

Effects of Different Nitrogen Concentrations on Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii
 ,
, 
Abstract
:1. Introduction
2. Results
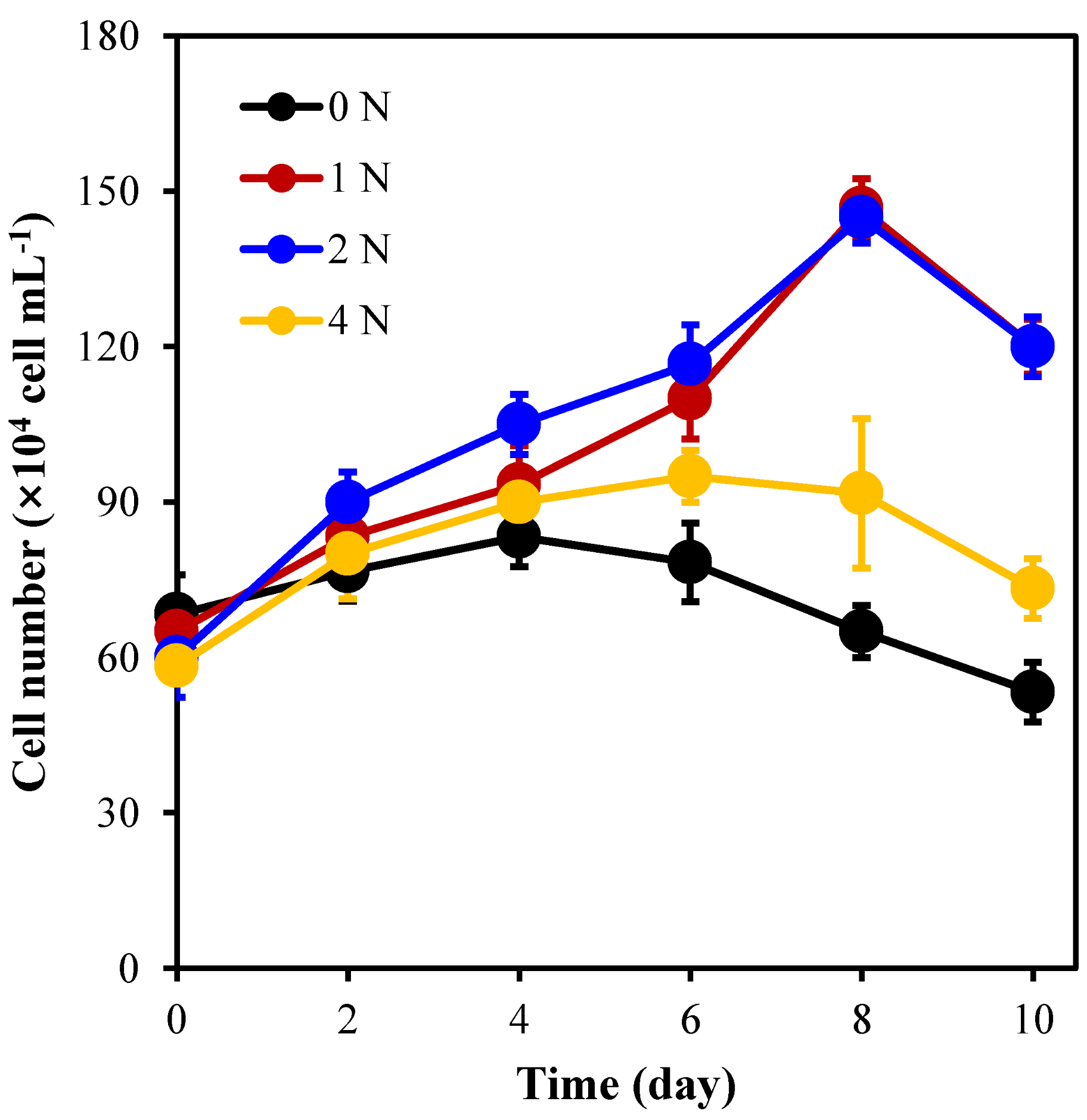
2.1. The Growth of C. weissflogii under Different Nitrogen Concentration Culture Conditions
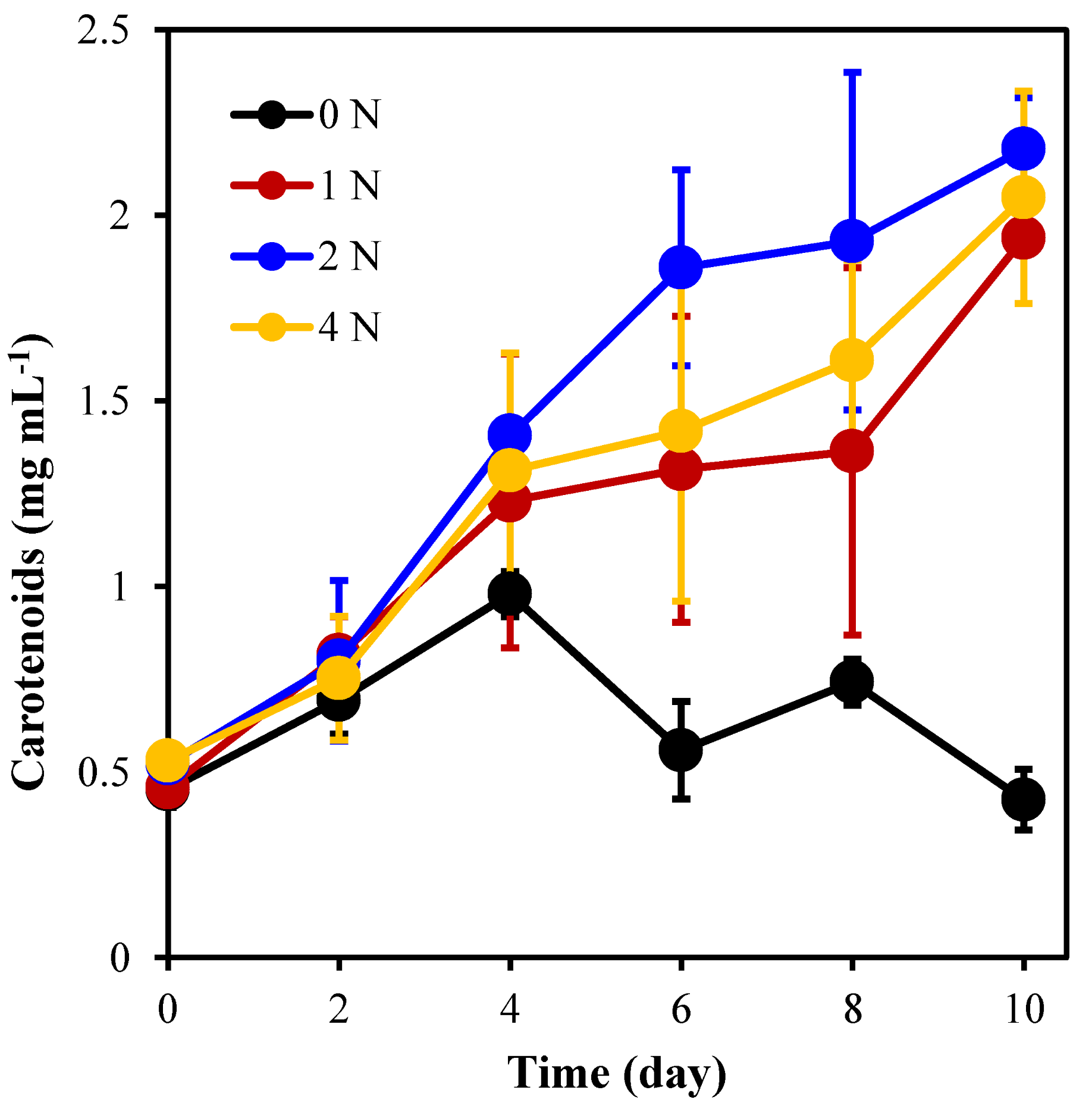
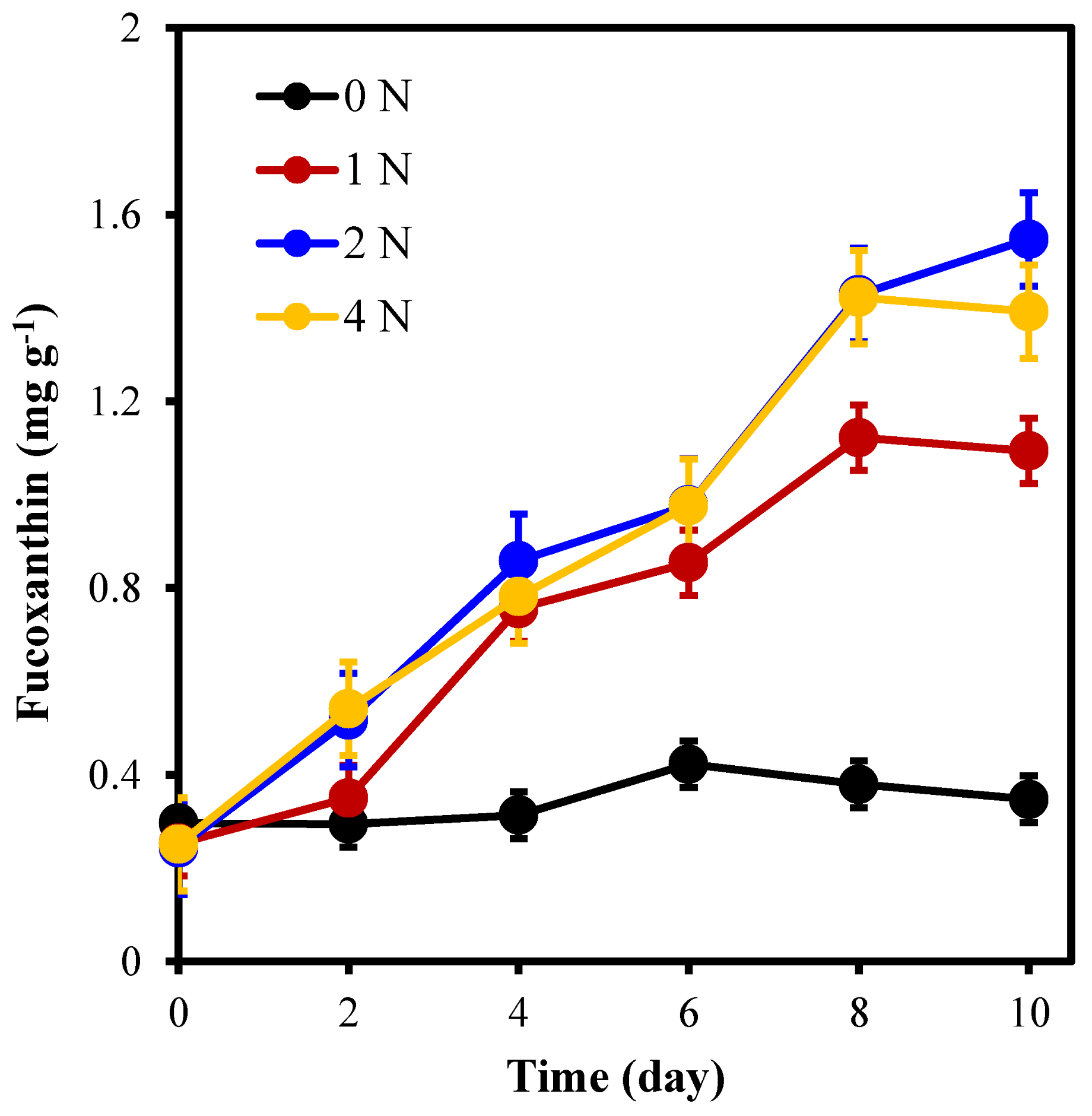
2.2. The Pigment Content of C. weissflogii under Different Nitrogen Concentration Culture Conditions
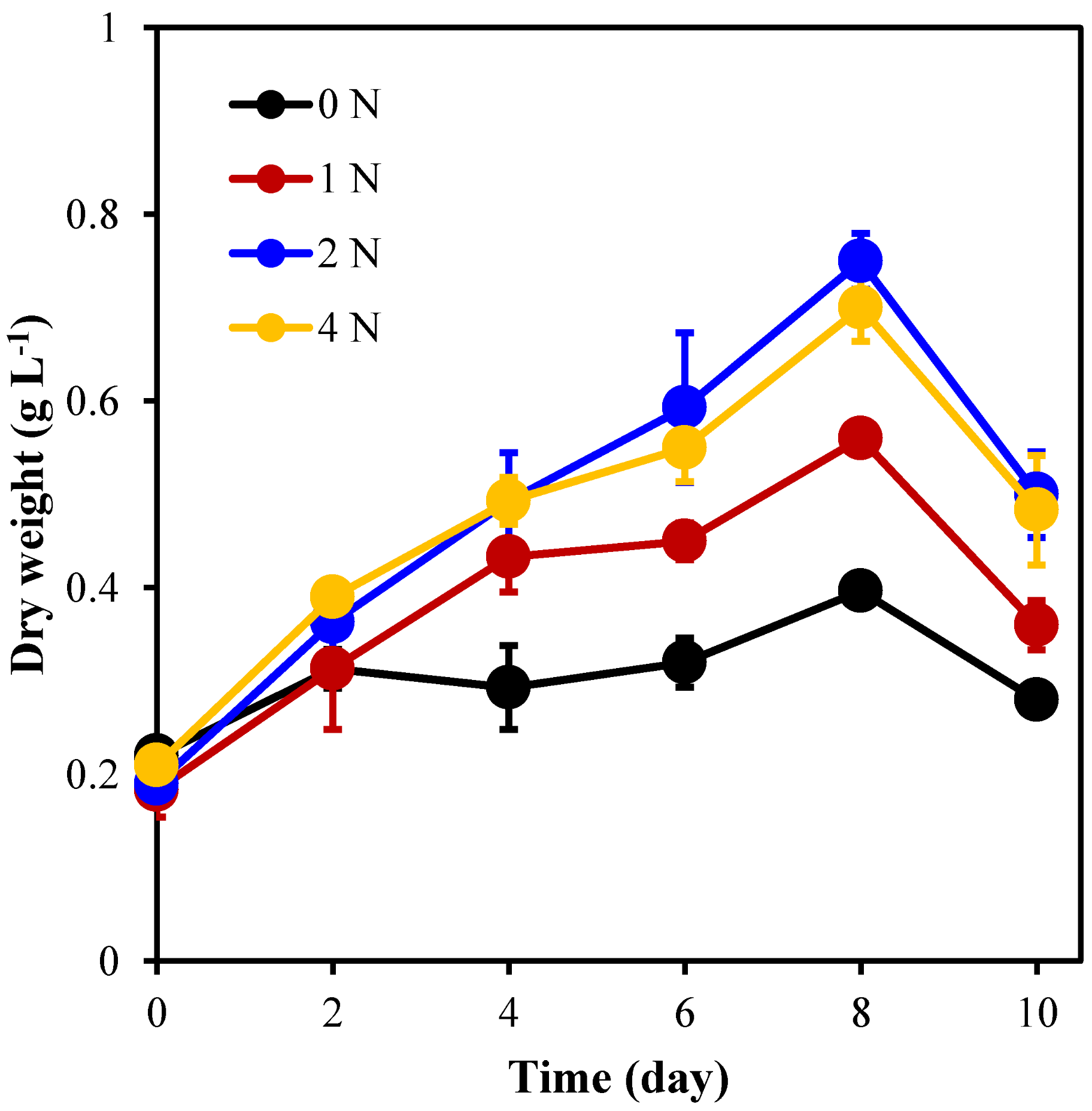
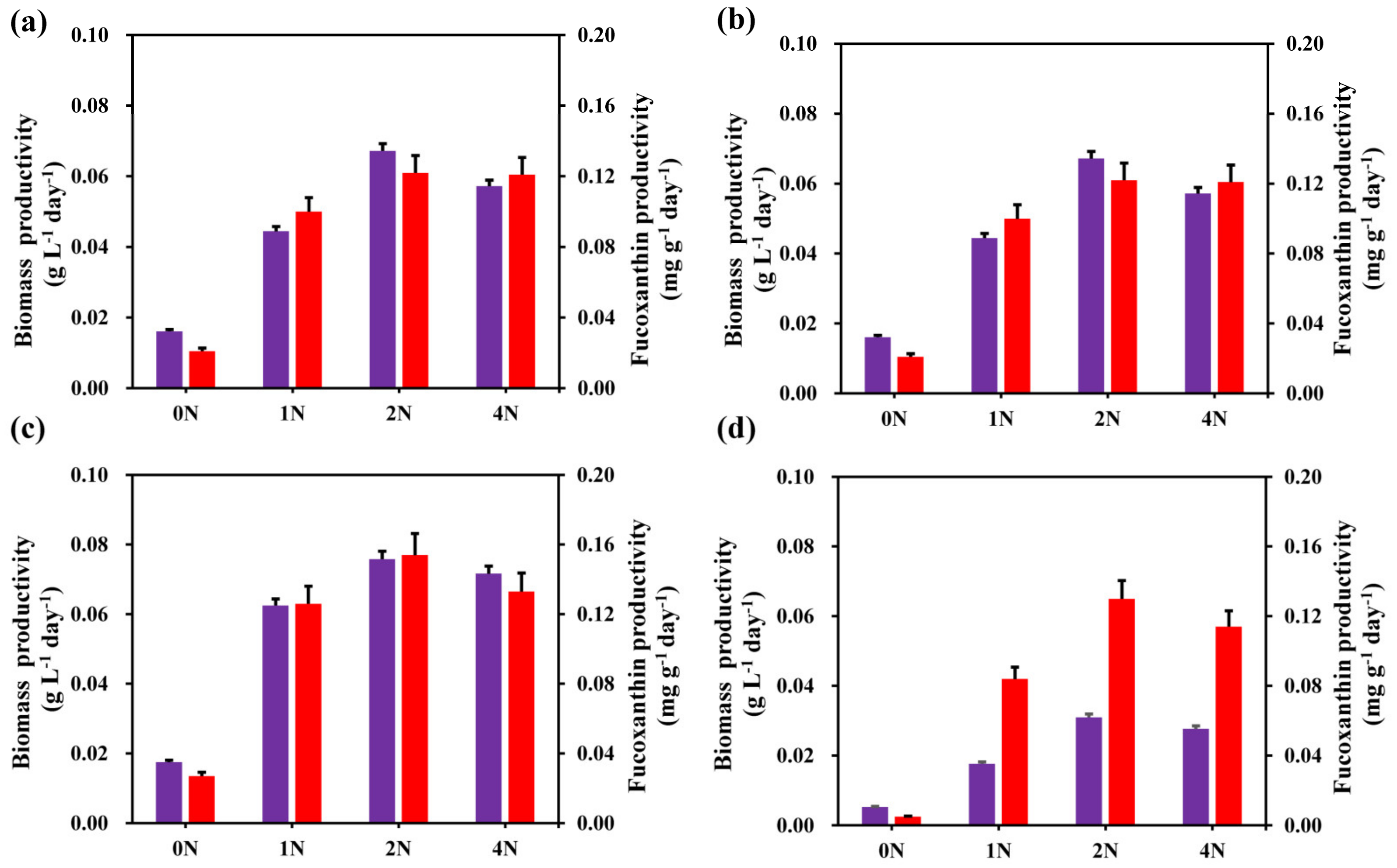
2.3. Effects of Different Nitrogen Concentrations on Biomass Productivity and Fucoxanthin Productivity of C. weissflogii
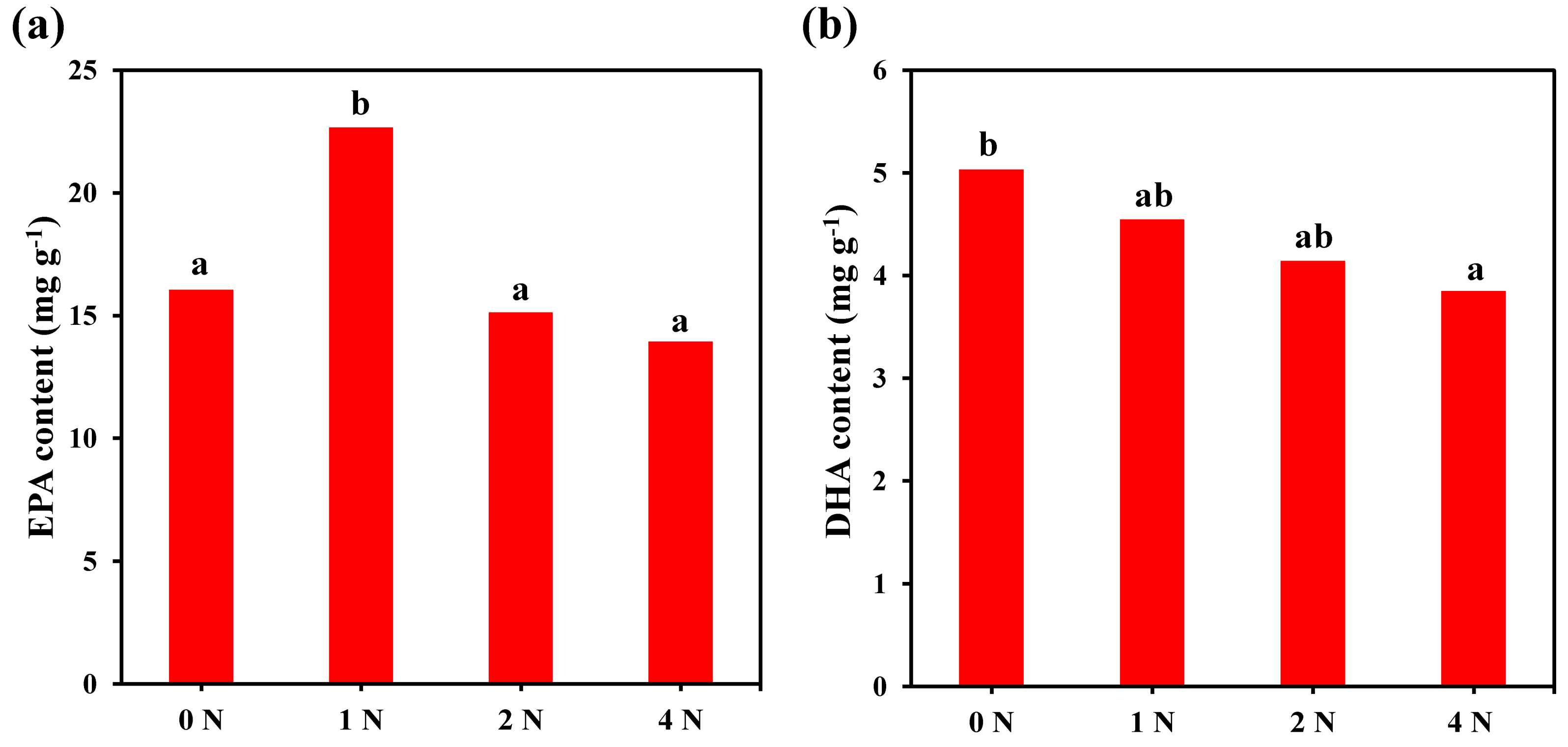
2.4. Fatty Acid Composition and Content of C. weissflogii under Different Nitrogen Concentrations
3. Discussion
4. Materials and Methods
4.1. Experimental Instruments and Materials
4.2. Algae Strain and Culture Conditions
4.3. Analytical Methods
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sarkarat, R.; Mohamadnia, S.; Tavakoli, O. Recent advances in non-conventional techniques for extraction of phycobiliproteins and carotenoids from microalgae. Braz. J. Chem. Eng. 2022. [Google Scholar] [CrossRef]
- Begum, H.; Yusoff, F.M.; Banerjee, S.; Khatoon, H.; Shariff, M. Availability and utilization of pigments from microalgae. Crit. Rev. Food Sci. Nutr. 2016, 56, 2209–2222. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.Q.; Hooi, R.L.; Kuan, S.K.; Hui, S.N.; Cai, Y.X.; Wang, J.H.; Andy, T.C.; Pau, L.S. An integration study of microalgae bioactive retention: From microalgae biomass to microalgae bioactive nanoparticle. Crit. Rev. Food Sci. Nutr. 2021, 158, 112607. [Google Scholar] [CrossRef] [PubMed]
- Yarnold, J.; Karan, H.; Oey, M.; Hankamer, B. Microalgal Aquafeeds As Part of a Circular Bioeconomy. Trends Plant Sci. 2019, 24, 959–970. [Google Scholar] [CrossRef]
- Zhang, G.W.; Li, Q.S.; Shao, D.H.; Qin, Y. Adances in the research and application of microalgae in aquaculture. China Fish. 2020, 2, 72–74. [Google Scholar]
- Zhang, W.Y.; Gao, B.Y.; Lei, X.Q.; Li, A.F.; Wu, J.; Zhang, C.W. Progress on physicochemical and biological properties, preparation techniques and physiological activities of fucoxanthin. Chin. J. Mar. Drugs 2015, 34, 81–95. [Google Scholar]
- Sugawara, T.; Matsubara, K.; Akagi, R.; Mori, M.; Hirata, T. Antiangiogenic activity of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. J. Agric. Food Chem. 2006, 54, 9805–9810. [Google Scholar] [CrossRef]
- Mikami, K.; Hosokawa, M. Biosynthetic pathway and health benefits of fucoxanthin, an algae-specific xanthophyll in brown seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef]
- Yoong, K.L.; Chun, Y.C.; Sunita, V.; Jo-Shu, C. Producing fucoxanthin from algae—Recent advances in cultivation strategies and downstream processing. Bioresour. Technol. 2021, 344, 126170. [Google Scholar]
- Song, W.; Sha, W.; Guan, P.Y.; Ke, H.P.; Lu, L.W.; Zhang, L.H. A review on the progress, challenges and prospects in commercializing microalgal fucoxanthin. Biotechnol. Adv. 2021, 53, 107865. [Google Scholar]
- Zhao, W.; Gao, B.Y.; Huang, L.D.; Wang, F.F.; Zhang, C.W. Nutritional value, physiological function and disease resistance activity of microalgae and their bioactive components in aquaculture. Feed. Ind. 2019, 40, 9–16. [Google Scholar]
- Lu, Q.; Li, H.K.; Xiao, Y.; Liu, H. A state-of-the-art review on the synthetic mechanisms, production technologies, and practical application of polyunsaturated fatty acids from microalgae. Algal Res. 2021, 55, 102281. [Google Scholar] [CrossRef]
- Daniel, G.B.; Vasile, C.; Dan, C.V. Microalgae as sources of omega-3 polyunsaturated fatty acids: Biotechnological aspects. Algal Res. 2021, 58, 102410. [Google Scholar]
- Hou, J.J.; Huang, B.Q.; Cao, Z.R.; Chen, J.X.; Hong, H.S. Effects of nutrient limitation on pigments in Thalassiosira weissflogii and Prorocentrum donghaiense. Chin. Bull. Bot. 2007, 49, 12. [Google Scholar] [CrossRef]
- Nuramina, M.; Ai, S.J.; Zhao, H.X. Advances in research on the pharmacological effects and mechanism of fucoxanthin. Yunnan Chem. Technol. 2021, 48, 4–8. [Google Scholar]
- Jin, X.D.; Wang, J.Q.; Cao, C.Q.; Wang, B.; Ding, H.M.; Wang, C.S.; Qian, G.Y. Study on the technology of extracting fucoxanthin from Hizikia fusiforme by cyclic ultrasound. Sci. Technol. Food Ind. 2021, 42, 170–178. [Google Scholar]
- Marella, T.K.; Tiwari, A. Marine diatom Thalassiosira weissflogii based biorefinery for co-production of eicosapentaenoic acid and fucoxanthin. Bioresour. Technol. 2020, 307, 123245. [Google Scholar] [CrossRef]
- Qiu, J.M.; Su, T.C.; Wang, X.; Jiang, L.; Shang, Y.; Jin, P.; Xu, J.T.; Fan, J.L.; Li, W.; Li, F.T. Comparative study of the physiological responses of Skeletonema costatum and Thalassiosira weissflogii to initial pCO2 in batch cultures, with special reference to bloom dynamics. Mar. Environ. Res. 2022, 175, 105581. [Google Scholar] [CrossRef]
- Hernández-Sandoval, F.E.; Del Ángel-Rodríguez, J.A.; Núñez-Vázquez, E.J.; Band-Schmidt, C.J.; Arredondo-Vega, B.O.; Campa-Córdova, Á.I.; Moreno-Legorreta, M.; Fernández-Herrera, L.J.; López-Cortés, D.J. Effects on cell growth, lipid and biochemical composition of Thalassiosira weissflogii (Bacillariophyceae) cultured under two nitrogen sources. Appl. Sci. 2022, 12, 961. [Google Scholar] [CrossRef]
- Dou, Y.; Shao, P.; Gao, J.W.; Jia, X.Y.; Zhou, W.L. Effects of the culture conditions on growth and photosynthesis indices of Thalassiosira weissflogii. J. Coast. Res. 2020, 115, 696. [Google Scholar] [CrossRef]
- Liang, J.J.; Jiang, X.M.; Ye, L.; Han, Q.X. Effects of nitrogen phosphorus and iron on the growth, total lipid content and fatty acid composition of Phaeodactylum tricornutum mutant strain. Chin. J. Ecol. 2016, 35, 189–198. [Google Scholar]
- Xi, Y.Q.; Chong, M.; Zhu, Q.Q.; Yang, H.; Da, W.Y. Effects of different nitrogen sources on growth and some biochemical composition of chlorella vulgaris in mixotrophy. J. N. W. Norm. Univ. 2015, 51, 82–86. [Google Scholar]
- He, S.S.; Gao, B.Y.; Lei, X.Q.; Wan, L.L.; Li, A.F.; Zhang, C.W. Effects of initial nitrogen supply on the growth, Morphology and Lipid Accumulation of oleaginous microalgae Eusyigmatos vischeri (Eustigmatophyceae). Acta Hydrobiol. Sin. 2015, 39, 574–582. [Google Scholar]
- Wang, Y.L.; Yang, H.B.; Yu, Y. Effects of sodium nitrate concentration on the growth of Platymonas subcordiformis and the contents of oil and starch. J. Anhui Agric. Sci. 2014, 42, 12247–12249. [Google Scholar]
- Zhou, N.; Cai, W.W.; Sun, D.H.; Yang, F.F. Effect of NaNO3 concentrations on growth of Haematococcus pluvialis and astaxanthin accumulation. J. Anhui Agric. Sci. 2015, 43, 232–234+239. [Google Scholar]
- Jing, Y.L.; Chang, H.L.; Christopher, Q.L.; Dan, K.L. Effects of sodium bicarbonate on cell growth, lipid accumulation, and morphology of Chlorella vulgaris. Microb. Cell Factories 2018, 17, 111. [Google Scholar]
- Gao, L.; Ding, W.Q.; Xi, J.J.; Gao, S.H.; Zhou, X.; Chen, Y.D.; Song, K.; Mao, X.R.; Tu, R.J.; Jiang, G.M. Effects of different nitrogen/phosphorus ratios on the growth and metabolism of microalgae Scenedesmus obliquus cultured in the mixed wastewater from primary settling tank and sludge thickener. Process Saf. Environ. Prot. 2023, 170, 824–833. [Google Scholar] [CrossRef]
- Dixit, R.; Singh, S.; Singh, A. Effect of nitrogen deficiency on the physiology and biochemical composition of microalga Scenedesmus rotundus-MG910488. J. Basic Microbiol. 2020, 60, 158–172. [Google Scholar] [CrossRef]
- Samantha, S.C.; Andréa, L.M.; Bianca, B.A.; Denilson, D.J.A.; Carolina, O.S.; Michele, G.D.M.; Jorge, A.V.C.; Janice, I.D. Influence of nitrogen on growth, biomass composition, production, and properties of polyhydroxyalkanoates (PHAs) by microalgae. Int. J. Biol. Macromol. 2018, 116, 552–562. [Google Scholar]
- Zhang, Y.B.; Tian, J.J.; Ye, Z.L.; Ye, Z.W.; Zhang, L.; Xun, J.L. Effects of Several Environmental Factors on Nutrient Accumulations of Nannochloropsis oceanica. J. Nucl. Agric. Sci. 2022, 36, 1273–1283. [Google Scholar]
- Liu, Z.Z.; Zhou, Z.W.; Zhang, H.; Huang, L.D.; Zhang, C.W.; Gao, B.Y. Effects of different ammonium salts and culture modes on the growth, lipid accumulation and photosynthetic physiology of Tetradesmus bernardii. Plant Physiol. J. 2021, 57, 2043–2056. [Google Scholar]
- Cointet, E.; Wielgosz-Collin, G.; Bougaran, G.; Rabesaotra, V.; Gonçalves, O.; Méléder, V. Effects of light and nitrogen availability on photosynthetic efficiency and fatty acid content of three original benthic diatom strains. PLoS ONE 2019, 14, e0224701. [Google Scholar] [CrossRef] [PubMed]
- Zhiguang, X.; Hai, Y.W.; Dong, M.Z.; Fu, X.S.; Jian, Z.S.; Guang, C.W. Combined effects of light intensity and NH4+-enrichment on growth, pigmentation, and photosynthetic performance of Ulva prolifera (Chlorophyta). Chin. J. Oceanol. Limnol. 2014, 32, 1016–1023. [Google Scholar]
- Han, T.T.; Qi, Z.H.; Huang, H.H.; Liang, Q.Y. Effects of Enrichment with Three Nitrogen Sources on Growth and Biochemical Composition of Sargassum hemiphyllum Seedlings. Prog. Fish. Sci. 2018, 39, 159–166. [Google Scholar]
- Carreto, J.I.; Catoggio, J.A. Variations in pigment contents of the diatom Phaeodactylum tricornutum during growth. Mar. Biol. 1976, 36, 105–112. [Google Scholar] [CrossRef]
- Laura, P.; Mohamed, D.; Fong, L.C.; François, P.; Louka, S.; Lekha, S.; Radovan, P. Carotenoid production and change of photosynthetic functions in Scenedesmus sp. exposed to nitrogen limitation and acetate treatment. J. Appl. Phycol. 2012, 24, 117–124. [Google Scholar]
- Pang, B.B.; Ye, F.C.; Li, Y.Y.; Shang, C.H. Research progress on carotenoid biosynthesis and metabolic regulation in microalgae. J. Guangxi Norm. Univ. 2021, 39, 13–23. [Google Scholar]
- Liang, Y.; Ji, W.W.; Shi, W.J.; Tian, C.Y.; Hu, N.X.; Yan, Y.Y. Effect of nitrogen limitation time on growth, total lipid content, and fatty acid composition of Halochlorococcum sarcotum and Nannochloris oculata. Mar. Sci. 2020, 02, 45–55. [Google Scholar]
- Tania, M.; Andressa, B.D.; Eliseu, R.; Rosane, R. Effect of temperature and nitrogen concentration on biomass composition of Heterochlorella luteoviridis. Food Sci. Technol. 2017, 37, 28–37. [Google Scholar]
- Omar, F.M.; Ilyas, M.; Mozahar, H.M.; Abdur, R.S. Influence of Nitrogen to Phosphorus Ratio and CO2 Concentration on Lipids Accumulation of Scenedesmus dimorphus for Bioenergy Production and CO2 Biofixation. Chem. Asian J. 2020, 15, 4307–4320. [Google Scholar] [CrossRef]
- Baky, H.E.; Baroty, G.E.; Mostafa, E. Optimization growth of spirulina (Arthrospira) platensis in photobioreactor under varied nitrogen concentration for maximized biomass, carotenoids and lipid contents. Recent Pat. Food Nutr. Agric. 2020, 11, 40–48. [Google Scholar] [CrossRef]
- Yongmanitchai, W.; Ward, P. Growth of and omega-3 fatty acid production by Phaeodactylum tricornutum under different culture conditions. Appl. Environ. Microbiol. 1991, 57, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, T.T.; Song, S.Q.; Li, C.W. Content and composition of fatty acid in planktonic dinoflagellate Akashiwo sanguinea cultured in different concentrations. Oceanol. Limnol. Sin. 2019, 50, 301–307. [Google Scholar]
- Zhao, P.P.; Gu, W.H.; Wu, S.C.; Xie, J.X.; Zhao, Y.P.; Huang, A.Y.; Huan, L.; Pan, G.H.; Wang, G.C. Changes in central carbon metabolism of Phaeodactylum tricornutum are beneficial for lipid accumulation under nitrogen starvation conditions. Chin. Sci. Bull. 2015, 60, 2196–2208. (In Chinese) [Google Scholar]
- Liao, L.X.; Yu, H.; Huang, J.Z. Advanced in polyunsaturated fatty acids synthesis pathway. J. Microbiol. 2014, 34, 80–85. [Google Scholar]
- Xie, L.S.; Zhang, N.; Li, C.L.; Huang, X.H. Effects of sodium nitrate on growth, biochemical components and sedimentation of Oocystis borgei. J. Guangdong Ocean. Univ. 2020, 40, 48–55. [Google Scholar]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. Determination of chlorophylls and total carotenoids: Spectrophotometric method. In Science Direct. A Manual of Chemical & Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984; pp. 101–104. [Google Scholar]
- Xu, R.R.; Gong, Y.F.; Chen, W.T.; Li, S.R.; Chen, R.Y.; Zheng, X.Y.; Chen, X.M.Z.; Wang, H.Y. Effects of LED monochromatic light quality of different colors on fucoxanthin content and expression levels of related genes in Phaeodactylum tricornutum. Acta Opt. Sin. 2019, 9, 299–307. [Google Scholar]
- Zhang, W.Y.; Gao, B.Y.; Li, A.F.; Zhang, C.W. Effects of different culture conditions on growth and accumulation of bioactive components by Phaeodactylum tricornutum. Mar. Sci. 2016, 040, 57–65. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Items | 0 N | 1 N | 2 N | 4 N |
|---|---|---|---|---|
| C4:0 | 510.09 | 72.78 | 72.43 | 95.07 |
| C8:0 | 451.62 | 205.67 | 111.82 | 144.97 |
| C10:0 | 837.58 | 91.67 | 749.12 | 897.8 |
| C11:0 | 205 | 146.07 | 80.59 | 103.76 |
| C12:0 | 845.07 | 312.87 | 128.85 | 152.21 |
| C14:0 | 202.17 | 19,855.7 | 12,896.25 | 11,319.31 |
| C14:1n5 | Not detected | 248.02 | 185.78 | 172.25 |
| C15:0 | 7911.41 | 3760.83 | 2158.12 | 1680.39 |
| C16:0 | 44,175.05 | 62,837.67 | 45,565.71 | 36,811.21 |
| C16:1n7 | 53,077.27 | 86,394.56 | 64,680 | 55,224.8 |
| C17:0 | 1385.3 | 672.74 | 443.44 | 478.44 |
| C18:0 | 1222.82 | 779.43 | 496.11 | 441.97 |
| C18:1n9c | 1640.05 | 1953.58 | 1160.62 | 875.18 |
| C18:2n6c | 1378.37 | 212.06 | 571.51 | 555.39 |
| C18:3n6 | 641.67 | 238.42 | 246.95 | 315.25 |
| C18:3n3 | Not detected | 165.81 | 260.83 | 468.74 |
| C20:0 | 799.48 | 212.06 | 106.15 | 145.1 |
| C20:2 | 2113.57 | 238.42 | 91.63 | 212.83 |
| C20:3n6 | 383.46 | 165.81 | 148.27 | 271.33 |
| C21:0 | 1090.76 | 124.29 | 96.37 | 473.16 |
| C20:4n6 | 905.97 | 1729.42 | 768.33 | 1204.2 |
| C20:5n3 | 16,058.6 | 22,672.09 | 15,140.51 | 13,953.13 |
| C22:0 | 109.23 | 412.22 | 347.82 | 332.29 |
| C22:1n9 | 511.66 | 82.83 | 232.45 | 139.27 |
| C22:2n6 | 6725.25 | 2653.1 | 1155.3 | 997.14 |
| C23:0 | 1369.49 | 809.51 | 753.65 | 569.1 |
| C24:0 | 1651.71 | 1292.15 | 833.15 | 853.62 |
| C22:6n3 | 5035.53 | 4550.28 | 4146.78 | 3852.98 |
| C24:1n9 | 501.59 | 134.97 | 103.68 | 98.63 |
| SFA | 62,766.78 | 91,585.66 | 64,839.58 | 54,498.4 |
| MUFA | 55,730.57 | 88,813.96 | 66,362.53 | 56,510.13 |
| PUFA | 33,242.42 | 32,625.41 | 22,530.11 | 21,830.99 |
| FA | 151,739.8 | 213,025 | 153,732.2 | 132,839.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rui, X.; Amenorfenyo, D.K.; Peng, K.; Li, H.; Wang, L.; Huang, X.; Li, C.; Li, F. Effects of Different Nitrogen Concentrations on Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii. Mar. Drugs 2023, 21, 106. https://doi.org/10.3390/md21020106
Rui X, Amenorfenyo DK, Peng K, Li H, Wang L, Huang X, Li C, Li F. Effects of Different Nitrogen Concentrations on Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii. Marine Drugs. 2023; 21(2):106. https://doi.org/10.3390/md21020106
Chicago/Turabian StyleRui, Xiangyu, David Kwame Amenorfenyo, Ke Peng, Haoming Li, Linfei Wang, Xianghu Huang, Changling Li, and Feng Li. 2023. "Effects of Different Nitrogen Concentrations on Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii" Marine Drugs 21, no. 2: 106. https://doi.org/10.3390/md21020106