
Paralytic Shellfish Toxins in the Gastropod Concholepas concholepas: Variability, Toxin Profiles and Mechanisms for Toxicity Reduction

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
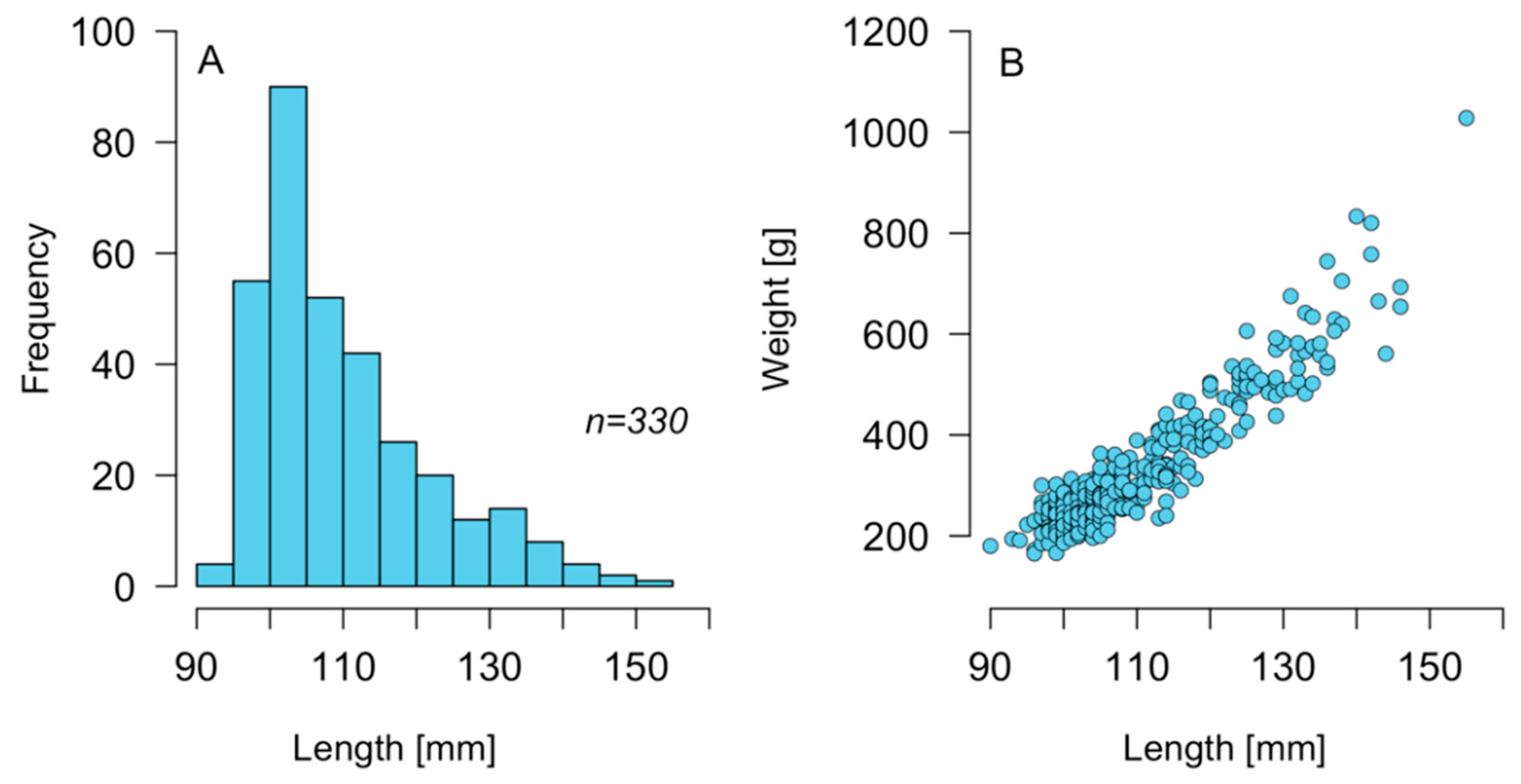
2.1. Size Structure of C. concholepas
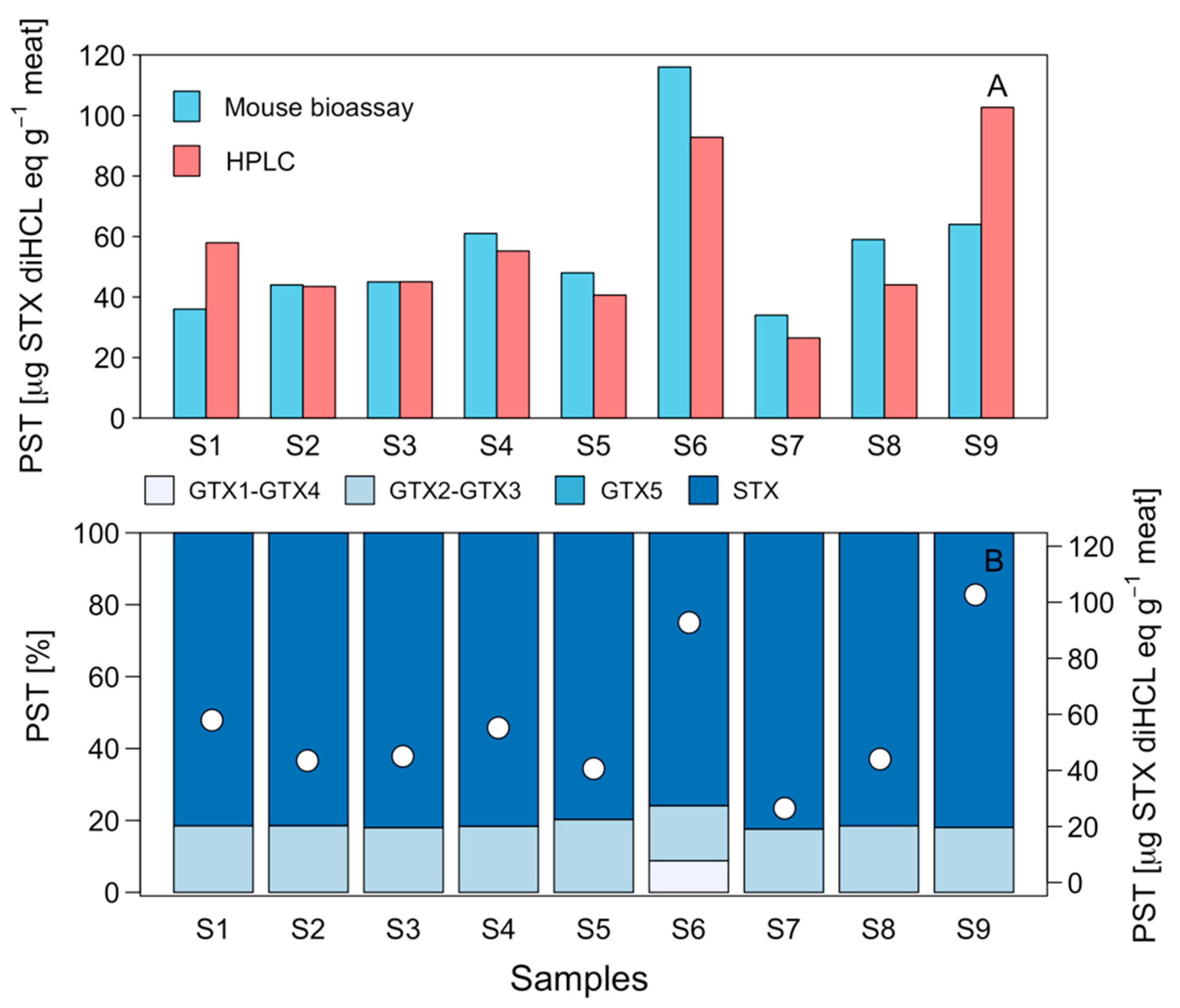
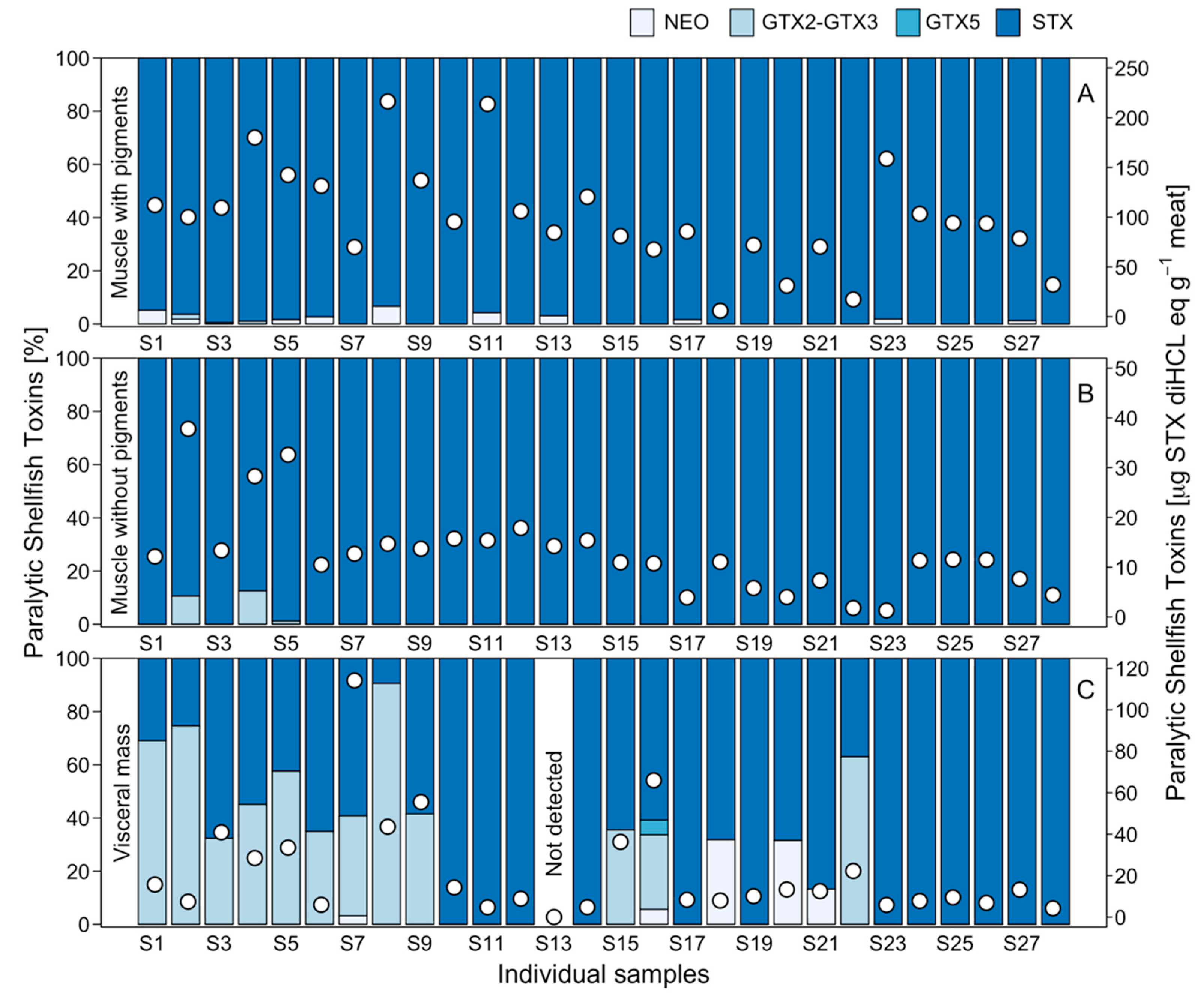
2.2. Toxicity and Toxin Profile in Potential Prey
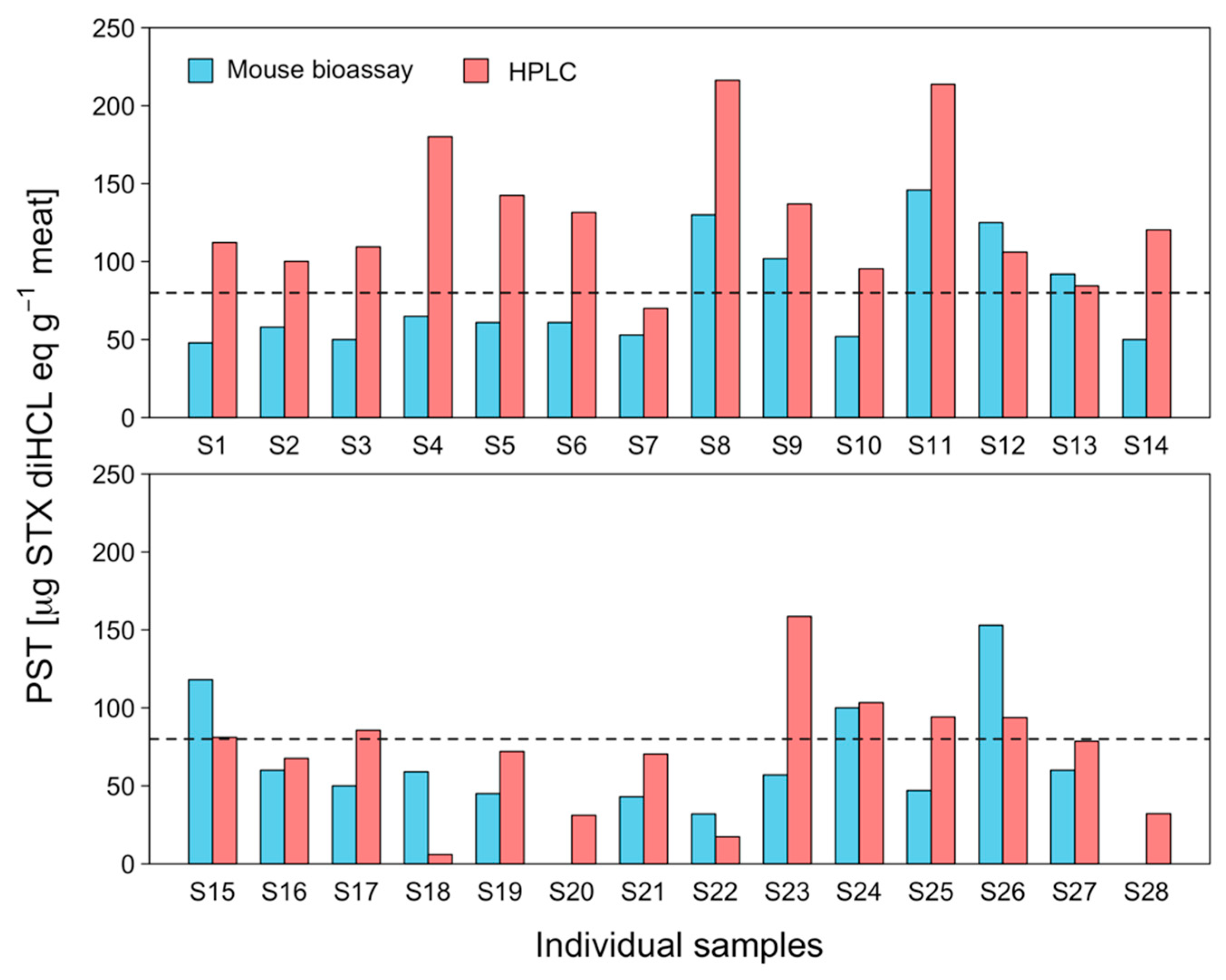
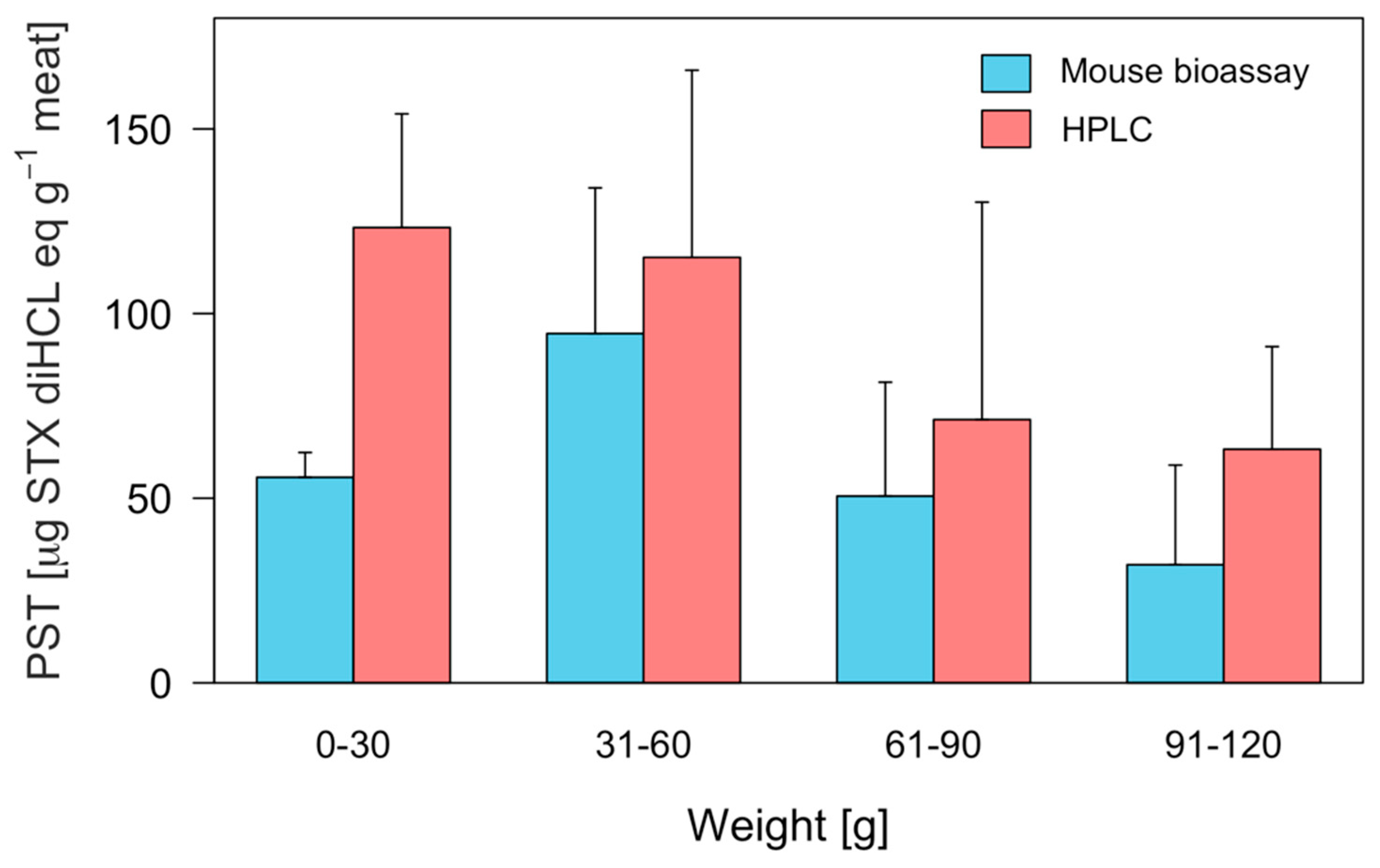
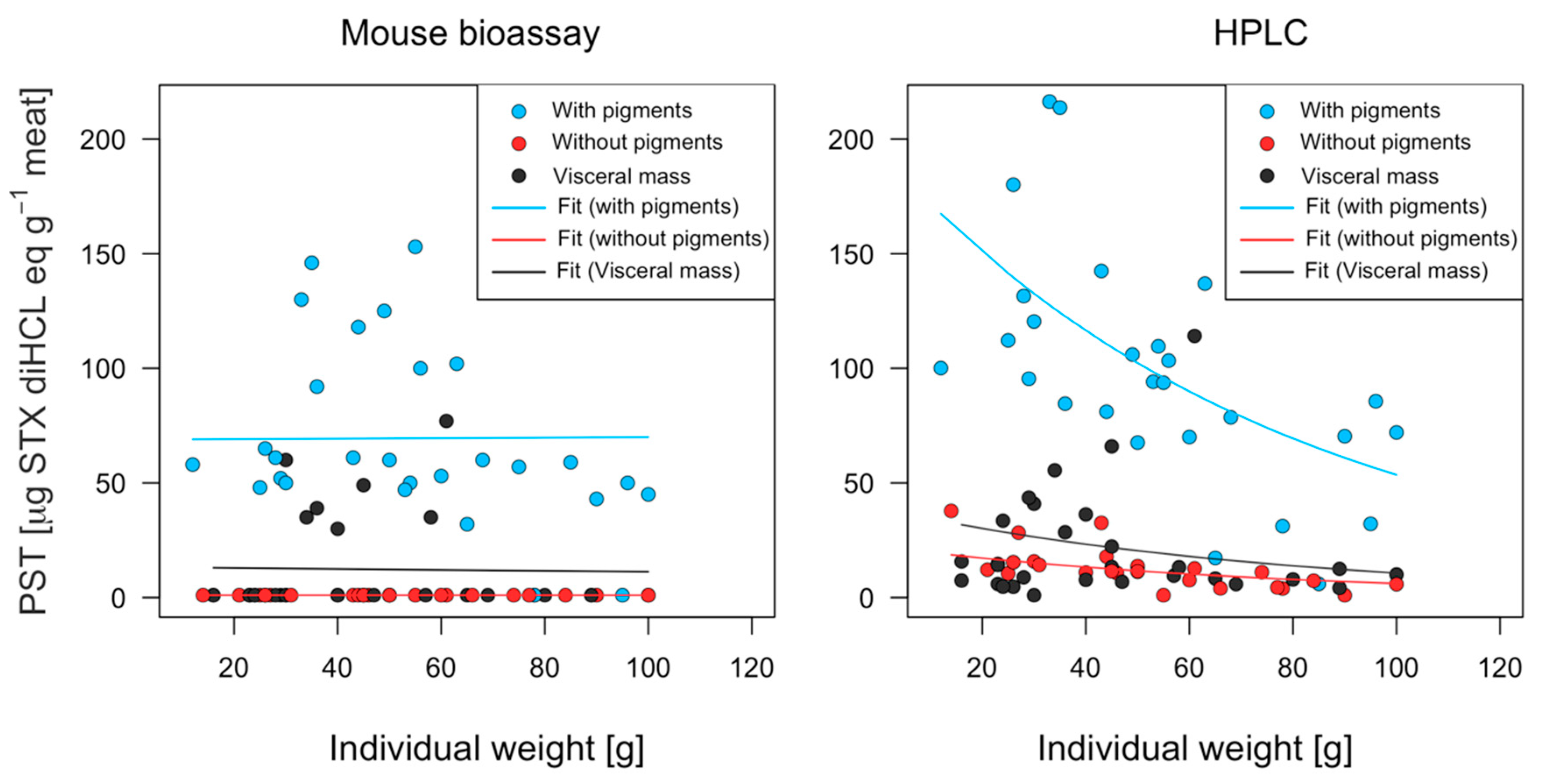
2.3. Interindividual Toxicity in Edible Tissues
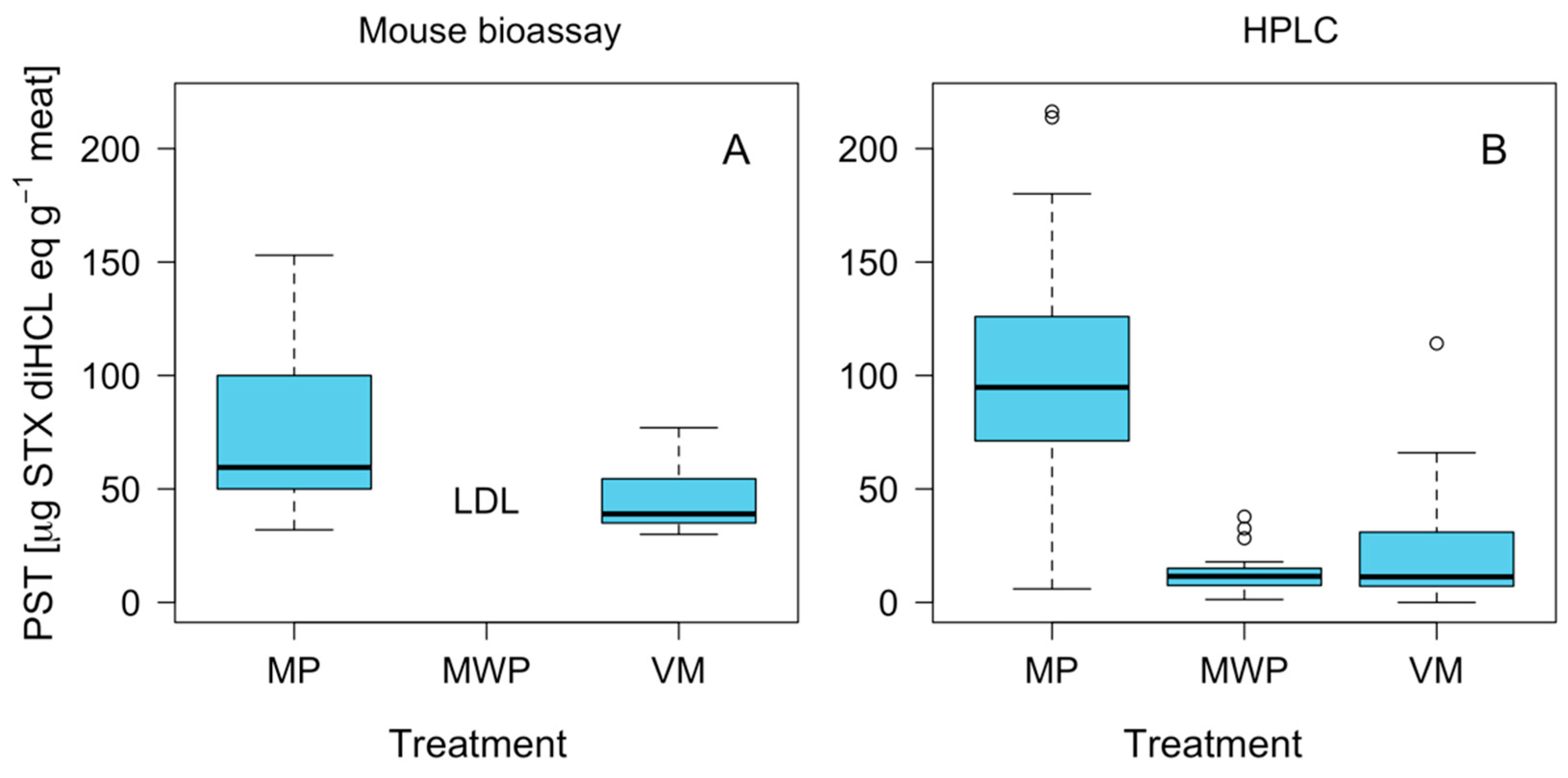
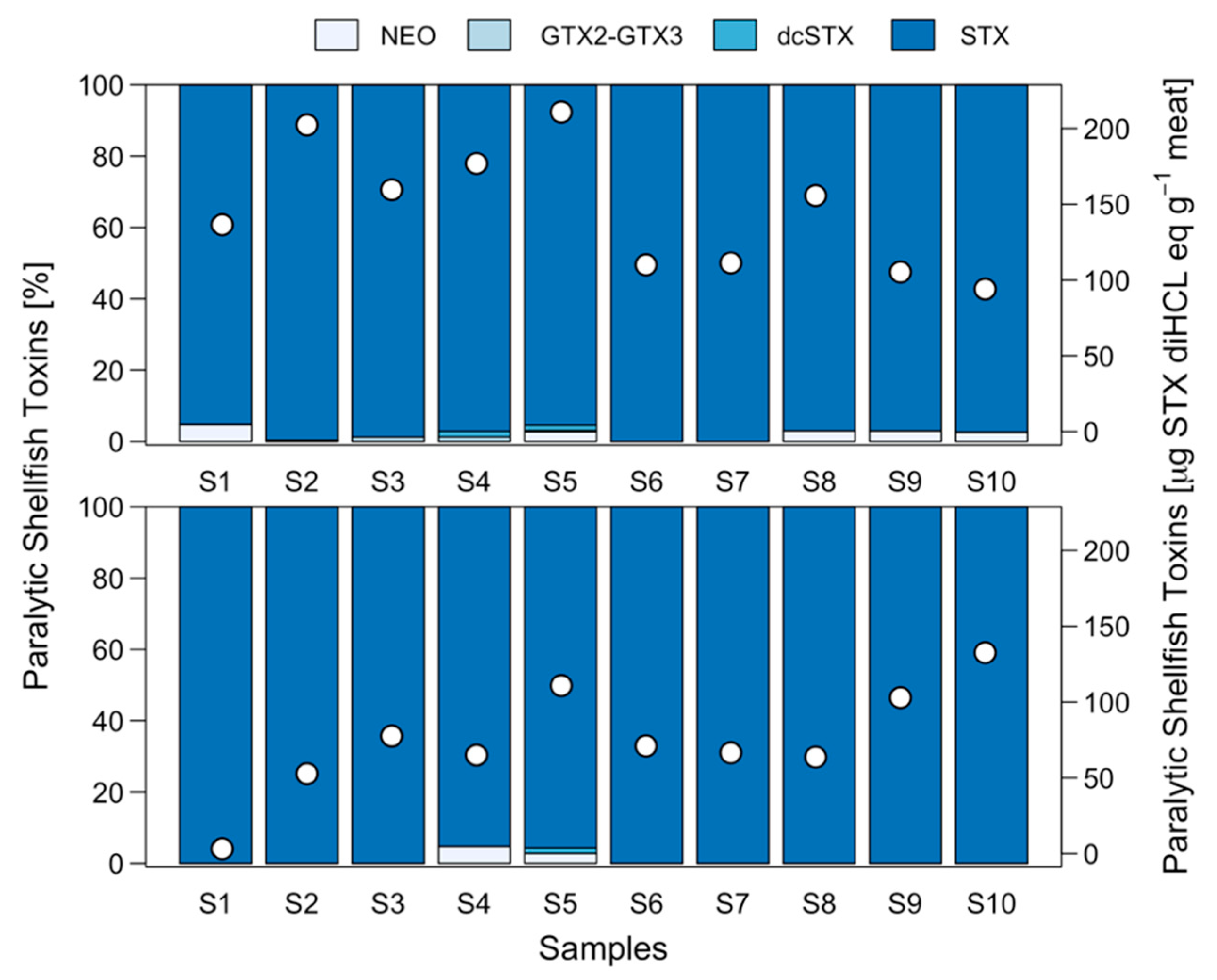
2.4. Effect of Foot Pigment Remotion Process on PST Detoxification
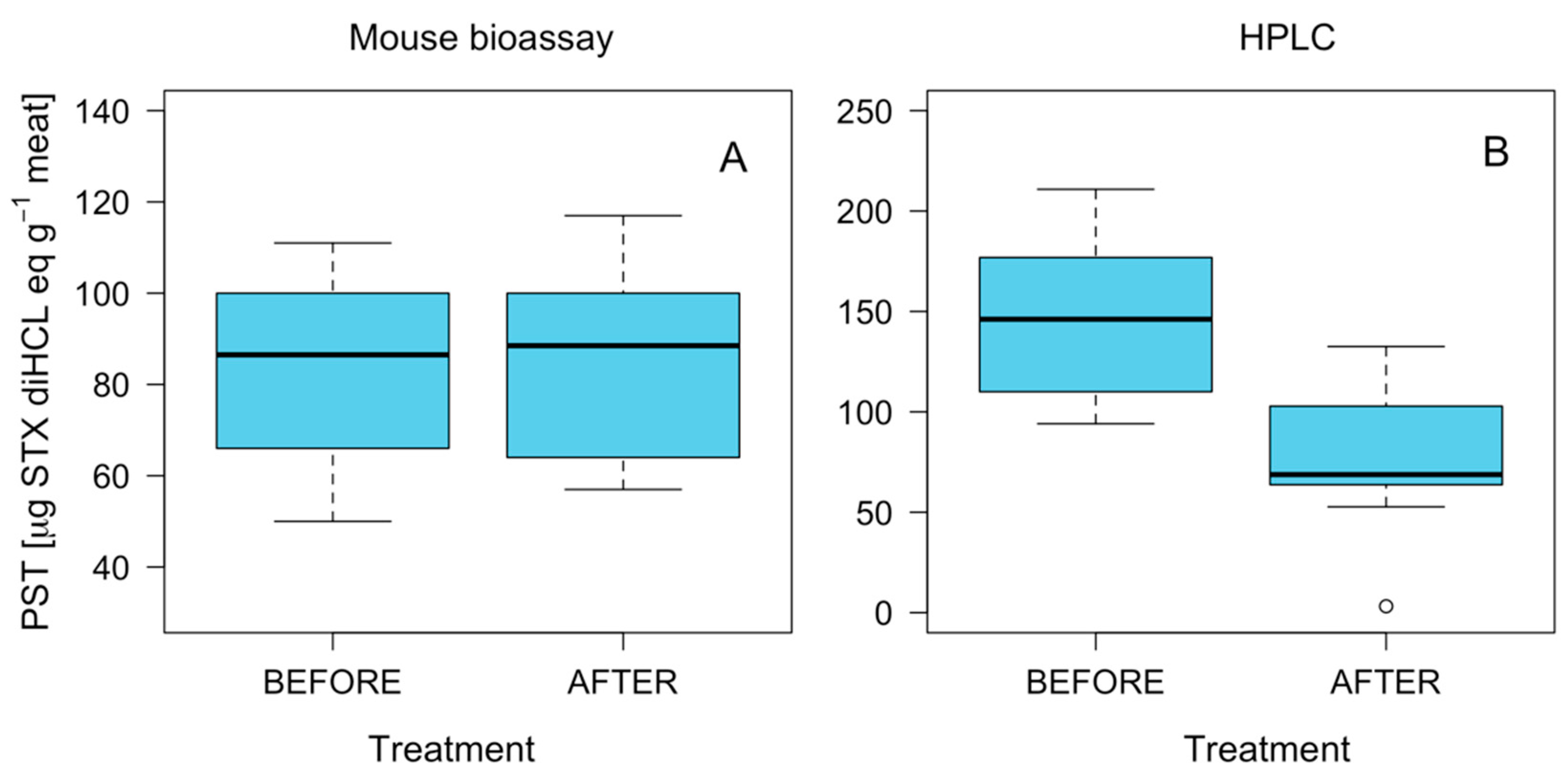
2.5. Effect of Fresh-Frozen Process on PST Detoxification
3. Discussion
3.1. Toxin Distribution and Mechanisms for Toxicity Reduction
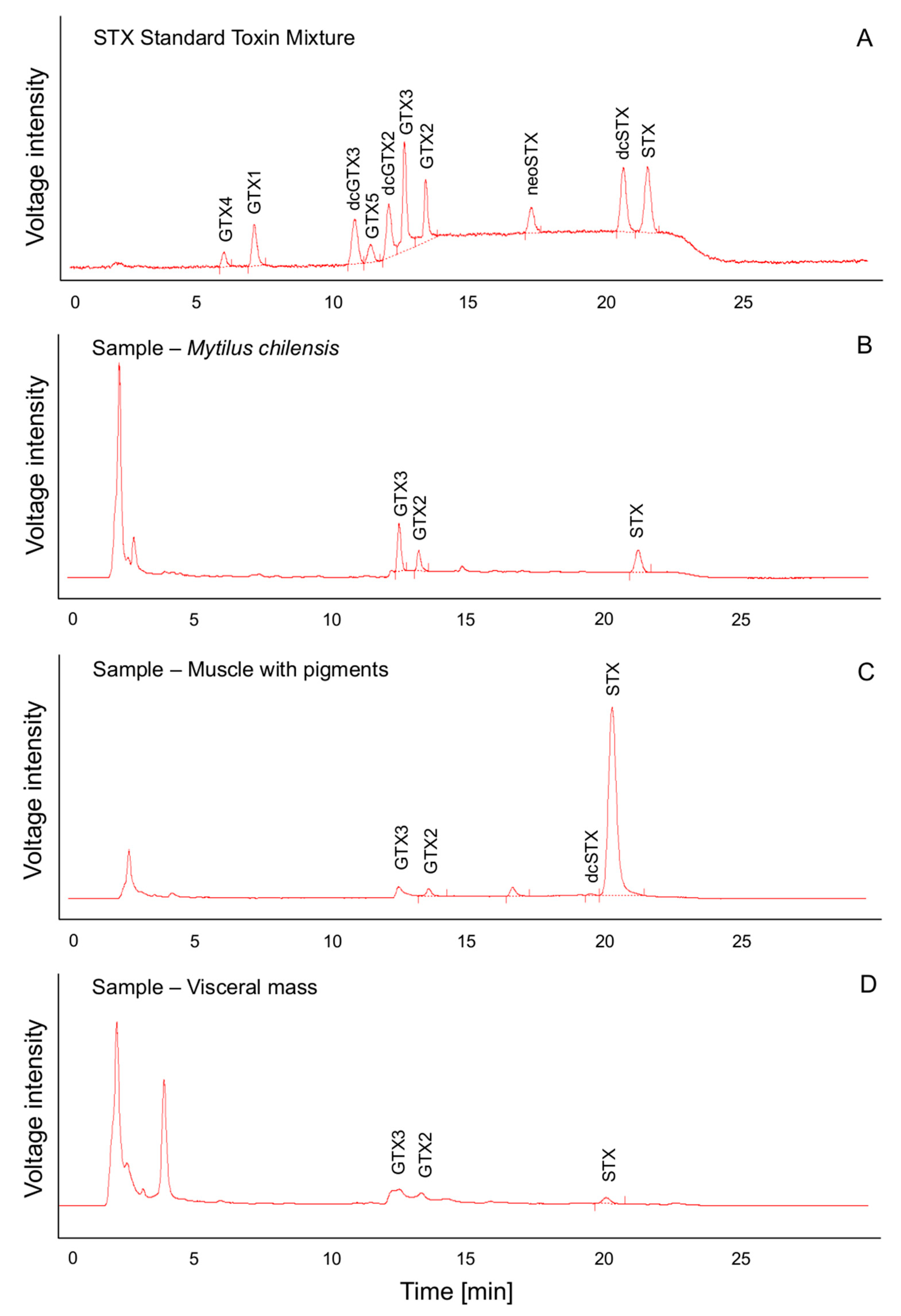
3.2. Biological and Chromatographic Analyses: Potential Implications
3.3. Implications for Management and Commercialization
4. Material and Methods
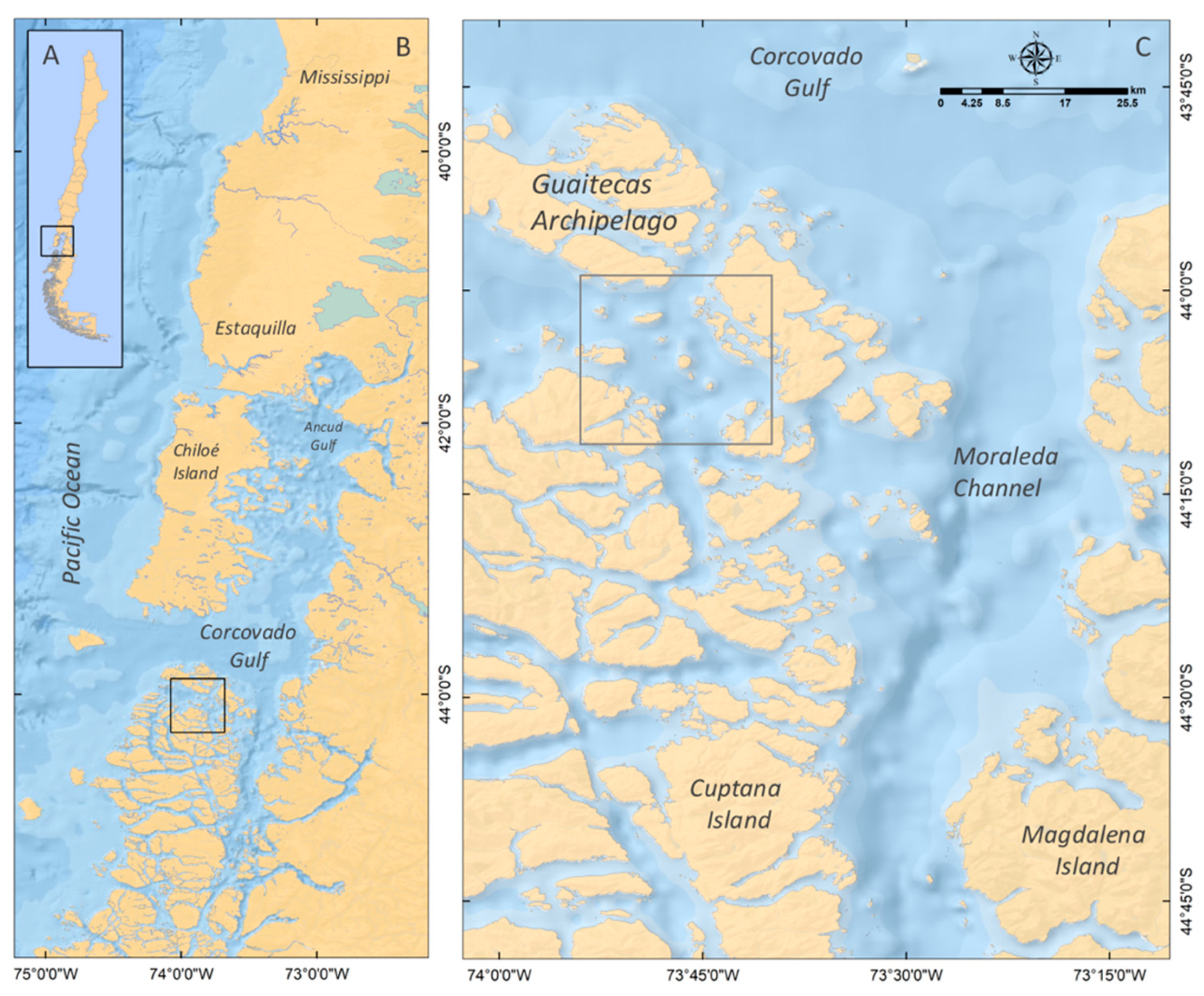
4.1. Study Area
4.2. Samples Collection
4.2.1. Experiment 1: Individual Variability of Toxicity and Effect of Pigment Extraction
4.2.2. Experiment 2: Effect of the Freezing Process
4.3. Toxicity Evaluation by Biological Method Analysis
4.4. Toxicity and Toxin Profile Determination by Chromatographic Analyses
4.5. Statistical Analysis and Graphic Representations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Mons, M.N.; Van Egmond, H.P.; Speijers, G.J.A. Paralytic Shellfish Poisoning: A Review. RIVM rapport 388802005. 1998, pp. 1–47. Available online: https://rivm.openrepository.com/handle/10029/10000 (accessed on 30 September 2022).
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M.; Blackburn, S.I.; Doblin, M.A.; Bolch, C.J.S. Global toxicology, ecophysiology and population relationships of the chainforming PST dinoflagellate Gymnodinium catenatum. Harmful Algae 2012, 14, 130–143. [Google Scholar] [CrossRef]
- Mertens, K.N.; Wolny, J.; Carbonell-Moore, C.; Bogus, K.; Ellegaard, M.; Limoges, A.; de Vernal, A.; Gurdebeke, P.; Omura, T.; Al-Muftah, A.; et al. Taxonomic re-examination of the toxic armored dinoflagellate Pyrodinium bahamense Plate 1906: Can morphology or LSU sequencing separate P. bahamense var. compressum from var. bahamense? Harmful Algae 2015, 41, 1–24. [Google Scholar] [CrossRef]
- Shin, H.H.; Li, Z.; Réveillon, D.; Rovillon, G.-A.; Mertens, K.N.; Hess, P.; Kim, H.J.; Lee, J.; Lee, K.-W.; Kim, D.; et al. Centrodinium punctatum (Dinophyceae) produces significant levels of saxitoxin and related analogs. Harmful Algae 2020, 100, 101923. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D′agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, Y. Chemical and enzymatic transformation of paralytic shellfish toxins in marine organisms. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard-Le Denn, E., Gentien, P., Marcaillou-Le Baut, C., Eds.; Lavoisier Publishing ILP: Paris, France, 1995; pp. 475–480. [Google Scholar]
- Negri, A.; Stirling, D.; Quilliam, M.; Blackburn, S.; Bolch, C.; Burton, I.; Eaglesham, G.; Thomas, K.; Walter, J.; Willis, R. Three novel hydroxybenzoate saxitoxin analogues isolated from the dinoflagellate Gymnodinium Catenatum. Chem. Res. Toxicol. 2003, 16, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numano, S.; Kudo, Y.; Cho, Y.; Konoki, K.; Kaga, Y.; Nagasawa, K.; Yotsu-Yamashita, M. Two new skeletal analogues of saxitoxin found in the scallop, Patinopecten yessoensis, as possible metabolites of paralytic shellfish toxins. Chemosphere 2021, 278, 130224. [Google Scholar] [CrossRef]
- Blanco, J.; Moroño, A.; Franco, J.; Reyero, M.I. PSP detoxification kinetics in the mussel Mytilus galloprovincialis. One- and two- comportment models and the effect of some environmental variables. Mar. Ecol. Prog. Ser. 1997, 158, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.; Reyero, M.; Franco, J. Kinetics of accumulation and transformation of paralytic shellfish toxins in the blue mussel Mytilus galloprovincialis. Toxicon 2003, 42, 777–784. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Cembella, A.D.; Laby, D. Temperature effects on kinetics of paralytic shellfish toxin elimination in Atlantic surfclams, Spisula solidissima. Deep Sea Res. II 2014, 103, 308–317. [Google Scholar] [CrossRef]
- Díaz, P.A.; Molinet, C.; Seguel, M.; Niklitschek, E.J.; Díaz, M.; Álvarez, G.; Pérez-Santos, I.; Varela, D.; Guzmán, L.; Rodríguez-Villegas, C.; et al. Modelling the spatial and temporal dynamics of paralytic shellfish toxins (PST) at different scales: Implications for research and management. Toxins 2022, 14, 786. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion of the panel on contaminants in the food chain on a request from the European commission on marine biotoxins in shellfish-summary on regulated marine biotoxins. EFSA J. 2009, 1306, 1–23. [Google Scholar]
- Shumway, S.E. A review of the effects of algal blooms on shellfish and aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Álvarez, G.; Rengel, J.; Araya, M.; Álvarez, F.; Pino, R.; Uribe, E.; Díaz, P.A.; Rossignoli, A.E.; López-Rivera, A.; Blanco, J. Rapid domoic acid depuration in the scallop Argopecten purpuratus and Its transfer from the digestive gland to other organs. Toxins 2020, 12, 698. [Google Scholar] [CrossRef] [PubMed]
- Bricelj, V.M.; Lee, J.H.; Cembella, A.D. Influence of dinoflagellate cell toxicity on uptake and loss of paralytic shellfish toxins in the northern quahog Mercenaria mercenaria. Mar. Ecol. Prog. Ser. 1991, 74, 33–46. [Google Scholar] [CrossRef]
- Choi, M.C.I.; Hsieh, D.P.H.; Lam, P.K.S.; Wang, W.X. Field depuration and biotransformation of paralytic shellfish toxins in scallop Chlamys nobilis and green-lipped mussel Perna viridis. Mar. Biol. 2003, 143, 927–934. [Google Scholar] [CrossRef]
- Shumway, S.E.; Sherman, S.A.; Cembella, A.D.; Selvin, R. Accumulation of paralytic shellfish toxins by surf-clams, Spisula solidissima (Dillwyn, 1897) in the Gulf of Maine: Seasonal changes, distribution between tissues, and notes on feeding habits. Nat. Tox. 1994, 2, 236–251. [Google Scholar] [CrossRef]
- Shumway, S. Phycotoxin-related shellfish poisoning: Bivalve molluscs are not the only vectors. Rev. Fish. Sci. 1995, 3, 1–31. [Google Scholar] [CrossRef]
- Ito, K.; Asakawa, M.; Beppu, R.; Takayama, H.; Miyazawa, K. PSP-toxicification of the carnivorous gastropod Rapana venosa inhabiting the estuary of Nikoh River, Hiroshima Bay, Hiroshima Prefecture, Japan. Mar. Pollut. Bull. 2004, 48, 1116–1121. [Google Scholar] [CrossRef]
- Kotaki, Y.; Oshima, Y.; Yasumoto, T. Analysis of paralytic shellfish toxins of marine snails. Bull. Japan. Soc. Sci. Fish. 1981, 47, 943–946. [Google Scholar] [CrossRef] [Green Version]
- Jen, H.-C.; Nguyen, T.A.-T.; Wu, Y.-J.; Hoang, T.; Arakawa, O.; Lin, W.-F.; Hwang, D.-F. Tetrodotoxin and paralytic shellfish poisons in gastropod species from Vietnam analyzed by high-performance liquid chromatography and liquid chromatography–tandem mass spectrometry. J. Food Drug Anal. 2014, 22, 178–188. [Google Scholar] [CrossRef]
- Hwang, P.A.; Tsai, Y.H.; Lu, Y.H. Paralytic toxins in three new gastropod (Olividae) species implicated in food poisoning in southern Taiwan. Toxicon 2003, 41, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, S. Report of 8 incidents of intoxication due to Nassarius (Zeuxis) succinctus. Chin. J. Surg. 1981, 20, 231–232. [Google Scholar]
- Montoya, N.G. Toxinas paralizantes de moluscos en el mar argentino: Impacto, transferencia trofica y perspectiva. Mar. Fish. Sci. 2019, 32, 47–69. [Google Scholar] [CrossRef]
- Turner, A.D.; Tarnovius, S.; Goya, A.B. Paralytic shellfish toxins in the marine gastropods Zidona dufresnei and Adelomelon beckii from Argentina: Toxicity and toxin profiles. J. Shellfish Res. 2014, 33, 519–530. [Google Scholar] [CrossRef]
- Compagnon, D.; Lembeye, G.; Marcos, N.; Ruiz-Tagle, N.; Lagos, N. Accumulation of paralitic shellfish poisoning toxins in the bivalve Aulacomya ater and two carnivorous gastropods Concholepas concholepas and Argobuccinum ranelliformes during an Alexandrium catenella bloom in southern Chile. J. Shellfish Res. 1998, 17, 67–73. [Google Scholar]
- Castilla, J.C.; Guiñez, R. Disjoint geographical distribution of intertidal and nearshore benthic invertebrates in the southern hemisphere. Rev. Chil. Hist. Nat. 2000, 73, 585–603. [Google Scholar] [CrossRef] [Green Version]
- Stotz, W. The management areas in the fishery Law: First experiences and evaluation of utility as a management tool for Concholepas concholepas. Est. Ocea. 1997, 16, 67–86. [Google Scholar]
- Castilla, J.C.; Espinosa, J.; Yamashiro, C.; Melo, O.; Gelcich, S. Telecoupling between catch, farming, and international trade for the gastropods Concholepas concholepas (loco), and Haliotis spp. (abalone). J. Shellfish Res. 2016, 35, 499–506. [Google Scholar] [CrossRef]
- Castilla, J.C.; Fernandez, M. Small-scale benthic fisheries in Chile: On co-management and sustainable use of benthic invertebrates. Ecol. Appl. 1998, 8, 124–132. [Google Scholar] [CrossRef]
- Cerda, O.; Stotz, W.B. Can the territorial use rights in fisheries (TURF) stabilize the landings of a highly variable benthic resource? Reexamining the fishery of Concholepas concholepas in North-Central Chile. Ocean Coast. Manag. 2022, 224, 106158. [Google Scholar] [CrossRef]
- Castilla, J.C.; Gelcich, S. The management of the loco (Concholepas concholepas) as a driver of self-governance of small-scale benthic fisheries in Chile. In Case Studies in Fisheries Self-Governance; FAO FisheriesTechnical Paper; Townsend, R., Shotton, R., Uchida, H., Eds.; FAO: Rome, Italy, 2008; Volume 504. [Google Scholar]
- Marín, A.; Berkes, F. Network approach for understanding small-scale fisheries governance: The case of the Chilean coastal co-management system. Mar. Policy 2010, 34, 851–858. [Google Scholar] [CrossRef]
- Sernapesca. Anuario Estadistico de Pesca; Servicio Nacional de Pesca: Valparaíso, Chile, 2021. [Google Scholar]
- Instituto de Salud Pública. Informes de Resultados del Programa Nacional de Vigilancia Ycontrol de las Intoxicaciones por Fenómenos Algales Nocivos; Instituto de Salud Pública, Gobierno de Chile: Santiago, Chile, 2013; Volume 15, Available online: http://www.ispch.cl/lab_amb/ (accessed on 22 September 2022).
- Zamorano, R.; Marín, M.; Cabrera, F.; Figueroa, D.; Contreras, C.; Barriga, A.; Lagos, N.; García, C. Determination of the variability of both hydrophilic and lipophilic toxins in endemic wild bivalves and carnivorous gastropods from the Southern part of Chile. Food Addit. Contam. Part A 2013, 30, 1660–1677. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Pérez, F.; Contreras, C.; Figueroa, D.; Barriga, A.; López-Rivera, A.; Araneda, O.F.; Contreras, H.R. Saxitoxins and okadaic acid group: Accumulation ans distribution in invertebrate vectors from Southern Chile. Food Addit. Contam. Part A 2015, 32, 984–1002. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, G.; Díaz, P.A.; Godoy, M.; Araya, M.; Ganuza, I.; Pino, R.; Álvarez, F.; Rengel, J.; Hernández, C.; Uribe, E.; et al. Paralytic Shellfish Toxins in Mesodesma donacium during an exceptional bloom of Alexandrium catenella associated to an intense mass mortality. Toxins 2019, 11, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armijo, J.; Oerder, V.; Auger, P.; Bravo, A.; Molina, E. The 2016 red tide crisis in southern Chile: Possible in uence of the mass oceanic dumping of dead salmons. Mar. Pollut. Bull. 2020, 150, 110603. [Google Scholar] [CrossRef]
- Díaz, P.A.; Álvarez, A.; Varela, D.; Pérez-Santos, I.; Díaz, M.; Molinet, C.; Seguel, M.; Aguilera-Belmonte, A.; Guzmán, L.; Uribe, E.; et al. Impacts of harmful algal blooms on the aquaculture industry: Chile as a case study. Perspect. Phycol. 2019, 6, 39–50. [Google Scholar] [CrossRef]
- Díaz, P.A.; Molinet, C.; Seguel, M.; Díaz, M.; Labra, G.; Figueroa, R. Coupling planktonic and benthic shifts during a bloom of Alexandrium catenella in southern Chile: Implications for bloom dynamics and recurrence. Harmful Algae 2014, 40, 9–22. [Google Scholar] [CrossRef]
- Seguel, M.; Díaz, P.A.; Molinet, C.; Díaz, M.; García, C.; Millanao, M.; Castillo, M. Estudio de alternativas tecnológicas para el procesamiento del loco Concholepas concholepas. In Ministerio de Salud, Instituto de Salud Pública—Universidad Austral de Chile; Informe Final: Puerto Montt, Chile, 2019; p. 67. [Google Scholar]
- Goya, A. Tratamientos tecnológicos para reducir la toxicidad en moluscos. Rev. Infopesca Int. 2011, 45, 24–29. [Google Scholar]
- Kelly, C.; Ortega, S.; Goya, A. Concentración de toxinas paralizantes de los moluscos (TPM) en caracol de mar Zidona dufresnei pre y post tratamiento térmico. La Industria Cárnica Latinoamericana NO. 2015, 194, 52–55. Available online: http://wwww.publitec.com.ar/contenido/objetos/Concentraciondetoxinas.pdf (accessed on 22 September 2022).
- Bravo, I.; Reyero, M.I.; Cacho, E.; Franco, J.M. Paralytic shellfish poisoning in Haliotis tuberculata from the Galician coast: Geographical distribution, toxicity by lengths and parts of the mollusc. Aquat. Toxicol. 1999, 46, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Bravo, I.; Franco, J.M.; Alonso, A.; Dietrich, R.; Molist, P. Cytological study and immunohistochemical location of PSP toxins in foot skin of the ormer, Haliotis tuberculata, from the Galician coast NW Spain. Mar. Biol. 2001, 138, 709–715. [Google Scholar] [CrossRef]
- Martinez, A.; Franco, J.M.; Bravo, I.; Mazoy, M.; Cacho, E. PSP toxicity in Haliotis tuberculata from NW Spain. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: New York, NY, USA, 1993; pp. 419–453. [Google Scholar]
- Oyaneder-Terrazas, J.; Contreras, H.R.; García, C. Prevalence, variability and bioconcentration of saxitoxin-group in different marine species present in the food chain. Toxins 2017, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Romero, P. Análisis del Mercado de Exportación del Producto Loco a Sus Principales Destinos; Technical Report; Instituto de Fomento Pesquero: Valparaíso, Chile, 2020. [Google Scholar]
- Maldonado, R. Estudio macroscópico, microscópico e histológico de Concholepas concholepas. Rev. Biol. Mar. 1965, 11, 121–127. [Google Scholar]
- Castilla, J.C.; Guisado, C.; Cancino, J. Aspectos ecológicos y conductuales relacionados con la alimentación de Concholepas concholepas (Mollusca: Gastropoda: Muricidae). Biol. Pesq. 1979, 12, 99–144. [Google Scholar]
- Molinet, C.; Lembeye, G.; González, C. Análisis y monitoreo de la marea roja en el recurso loco (Concholepas concholepas) en el litoral de Aysén. In Informe Final Proyecto Fondo Nacional de Desarrollo Regional; Universidad Austral de Chile: Coyhaique, Chile, 1998; p. 45. [Google Scholar]
- EC. Commission Decision of 18 January 1996 establishing the conditions for the harvesting and processing of certain bivalve molluscs coming from areas where the paralytic shellfish poison level exceeds the limit laid down by Council Directive 91/492/EEC (96/77/EC). Off. J. Eur. Comm. 1996, L15, 46–47. [Google Scholar]
- Cabado, A.G.; Lago, J.; González, V.; Blanco, L.; Paz, B.; Diogène, J.; Ferreres, L.; Rambla-Alegre, M. Detoxification of paralytic shellfish poisoning toxins in naturally contaminated mussels, clams and scallops by an industrial procedure. Food Chem. Toxicol. 2020, 141, 111386. [Google Scholar] [CrossRef] [PubMed]
- Vieites, J.M.; Botana, L.M.; Vieites, M.R.; Leira, F.J. Canning process that di-minishes paralytic shellfish poison in naturally contaminated mussels (Mytilus galloprovincialis). J. Food Prot. 1999, 62, 515–519. [Google Scholar] [CrossRef]
- Reboreda, A.; Lago, J.; Chapela, M.-J.; Vieites, J.M.; Botana, L.M.; Alfonso, A.; Cabado, A. Decrease of marine toxin content in bivalves by industrial process. Toxicon 2010, 55, 235–243. [Google Scholar] [CrossRef]
- Dowsett, N.; Hallegraeff, G.; van Ruth, P.; van Ginkel, R.; McNabb, P.; Hay, B.; O′Connor, W.; kiermeier, A.; Deveney, M.; McLeod, C. Uptake, distribution and depuration of paralytic shellfish toxins from Alexandrium minutum in Australian greenlip abalone, Haliotis laevigata. Toxicon 2011, 58, 101–111. [Google Scholar] [CrossRef]
- Seger, A.; Hallegraeff, G.; Stone, D.A.J.; Bansemer, M.S.; Harwood, D.T.; Turnbull, A. Uptake of Paralytic Shellfish Toxins by Blacklip Abalone (Haliotis rubra rubra Leach) from direct exposure to Alexandrium catenella microalgal cells and toxic aquaculture feed. Harmful Algae 2020, 99, 101925. [Google Scholar] [CrossRef]
- Blanco, J.; Correa, J.; Muñíz, S.; Mariño, C.; Martín, H.; Arévalo, F. Evaluación del impacto de los métodos y niveles utilizados para el control de toxinas en el mejillón. Rev. Galega Dos Recur. Mariños 2013, 3, 1–55. [Google Scholar]
- Hignutt, E. Suitability of postcolumn oxidation liquid chromatography method AOAC 2011.02 for monitoring paralytic shellfish toxins in Alaskan shellfish—Initial pilot study versus mouse bioassay and in-house validation. J. AOAC Int. 2014, 97, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Tarnovius, S.; Hatfield, R.G.; Teixeira-Alves, M.; Broadwater, M.; Dolah, F.V.; Garcia-Mendoza, E.; Medina, D.; Salhi, M.; Goya, A.B.; et al. Application of six detection methods for analysis of paralytic shellfish toxins in shellfish from four regions within Latin America. Mar. Drugs 2020, 18, 616. [Google Scholar] [CrossRef] [PubMed]
- Gelcich, S.; Edwards-Jones, G.; Kaiser, M.J.; Castilla, J.C. Co-management policy can reduce resilience in traditionally managed marine ecosystems. Ecosystems 2006, 9, 951–966. [Google Scholar] [CrossRef]
- Aranceta-Garza, F.; Pérez-Enríquez, R.; Cruz, P. PCR-SSCP method for genetic differentiation of canned abalone and commercial gastropods in the Mexican retail market. Food Control 2011, 22, 1015–1020. [Google Scholar] [CrossRef]
- Oyanedel, R.; Keim, A.; Castilla, J.C.; Gelcich, S. Illegal fishing and territorial user rights in Chile. Conserv. Biol. 2018, 32, 619–627. [Google Scholar] [CrossRef]
- Pickard, G.L. Some physical oceanographic features of inlets of Chile. J. Fish. Res. Board Can. 1971, 28, 1077–1106. [Google Scholar] [CrossRef]
- Anon, S. AOAC Official Method 959.08. Paralytic shellfish poison. Biological method. Final action. In AOAC Official Methods for Analysis; Truckses, M.W., Ed.; AOAC International: Gaithersburg, MD, USA, 2008; pp. 79–80. [Google Scholar]
- van de Riet, J.; Gibbs, R.S.; Muggah, P.M.; Rourke, W.A.; MacNeil, J.D.; Quilliam, M.A. Liquid Chromatography Post-Column Oxidation (PCOX) method for the determination of Paralytic Shellfish Toxins in mussels, clams, and scallops: Collaborative study. J. AOAC Int. 2011, 94, 1154–1176. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Social. Method. Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. Available online: http://www.r-project.org/ (accessed on 30 September 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 25 September 2022).
- Zeileis, A.; Hothorn, T. Diagnostic checking in regression relationships. R News 2002, 2, 7–10. Available online: https://CRAN.R-project.org/doc/Rnews/ (accessed on 15 August 2022).
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version. 2020. Available online: https://cran.r-project.org (accessed on 30 September 2022).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model/Hypotheses | K | AIC | Delta AIC | AIC Wt | Cum. Wt | LL |
|---|---|---|---|---|---|---|
| Mouse bioassay | ||||||
| GLM Gamma: Toxicity = Treatment | 4 | 554.93 * | 0 | 0.63 | 0.63 | −273.21 |
| GLM Gamma: Toxicity = Individual weight + Treatment | 5 | 555.99 | 1.06 | 0.37 | 1 | −272.61 |
| GLM negative binomial: Toxicity = Treatment | 4 | 578.28 | 23.35 | 0 | 1 | −284.89 |
| GLM negative binomial: Toxicity = Individual weight + Treatment | 5 | 579.59 | 24.65 | 0 | 1 | −284.41 |
| Linear model: Toxicity = Individual weight + Treatment | 7 | 784.93 | 229.99 | 0 | 1 | −384.73 |
| Linear model: Toxicity = Treatment | 3 | 858.73 | 303.8 | 0 | 1 | −426.22 |
| GLM poison: Toxicity = Individual weight + Treatment | 4 | 1654.87 | 1099.93 | 0 | 1 | −823.18 |
| HPLC-FLD PCOX | ||||||
| GLM negative binomial: Toxicity = Individual weight + Treatment | 5 | 727.15 * | 0 | 0.93 | 0.93 | −358.19 |
| GLM Gamma: Toxicity = Individual weight + Treatment | 5 | 732.83 | 5.67 | 0.05 | 0.98 | −361.03 |
| GLM Gamma: Toxicity = Treatment | 4 | 736.30 | 9.15 | 0.01 | 0.99 | −363.90 |
| GLM negative binomial: Toxicity = Treatment | 4 | 736.62 | 9.46 | 0.01 | 1 | −364.06 |
| Linear model: Toxicity = Individual weight + Treatment | 7 | 815.59 | 88.44 | 0 | 1 | −400.06 |
| Linear model: Toxicity = Treatment | 4 | 832.54 | 105.39 | 0 | 1 | −412.02 |
| GLM poison: Toxicity = Individual weight + Treatment | 3 | Inf | Inf | 0 | 1 | −449.77 |
| GLM poison: Toxicity = Treatment | 4 | Inf | Inf | 0 | 1 | Inf |
| Degree of Freedom | Deviance | Residual Degree of Freedom | Residual Deviance | Pr(>Chi) | |
| NULL | 83 | 226.233 | |||
| Individual weight | 1 | 0.13 | 82 | 226.103 | 0.7185 |
| Treatment | 2 | 144.35 | 80 | 81.757 | <2 × 10−16 |
| Linear hypotheses: | |||||
| Treatment factor level | Estimate | Std. error | z value | Pr(>|z|) | |
| Brushed foot − no brushed foot = 0 | 4.2274 | 0.3405 | −12.415 | <1 × 10−8 | |
| Visceral mas − no brushed foot = 0 | 1.7131 | 0.2883 | −5.942 | <1 × 10−8 | |
| Visceral mass − brushed foot = 0 | 2.5142 | 0.344 | 7.31 | <1 × 10−8 | |
| Degree of Freedom | Deviance | Residual Degree of Freedom | Residual Deviance | Pr(>Chi) | |
| NULL | 83 | 245.056 | |||
| Individual weight | 1 | 3.73 | 82 | 241.326 | 0.05345 |
| Treatment | 2 | 151.98 | 80 | 89.349 | <2 × 10−16 |
| Linear hypotheses: | |||||
| Treatment factor level | Estimate | Std. error | z value | Pr(>|z|) | |
| Brushed foot − no brushed foot = 0 | −2.1705 | 0.1879 | −11.553 | <0.001 | |
| Visceral mass − no brushed foot = 0 | −1.6108 | 0.1859 | −8.665 | <0.001 | |
| Visceral mass − brushed foot = 0 | 0.5597 | 0.1907 | 2.936 | 0.00933 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seguel, M.; Molinet, C.; Díaz, M.; Álvarez, G.; García, C.; Marín, A.; Millanao, M.O.; Díaz, P.A. Paralytic Shellfish Toxins in the Gastropod Concholepas concholepas: Variability, Toxin Profiles and Mechanisms for Toxicity Reduction. Mar. Drugs 2023, 21, 44. https://doi.org/10.3390/md21010044
Seguel M, Molinet C, Díaz M, Álvarez G, García C, Marín A, Millanao MO, Díaz PA. Paralytic Shellfish Toxins in the Gastropod Concholepas concholepas: Variability, Toxin Profiles and Mechanisms for Toxicity Reduction. Marine Drugs. 2023; 21(1):44. https://doi.org/10.3390/md21010044
Chicago/Turabian StyleSeguel, Miriam, Carlos Molinet, Manuel Díaz, Gonzalo Álvarez, Carlos García, Andrés Marín, María Olga Millanao, and Patricio A. Díaz. 2023. "Paralytic Shellfish Toxins in the Gastropod Concholepas concholepas: Variability, Toxin Profiles and Mechanisms for Toxicity Reduction" Marine Drugs 21, no. 1: 44. https://doi.org/10.3390/md21010044