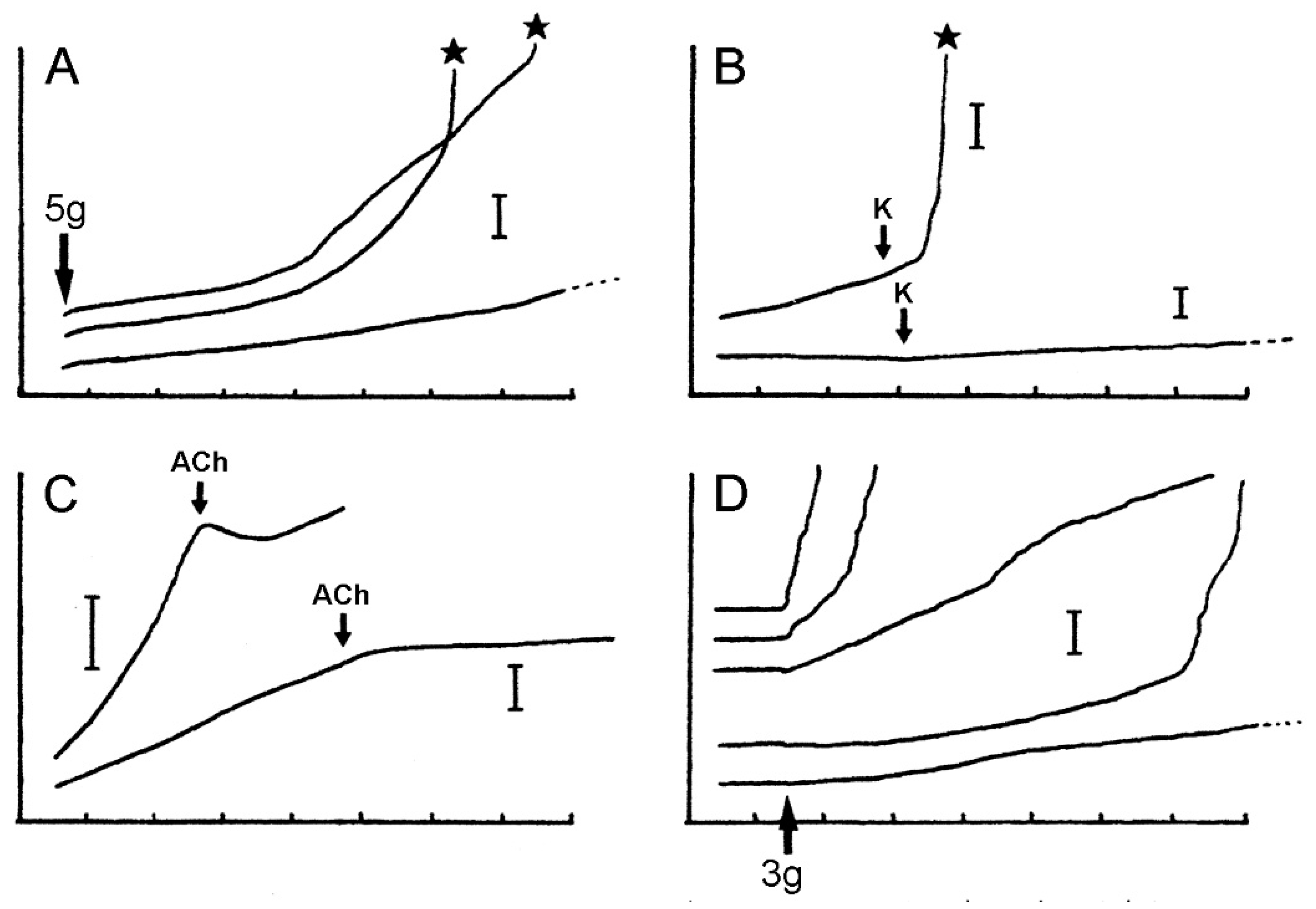
3.1. Dorsolateral Body Wall
3.2. Ambulacral Body Wall
3.3. Extrinsic Stomach Retractor Apparatus
3.4. Comments on Physiological Aspects
4. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations Used in Main Text
| ABW | isolated preparation of proximal dorsolateral body wall including autotomy plane |
| AGE | advanced glycation end-product |
| CMB | cupromeronic blue |
| DBW | isolated preparation of distal dorsolateral body wall lacking autotomy plane |
| ESFR | expanded sparsely fibrous region |
| ESRA | extrinsic stomach retractor apparatus |
| JLC | juxtaligamental cell |
| LDCV | large dense-core vesicle |
| LM | light microscopy |
| MCT | mutable collagenous tissue |
| MPa | megapascal |
| PEI | polyethyleneimine |
| PG | proteoglycan |
| SEM | scanning electron microscopy |
| STM | stereomicroscopy |
| TEM | transmission electron microscopy |
References
- Wilkie, I.C.; Sugni, M.; Gupta, H.S.; Candia Carnevali, M.D.; Elphick, M.R. The mutable collagenous tissue of echinoderms: From biology to biomedical applications. In Soft Matter for Biomedical Applications; Azevedo, H.S., Mano, J.F., Borges, J., Eds.; Royal Society of Chemistry: Cambridge, UK, 2021; pp. 3–33. [Google Scholar]
- Wilkie, I.C. Is muscle involved in the mechanical adaptability of echinoderm mutable collagenous tissue? J. Exp. Biol. 2002, 205, 159–165. [Google Scholar] [CrossRef]
- Takemae, N.; Motokawa, T. Mechanical properties of the isolated catch apparatus of the sea urchin spine joint: Muscle fibers do not contribute to passive stiffness changes. Biol. Bull. 2005, 208, 29–35. [Google Scholar] [CrossRef]
- Motokawa, T.; Sato, E.; Umeyama, K. Energy expenditure associated with softening and stiffening of echinoderm connective tissue. Biol. Bull. 2012, 222, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, I.C.; Emson, R.H.; Mladenov, P.V. Morphological and mechanical aspects of fission in Ophiocomella ophiactoides. Zoomorphology 1984, 104, 310–322. [Google Scholar] [CrossRef]
- Wilkie, I.C. Autotomy as a prelude to regeneration in echinoderms. Microscop. Res. Tech. 2001, 55, 369–396. [Google Scholar] [CrossRef]
- Rubilar, T.; Pastor, C.; Díaz de Vivar, E. Timing of fission in the starfish Allostichaster capensis (Echinodermata: Asteroidea) in laboratory. Rev. Biol. Trop. 2005, 53 (Suppl. 3), 299–303. [Google Scholar] [PubMed]
- Dolmatov, I.Y.; Afanasyev, S.V.; Boyko, A.V. Molecular mechanisms of fission in echinoderms: Transcriptome analysis. PLoS ONE 2018, 13, e0195836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, E. A History of British Starfishes, and Other Animals of the Class Echinodermata; John Van Voorst: London, UK, 1841. [Google Scholar]
- King, H.D. Regeneration in Asterias vulgaris. Roux Arch. Dev. Biol. 1898, 7, 351–363. [Google Scholar] [CrossRef]
- Chadwick, H.C. L.M.B.C. Memoirs. XXV. Asterias; Hodder & Stoughton Ltd.: London, UK, 1923. [Google Scholar]
- Cuénot, L. Anatomie, éthologie et systématiques des échinodermes. In Traité de Zoologie; Grassé, P., Ed.; Masson: Paris, France, 1948; Volume 11. [Google Scholar]
- Lawrence, J.M.; Vasquez, J. The effect of sublethal predation on the biology of echinoderms. Oceanol. Acta 1996, 19, 431–440. [Google Scholar]
- Ramsay, K.; Kaiser, M.J.; Richardson, C.A. Invest in arms: Behavioural and energetic implications of multiple autotomy in starfish (Asterias rubens). Behav. Ecol. Sociobiol. 2001, 50, 360–365. [Google Scholar]
- Marrs, J.; Wilkie, I.C.; Sköld, M.; Maclaren, W.M.; McKenzie, J.D. Size-related aspects of arm damage, tissue mechanics, and autotomy in the starfish Asterias rubens. Mar. Biol. 2000, 137, 59–70. [Google Scholar] [CrossRef]
- Clark, A.M.; Downey, M.E. Starfishes of the Atlantic; Chapman & Hall: London, UK, 1992. [Google Scholar]
- Wares, J.P. Biogeography of Asterias: North Atlantic climate change and speciation. Biol. Bull. 2001, 201, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Harper, F.M.; Hart, M.W. Morphological and phylogenetic evidence for hybridization and introgression in a sea star secondary contact zone. Invert. Biol. 2007, 126, 373–384. [Google Scholar] [CrossRef]
- Wilkie, I.C.; Griffiths, G.V.R.; Glennie, S.F. Morphological and physiological aspects of the autotomy plane in the aboral integument of Asterias rubens L. (Echinodermata). In Echinoderm Research; De Ridder, C., Dubois, P., Lahaye, M., Jangoux, M., Eds.; Balkema: Rotterdam, The Netherlands, 1990; pp. 301–313. [Google Scholar]
- Blowes, L.M.; Egertová, M.; Liu, Y.; Davis, G.R.; Terrill, N.J.; Gupta, H.S.; Elphick, M.R. Body wall structure in the starfish Asterias rubens. J. Anat. 2017, 231, 325–341. [Google Scholar] [CrossRef] [Green Version]
- Schwertmann, L.; Focke, O.; Dirks, J.H. Morphology, shape variation and movement of skeletal elements in starfish (Asterias rubens). J. Anat. 2019, 234, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.D. Echinodermata. Epidermal cells. In Biology of the Integument. 1. Invertebrates; Bereiter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin, Germany, 1984; pp. 756–774. [Google Scholar]
- Viehweg, J.; Naumann, W.W.; Olsson, R. Secretory radial glia in the ectoneural system of the sea star Asterias rubens (Echinodermata). Acta Zool. 1998, 79, 119–131. [Google Scholar] [CrossRef]
- Ferguson, J.C. Translocative functions of the enigmatic organs of starfish—The axial organ, hemal vessels, Tiedemann’s bodies, and rectal caeca: An autoradiographic study. Biol. Bull. 1984, 166, 140–155. [Google Scholar] [CrossRef]
- Emson, R.H.; Wilkie, I.C. Fission and autotomy in echinoderms. Oceanogr. Mar. Biol. Ann. Rev. 1980, 18, 155–250. [Google Scholar]
- Wilkie, I.C.; Candia Carnevali, M.D. The juxtaligamental cells of echinoderms and their role in the mechano-effector function of connective tissue. In Frontiers in Invertebrate Physiology: A Collection of Reviews, Volume 3: Annelida and Echinodermata; Saleuddin, A.S., Leys, S., Roer, R., Wilkie, I.C., Eds.; Apple Academic Press: Point Pleasant, NJ, USA, 2023; in press. [Google Scholar]
- McKenzie, J.D. Echinoderm surface coats: Their ultrastructure, function and significance. In Echinoderm Biology; Burke, R.D., Mladenov, P.V., Lambert, P., Parsley, R.L., Eds.; Balkema: Rotterdam, The Netherlands, 1988; pp. 697–706. [Google Scholar]
- Guatelli, S.; Ferrario, C.; Bonasoro, F.; Anjo, S.I.; Manadas, B.; Candia Carnevali, M.D.; Coelho, A.V.; Sugni, M. More than a simple epithelial layer: Multifunctional role of echinoderm coelomic epithelium. Cell Tissue Res. 2022, 390, 207–227. [Google Scholar] [CrossRef]
- Yourassowsky, C. Le Système Coelomique de l’Asterie Asterias rubens L. (Echinodermata—Asteroidea). Développement, Morphologie Compare et Signification Fonctionelle. Ph.D. Thesis, Université Libre de Bruxelles, Bruxelles, Belgium, 1985. Volume 2. [Google Scholar]
- Jangoux, M.; Dubois, P.; Lambert, A.; Yourassowsky, C. Occurrence of a microcanalicular system within the ossicles and dermal tissue of Asterias rubens and Marthasterias glacialis (Echinodermata: Asteroidea). Cell Tissue Res. 1985, 242, 657–660. [Google Scholar] [CrossRef]
- Humason, G.L. Animal Tissue Techniques, 4th ed.; Freeman: San Francisco, CA, USA, 1979. [Google Scholar]
- Ameye, R.H.; Killian, C.; Wilt, F.; Dubois, P. Ultrastructural localization of proteins involved in sea urchin biomineralization. J. Histochem. Cytochem. 1999, 47, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.E. Elasticity in extracellular matrix ‘shape modules’ of tendon, cartilage, etc. A sliding proteoglycan-filament model. J. Physiol. 2003, 553, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Linka, K.; Khiêm, V.N.; Itskov, M. Multi-scale modeling of soft fibrous tissues based on proteoglycan mechanics. J. Biomech. 2016, 49, 2349–2357. [Google Scholar] [CrossRef]
- Robinson, K.A.; Sun, M.; Barnum, C.E.; Weiss, S.N.; Huegel, J.; Shetye, S.S.; Lin, L.; Saez, D.; Adams, S.M.; Iozzo, R.V.; et al. Decorin and biglycan are necessary for maintaining collagen fibril structure, fiber realignment, and mechanical properties of mature tendons. Matrix Biol. 2017, 64, 81–93. [Google Scholar] [CrossRef]
- Sauren, Y.M.H.F.; Mieremet, R.H.P.; Groot, C.G.; Koerten, H.K.; Scherft, J.P. Polyethyleneimine as a contrast agent for ultrastructural localization and characterization of proteoglycans in the matrix of cartilage and bone. J. Histochem. Cytochem. 1991, 39, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Erlinger, R.; Welsch, U.; Scott, J.E. Ultrastructural and biochemical observations on proteoglycans and collagen in the mutable collagenous tissue of the feather star Antedon bifida (Echinodermata, Crinoidea). J. Anat. 1993, 183, 1–11. [Google Scholar]
- Wilkie, I.C. Design for disaster: The ophiuroid intervertebral ligament as a typical mutable collagenous structure. In Echinoderm Biology; Burke, R.D., Mladenov, P.V., Lambert, P., Parsley, R.L., Eds.; Balkema: Rotterdam, The Netherlands, 1988; pp. 25–38. [Google Scholar]
- Trotter, J.A.; Koob, T.J. Collagen and proteoglycan in a sea urchin ligament with mutable mechanical properties. Cell Tissue Res. 1989, 258, 527–539. [Google Scholar] [CrossRef]
- Scott, J.E. Proteoglycan: Collagen interactions un connective tissues. Ultrastructural, biochemical, functional and evolutionary aspects. Int. J. Biol. Macromol. 1991, 13, 157–161. [Google Scholar] [CrossRef]
- Fujimoto, T.; Parton, R.G. Not just fat: The structure and function of the lipid droplet. Cold Spring Harb. Perspect. Biol. 2011, 3, a004838. [Google Scholar] [CrossRef] [Green Version]
- Von Hehn, G. Über den Feinbau des hyponeuralen Nervensystems des Seesternes (Asterias rubens L.). Z. Zellforsch. 1970, 105, 137–154. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Butt, A. Glial Physiology and Pathophysiology; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Scranton, T.W.; Iwata, M.; Carlson, S.S. The SV2 protein of synaptic vesicles is a keratan sulfate proteoglycan. J. Neurochem. 1993, 61, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Stout, K.A.; Dunn, A.R.; Hoffman, C.; Miller, G.W. The synaptic vesicle glycoprotein 2: Structure, function, and disease relevance. ACS Chem. Neurosci. 2019, 10, 3927–3938. [Google Scholar] [CrossRef] [PubMed]
- Welsch, U.; Lange, A.; Bals, R.; Heinzeller, T. Juxtaligamental cells in feather stars. In Echinoderm Research 1995; Emson, R.H., Smith, A.B., Campbell, A.C., Eds.; Balkema: Rotterdam, The Netherlands, 1995; pp. 129–135. [Google Scholar]
- Kolset, S.O.; Prydz, K.; Pejler, G. Intracellular proteoglycans. Biochem. J. 2004, 379, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.; Mazzone, F.; Elphick, M.R.; Thorndyke, M.C.; Cisternas, P. Expression of the neuropeptide SALMFamide-1 during regeneration of the seastar radial nerve cord following arm autotomy. Proc. R. Soc. B 2019, 286, 20182701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlmann, K. Über die Verbindung der Muskulatur mit dem Skelett bei dem Echinodermen Asterias rubens L. Z. Zellforsch. 1968, 87, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Stauber, M.; Märkel, K. Comparative morphology of muscle-skeleton attachments in the Echinodermata. Zoomorphology 1988, 108, 137–148. [Google Scholar] [CrossRef]
- Hotchkiss, F.H.C.; Churchill, S.E.; Gelormini, R.G.; Hepp, W.R.; Rentler, R.J.; Tummarello, M.T. Events of autotomy in the starfish Asterias forbesi and A. vulgaris. In Biology of Echinodermata; Yanagisawa, T., Yasumasu, I., Oguro, C., Suzuki, N., Motokawa, T., Eds.; Balkema: Rotterdam, The Netherlands, 1991; pp. 537–541. [Google Scholar]
- Tartarin, A. Observations préliminaires sur l’autotomie chez Coscinasterias tenuispina Lamarck. Bull. Inst. Océanogr. Monaco 1949, 948, 1–4. [Google Scholar]
- Tartarin, A. Observations sur le mutilations, la regeneration, les néoformations, et l’anatomie de Coscinasterias tenuispina Lamarck. Recl. Trav. Stn. Mar. Endoume 1953, 5, 1–107. [Google Scholar]
- Anderson, J.M. Observations on autotomy in the starfish, Asterias forbesi. Biol. Bull. 1956, 111, 297. [Google Scholar]
- Wilkie, I.C.; Emson, R.H.; Mladenov, P.V. Autotomy mechanism and its control in the starfish Pycnopodia helianthoides. In Echinoderm Research 1995; Emson, R.H., Smith, A.B., Campbell, A.C., Eds.; Balkema: Rotterdam, The Netherlands, 1995; pp. 137–146. [Google Scholar]
- Anderson, J.M. Studies on the cardiac stomach of the starfish, Asterias forbesi. Biol. Bull. 1954, 107, 158–173. [Google Scholar] [CrossRef]
- Ferguson, J.C. The nature of the connective tissue of the body wall, retractor harness and cardiac stomach of the starfish, Asterias forbesi. Anat. Rec. 1960, 138, 348. [Google Scholar]
- Jangoux, M. Étude structurelle et fonctionnelle du tube digestif d’Asterias rubens L. (Echinodermata: Asteroidea). Austral. Mus. Mem. 1982, 16, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Wilson, P.O.G.; Chalk, B.T. The neuroendocrine system. In Theory and Practice of Histological Techniques, 3rd ed.; Bancroft, J.D., Stevens, A., Eds.; Churchill Livingstone: Edinburgh, UK, 1990; pp. 269–288. [Google Scholar]
- Wilkie, I.C.; Emson, R.H. The tendons of Ophiocomina nigra and their role in autotomy (Echinodermata, Ophiuroida). Zoomorphology 1987, 107, 33–44. [Google Scholar] [CrossRef]
- Wilkie, I.C. Functional morphology of the autotomy plane in the brittlestar Ophiocomina nigra (Abildgaard) (Ophiuroidea, Echinodermata). Zoomorphologie 1978, 91, 289–305. [Google Scholar] [CrossRef]
- Twarog, B.M. Aspects of smooth muscle function in molluscan catch muscle. Physiol. Rev. 1976, 56, 829–838. [Google Scholar] [CrossRef]
- Vevers, H.G. The biology of Asterias rubens L.: Growth and reproduction. J. Mar. Biol. Ass. UK 1949, 28, 165–187. [Google Scholar] [CrossRef] [Green Version]
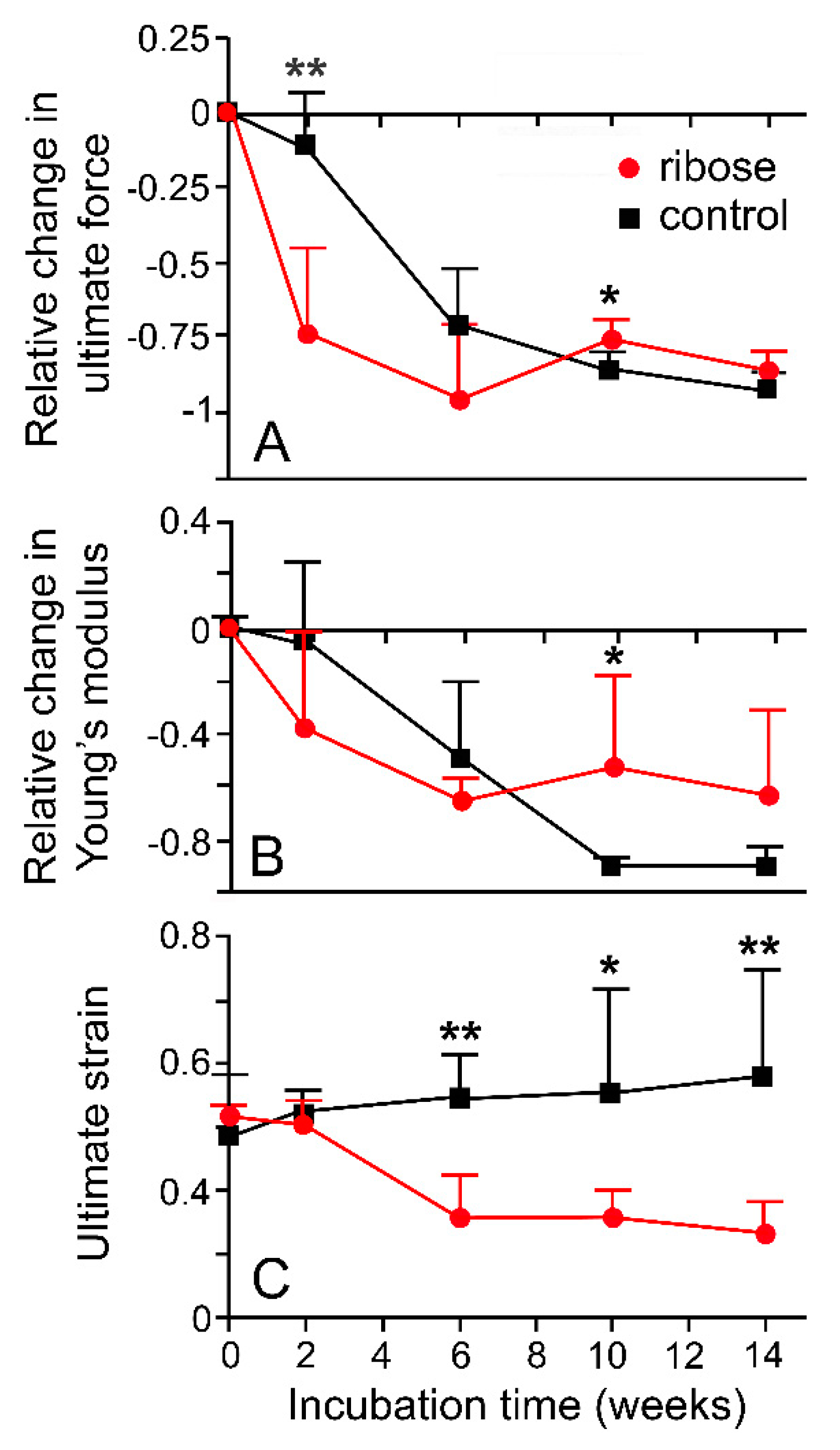
- Hayashi, K.; Hirayama, E. Age-related changes of wall composition and collagen cross-linking in the rat carotid artery—In relation with arterial mechanics. J. Mech. Behav. Biomed. Mat. 2017, 65, 881–889. [Google Scholar] [CrossRef]
- Goh, K.L.; Holmes, D.F.; Lu, Y.H.; Kadler, K.E.; Purslow, P.P. Age-related dataset on the mechanical properties and collagen. fibril structure of tendons from a murine model. Sci. Data 2018, 5, 180140. [Google Scholar] [CrossRef] [Green Version]
- Joodaki, H.; Panzer, M.B. Skin mechanical properties and modeling: A review. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2018, 232, 323–343. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Passini, F.S.; Silván, U.; Guizar-Sicairos, M.; Carimati, G.; Volpi, P.; Moretti, M.; Schoenhuber, H.; Redaelli, A.; Berli, M.; et al. Advanced glycation end-products: Mechanics of aged collagen from molecule to tissue. Matrix Biol. 2017, 59, 95–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, R.B.; Smith, S.T.; Moyer, P.J.; Magnusson, S.P. Effects of maturation and advanced glycation on tensile mechanics of collagen fibrils from rat tail and Achilles tendon. Acta Biomater. 2018, 70, 270–280. [Google Scholar] [CrossRef]
- Reddy, G.K.; Stehno-Bittel, L.; Enwekema, C.S. Glycation-induced matrix stability in the rabbit Achilles tendon. Arch. Biochem. Biophys. 2002, 399, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.K.; Fischer, T.H.; Wagner, M.; Dewenter, M.; Vettel, C.; Bork, N.I.; Maier, L.S.; Conti, M.; Wess, J.; El-Armouche, A.; et al. Atropine augments cardiac contractility by inhibiting cAMPspecific phosphodiesterase type 4. Sci. Rep. 2017, 7, 15222. [Google Scholar] [CrossRef] [Green Version]
- Wen, Q.; Fan, T.J.; Tian, C.L. Cytotoxicity of atropine to human corneal endothelial cells by inducing mitochondrion-dependent apoptosis. Exp. Biol. Med. 2016, 241, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Burnett, A.L.; Anderson, J.M. The contractile properties of the retractor mechanism of the cardiac stomach in Asterias forbesi. Anat. Rec. 1955, 122, 463–464. [Google Scholar]
- Basch, P.F. Observations on the retractor strands of the starfish stomach. Biol. Rev. City Coll. N. Y. 1956, 18, 14–17. [Google Scholar]
- Lin, M.; Egertová, M.; Zampronio, C.G.; Jones, A.M.; Elphick, M.R. Functional characterization of a second pedal peptide/orcokinin-type neuropeptide signaling system in the starfish Asterias rubens. J. Comp. Neurol. 2018, 526, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, A.B.; Barreiro-Iglesias, A.; Guerra, L.A.Y.; Delroisse, J.; Zhang, Y.; Gunner, E.F.; Zampronio, C.G.; Jones, A.M.; Egertová, M.; Elphick, M.R. Ancient role of sulfakinin/cholecystokinintype signalling in inhibitory regulation of feeding processes revealed in an echinoderm. eLife 2021, 10, e65667. [Google Scholar] [CrossRef] [PubMed]
- Motokawa, T.; Wainwright, S.A. Stiffness of starfish arm and involvement of catch connective tissue in the stiffness change. Comp. Biochem. Physiol. A 1991, 100, 393–397. [Google Scholar]
- Paramonov, A.S.; Shulepko, A.S.; Makhonin, M.A.; Bychkov, M.L.; Kulbatskii, D.S.; Chernikov, A.M.; Myshkin, M.Y.; Shabelnikov, S.V.; Shenkarev, Z.O.; Kirpichnikov, M.P.; et al. New three-finger protein from starfish Asterias rubens shares structure and pharmacology with human brain neuromodulator Lynx2. Mar. Drugs 2022, 20, 503. [Google Scholar] [CrossRef]
- Morales, M.; Sierra, C.; Vidal, A.; del Castillo, J.; Smith, D.S. Pharmacological sensitivity of the articular capsule of the primary spines of Eucidaris tribuloides. Comp. Biochem. Physiol. C 1993, 105, 25–30. [Google Scholar] [CrossRef]
- Hunter, A.W.; Ortega-Hernández, J. A new somasteroid from the Fezouata Lagerstätte in Morocco and the Early Ordovician origin of Asterozoa. Biol. Lett. 2021, 17, 20200809. [Google Scholar] [CrossRef] [PubMed]
- Preyer, W. Über die Bewegungen der Seesterne. Mitt. Zool. Stat. Neapel 1886, 7, 27–127, 191–233. [Google Scholar]
- Lo Schiavo, A.; Ruocco, E.; Russo, T.; Brancaccio, G. Locus minoris resistentiae: An old but still valid way of thinking in medicine. Clin. Dermatol. 2014, 32, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, L.; Bellairs, A.A. The mechanism of autotomy in Lacerta. Br. J. Herpetol. 1972, 4, 276–286. [Google Scholar]
- McVean, A.R. Autotomy in Carcinus maenas (Decapoda: Crustacea). J. Zool. 1973, 169, 349–364. [Google Scholar] [CrossRef]
- Hodgson, A.N. Use of the intrinsic musculature for siphonal autotomy in the Solenacea (Mollusca: Bivalvia). Trans. R. Soc. S. Afr. 1984, 45, 129–137. [Google Scholar] [CrossRef]
- Wilkie, I.C. Functional morphology and biomechanics of cuticular fracture at the elytrophoral autotomy plane of the scaleworm Alentia gelatinosa (Annelida: Polynoidae). Invert. Biol. 2011, 130, 129–137. [Google Scholar] [CrossRef]
- Wilkie, I.C. Fracture promotion versus prevention: The cuticular breakage zone at the trunk autotomy plane of the scaleworm Harmothoe impar (Annelida: Polynoidae). Invert. Zool. 2020, 17, 76–92. [Google Scholar] [CrossRef]
- Baban, N.S.; Orozaliev, A.; Kirchhof, S.; Stubbs, C.J.; Song, Y.A. Biomimetic fracture model of lizard tail autotomy. Science 2022, 375, 770–774. [Google Scholar] [CrossRef]
- Bickell-Page, L.R. Autotomy of cerata by the nudibranch Melibe leonina (Mollusca): Ultrastructure of the autotomy plane and neural correlate of the behaviour. Phil. Trans. R. Soc. Lond. B 1989, 324, 149–172. [Google Scholar]
- Bickell-Page, L.R.; Mackie, G.O. Tentacle autotomy in the hydromedusa Aglantha digitale (Cnidaria): An ultrastructural and neurophysiological analysis. Phil. Trans. R. Soc. Lond. B 1991, 331, 155–170. [Google Scholar]
- Semmens, D.C.; Mirabeau, O.; Moghul, I.; Pancholi, M.R.; Wurm, Y.; Elphick, M.R. Transcriptomic identification of starfish neuropeptide precursors yields new insights into neuropeptide evolution. Open Biol. 2016, 6, 150224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento, P.; Little, D. Tendon and multiomics: Advantages, advances, and opportunities. npj Regen. Med. 2021, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Candia Carnevali, M.D.; Bonasoro, F. Microscopic overview of crinoid regeneration. Microsc. Res. Tech. 2001, 55, 403–426. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M. The link between autotomy and CNS regeneration: Echinoderms as non-model species for regenerative biology. BioEssays 2020, 42, 1900219. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
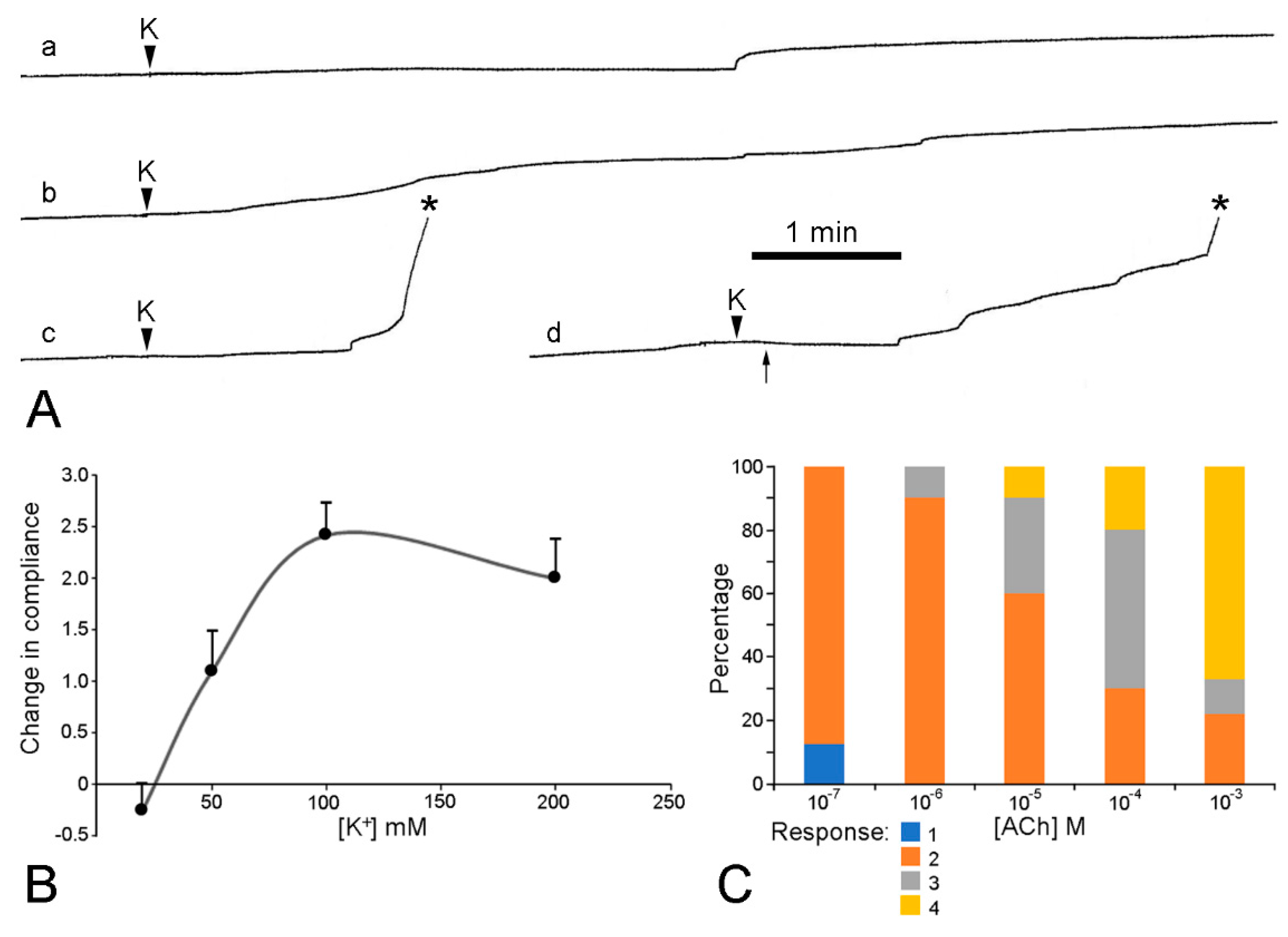
| Agent | Effect |
|---|---|
| Acetylcholine | decrease in extension rate (+++) |
| Muscarinic agonists | contraction |
| methacholine | decrease in extension rate (++) |
| carbachol | decrease in extension rate (+) |
| pilocarpine | 0 |
| arecoline | 0 |
| Nicotinic agonists | |
| acetylthiocholine | 0 |
| nicotine | 0 |
| DMPP | 0 |
| Muscarinic antagonists | |
| atropine | increase in extension rate |
| block of ACh effect | |
| no block of K+ effect | |
| isopropamide | no block of ACh or K+ effects |
| Nicotinic antagonists | |
| hexamethonium | no block of ACh or K+ effects |
| tetraethylammonium | no block of ACh or K+ effects |
| Amines | |
| adrenaline | 0 |
| noradrenaline | 0 |
| dopamine | 0 |
| 5-hydroxytryptamine | 0 |
| histamine | 0 |
| Agent | Effect |
|---|---|
| Acetylcholine | decrease in extension rate contraction |
| Muscarinic antagonists | |
| atropine | increase in extension rate |
| block of ACh effect | |
| no block of K+ effect | |
| isopropamide | no block of K+ effect |
| Nicotinic antagonists | |
| hexamethonium | no block of K+ effect |
| tetraethylammonium | no block of K+ effect |
| Amines | |
| tryptamine | 0 |
| 5-hydroxytryptamine | 0 |
| octopamine | 0 |
| GABA agonists | |
| GABA | 0 |
| imidazole | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilkie, I.C.; Candia Carnevali, M.D. Morphological and Physiological Aspects of Mutable Collagenous Tissue at the Autotomy Plane of the Starfish Asterias rubens L. (Echinodermata, Asteroidea): An Echinoderm Paradigm. Mar. Drugs 2023, 21, 138. https://doi.org/10.3390/md21030138
Wilkie IC, Candia Carnevali MD. Morphological and Physiological Aspects of Mutable Collagenous Tissue at the Autotomy Plane of the Starfish Asterias rubens L. (Echinodermata, Asteroidea): An Echinoderm Paradigm. Marine Drugs. 2023; 21(3):138. https://doi.org/10.3390/md21030138
Chicago/Turabian StyleWilkie, Iain C., and M. Daniela Candia Carnevali. 2023. "Morphological and Physiological Aspects of Mutable Collagenous Tissue at the Autotomy Plane of the Starfish Asterias rubens L. (Echinodermata, Asteroidea): An Echinoderm Paradigm" Marine Drugs 21, no. 3: 138. https://doi.org/10.3390/md21030138