
Recent Advancement in Anticancer Compounds from Marine Organisms: Approval, Use and Bioinformatic Approaches to Predict New Targets



Abstract
:1. Introduction
2. The Evolution of Phenotypic Plasticity in the Marine Environment
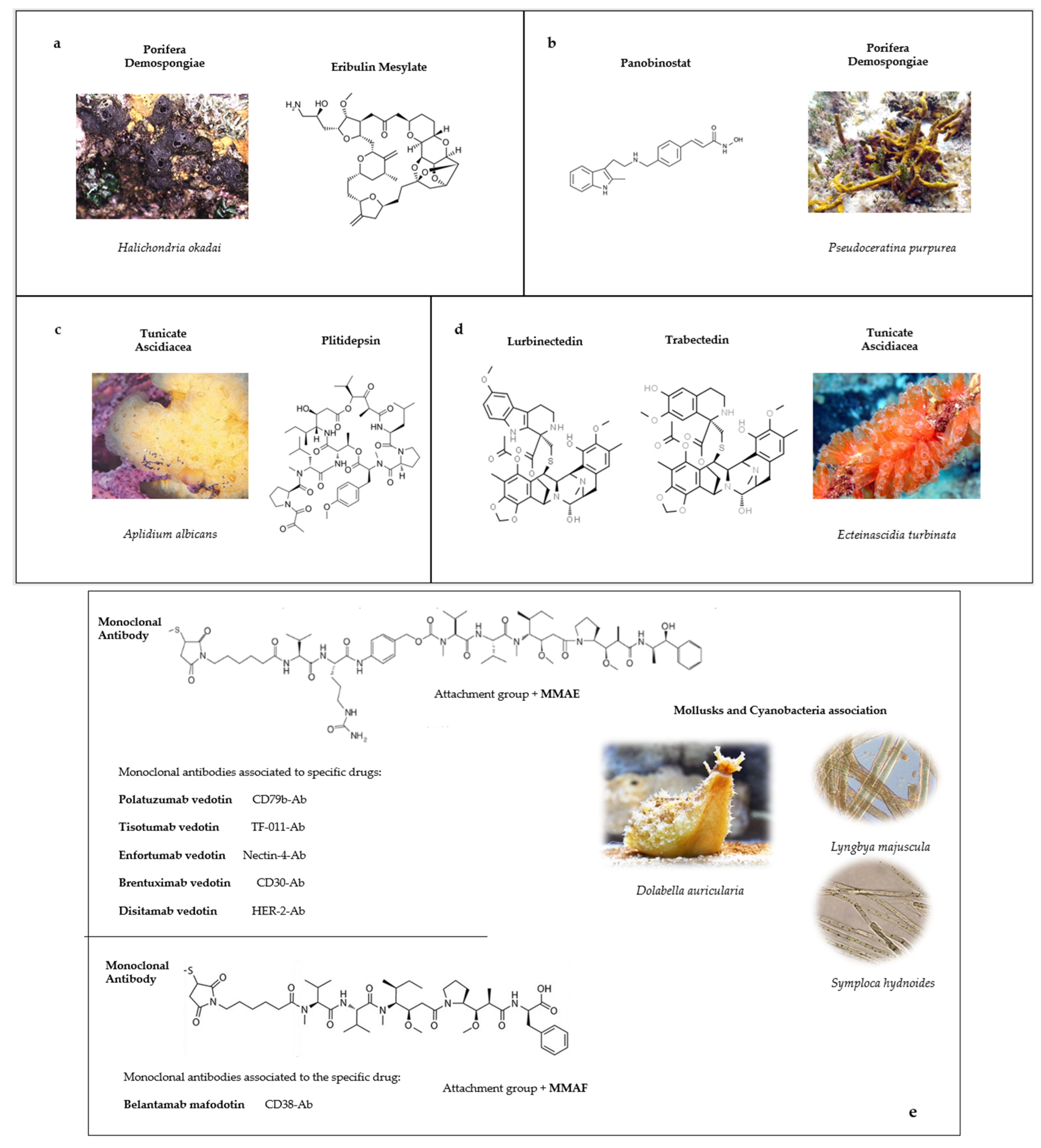
3. Approved Anticancer Compounds from Marine Sponges
3.1. Eribulin Mesylate
3.2. Panobinostat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Chemical Class | Marine Source | Species | Mechanism of Action | References |
|---|---|---|---|---|---|
| Eribulin mesylate | Macrolide | Sponge | Halichondria okadai | Interfering tubulin polymerization | [21,22,24,26,46,47,48] |
| Panobinostat | Hydroxamic acid derivative | Sponge | Pseudoceratina purpurea | HDAC inhibitor | [40,43,45,49,50,51] |
| Aminopeptidase-N inhibitor | [44] | ||||
| DNA Methyltransferase inhibitor | [35] |
4. Approved Anticancer Compounds from Marine Tunicates
4.1. Plitidepsin
4.2. Trabectedin
4.3. Lurbinectedin
| Compound Name | Chemical Class | Marine Source | Species | Mechanism of Action | References |
|---|---|---|---|---|---|
| Lurbinectedin | Alkaloid | Tunicate/bacteria | Ecteinascidia turbinata/ Candidatus Endoecteinascidia frumentensis | Alkylation of DNA | [79] |
| Inhibition of the transcriptional factors binding to DNA | [80,81] | ||||
| Trabectedin | Alkaloid | Tunicate/bacteria | Ecteinascidia turbinata/ Candidatus Endoecteinascidia frumentensis | Block RNA Pol-II | [68,69] |
| Inhibition of the transcriptional factors binding to DNA | [70,71,72] | ||||
| Reduction in TAM | [75,76] | ||||
| Plitidepsin | Peptide | Tunicate | Aplidium albicans | Activation of eEF1A2 | [55,82,83,84] |
5. Approved Anticancer Compounds from Mollusks/Cyanobacteria Association
5.1. Brentuximab Vedotin
5.2. Polatuzumab Vedotin
5.3. Enfortumab Vedotin-Eifv
5.4. Disitamab Vedotin
5.5. Tisotumab Vedotin-tftv
5.6. Belantamab Mafodotin-blmf
| Compound Name | Chemical Class | Marine Source | Species | Mechanism of Action | References |
|---|---|---|---|---|---|
| Brentuximab vedotin | ADC (MMAE + CD30Ab) | Mollusk/ Cyanobacteria | Dolabella auricolaria/ Symploca hynoides, Lyngbya majuscula | Microtubulin targeting agent via CD 30 | [95,96,97] |
| Polatuzumab vedotin | ADC (MMAE + CD-79bAb) | Mollusk/ Cyanobacteria | Dolabella auricolaria/ Symploca hynoides, Lyngbya majuscula | Microtubulin targeting agent via CD79 | [100,101,121] |
| Enfortumab vedotin | ADC (MMAE + Nectin-4 Ab) | Mollusk/ Cyanobacteria | Dolabella auricolaria/ Symploca hynoides, Lyngbya majuscula | Microtubulin targeting agent via Nectin4 | [103,122] |
| Disitamab Vedotin | MMAE + HER-2 Ab | Mollusk/ Cyanobacteria | Dolabella auricolaria/ Symploca hynoides, Lyngbya majuscula | Microtubulin targeting agent via HER-2 | [105,107,108,123] |
| Tisotumab Vedotin | MMAE + TF-011 Ab | Mollusk/ Cyanobacteria | Dolabella auricolaria/ Symploca hynoides, Lyngbya majuscula | Microtubulin targeting agent via TF-011 | [113,114,115,116] |
| Belantamab mafodotin | MMAF + CD38Ab | Mollusk/ Cyanobacteria | Dolabella auricolaria/ Symploca hynoides, Lyngbya majuscula | Microtubulin targeting agent via CD38 | [118,119,120] |
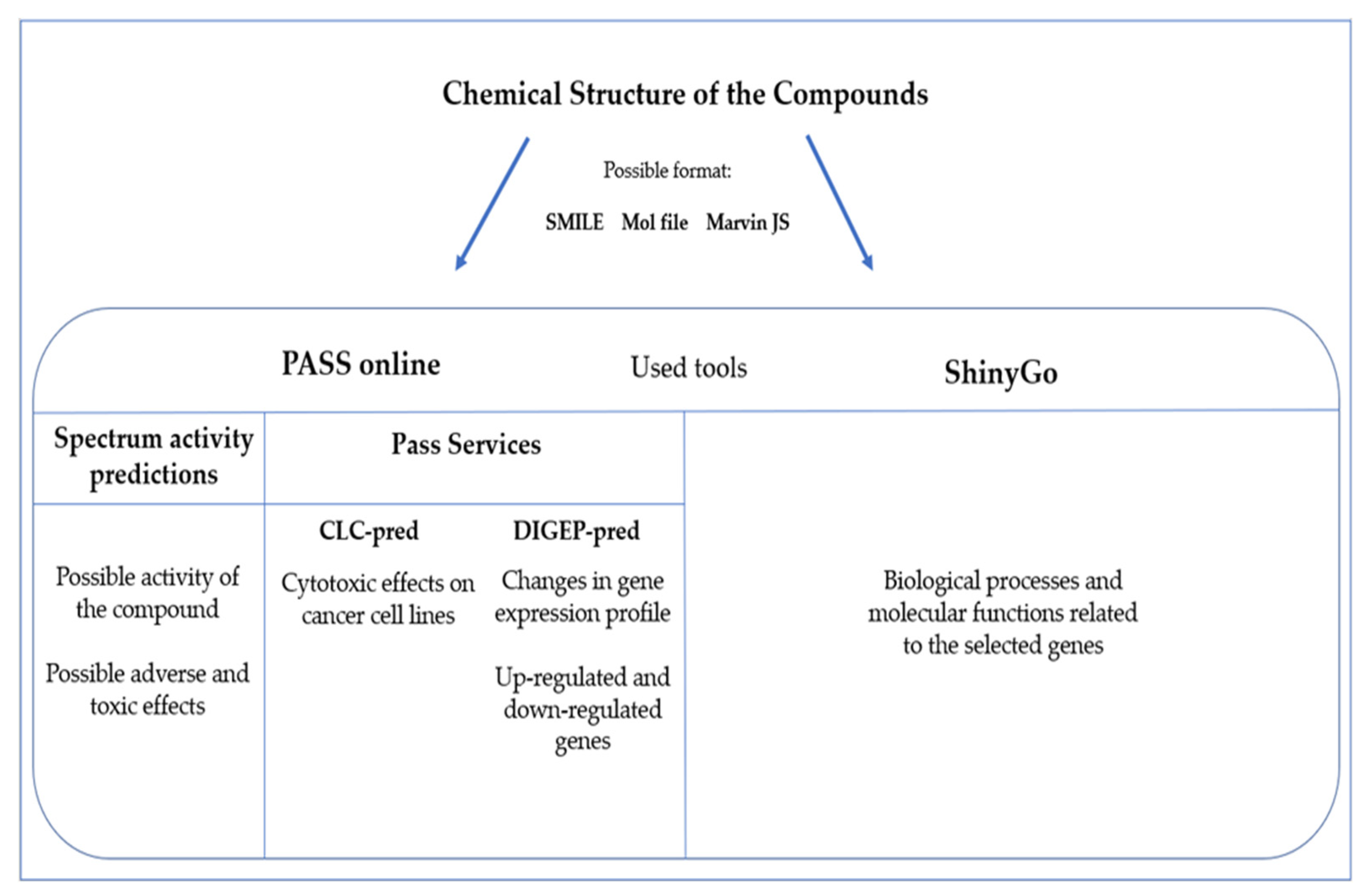
6. Prediction Bioinformatics Tools in Marine-Derived Compounds
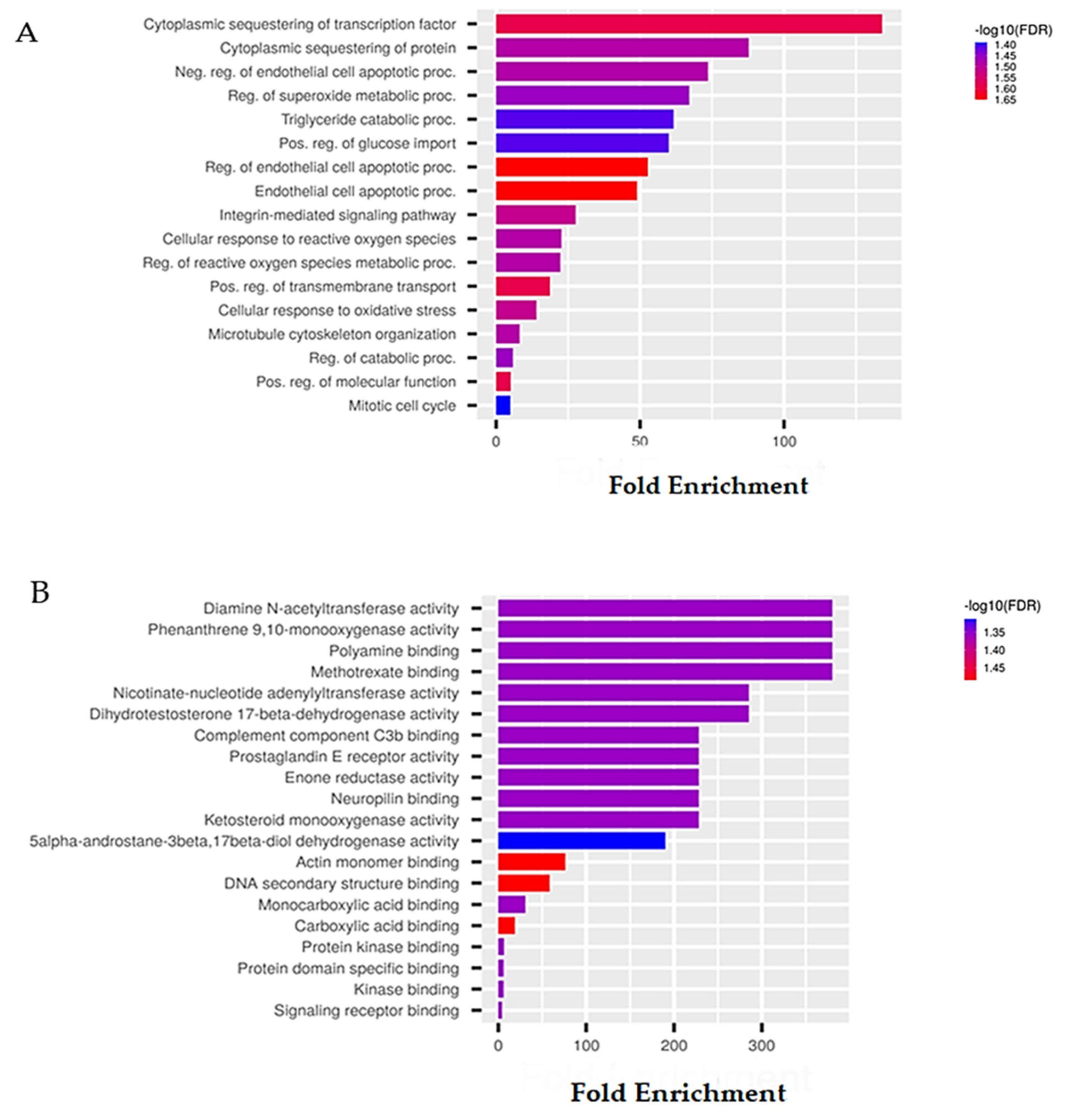
6.1. Sponge-Derived Compound Prediction: The Example of Panobinostat
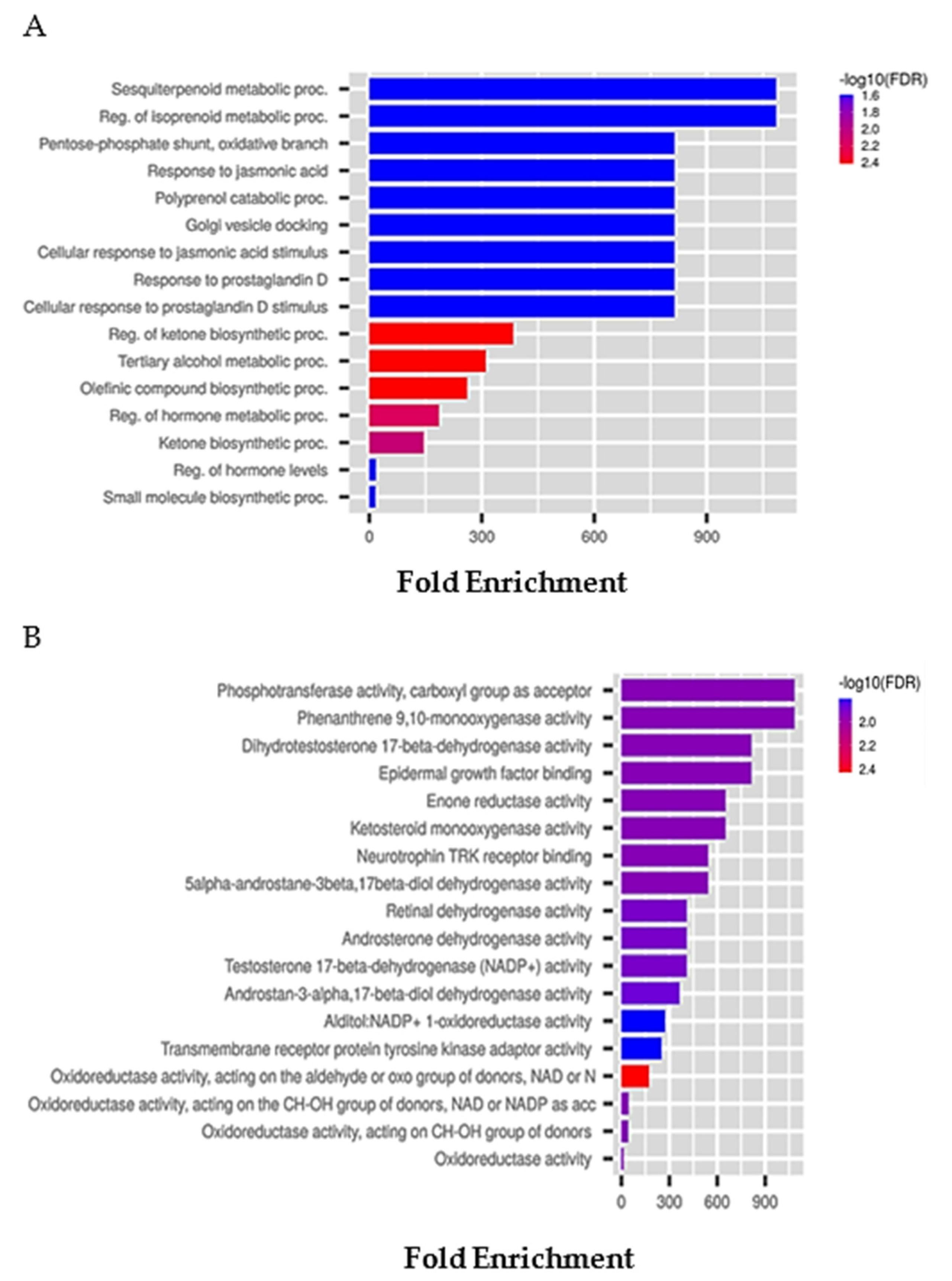
6.2. Tunicate-Derived Compound Prediction: The Example of Plitidepsin
6.3. Mollusks/Cyanobacteria Association-Derived Compound Prediction: The Example of Belantamab Mafodotin
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Wang, Z.; Deisboeck, T.S. Dynamic Targeting in Cancer Treatment. Front. Physiol. 2019, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor Microenvironment Complexity and Therapeutic Implications at a Glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef] [Green Version]
- Bayona, L.M.; de Voogd, N.J.; Choi, Y.H. Metabolomics on the Study of Marine Organisms. Metabolomics 2022, 18, 1–24. [Google Scholar] [CrossRef]
- Lane, A.L.; Moore, B.S. A Sea of Biosynthesis: Marine Natural Products Meet the Molecular Age. Nat. Prod. Rep. 2011, 28, 411–428. [Google Scholar] [CrossRef] [Green Version]
- Barzkar, N.; Jahromi, S.T.; Poorsaheli, H.B.; Vianello, F. Metabolites from Marine Microorganisms, Micro, and Macroalgae: Immense Scope for Pharmacology. Mar. Drugs 2019, 17, 464. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, S.; Abbasi, B.H.; Uzair, B.; Abbasi, R. Natural Products as Reservoirs of Novel Therapeutic Agents. EXCLI J. 2018, 17, 420–451. [Google Scholar] [CrossRef]
- Sigwart, J.D.; Blasiak, R.; Jaspars, M.; Jouffray, J.B.; Tasdemir, D. Unlocking the Potential of Marine Biodiscovery. Nat. Prod. Rep. 2021, 38, 1235–1242. [Google Scholar] [CrossRef]
- Romano, G.; Almeida, M.; Coelho, A.V.; Cutignano, A.; Gonçalves, L.G.; Hansen, E.; Khnykin, D.; Mass, T.; Ramšak, A.; Rocha, M.S.; et al. Biomaterials and Bioactive Natural Products from Marine Invertebrates: From Basic Research to Innovative Applications. Mar. Drugs 2022, 20, 219. [Google Scholar] [CrossRef]
- Gibney, E.R.; Nolan, C.M. Epigenetics and Gene Expression. Heredity 2010, 105, 4–13. [Google Scholar] [CrossRef]
- Duncan, E.J.; Gluckman, P.D.; Dearden, P.K. Epigenetics, Plasticity, and Evolution: How Do We Link Epigenetic Change to Phenotype? J. Exp. Zool. B Mol. Dev. Evol. 2014, 322, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Heard, E.; Martienssen, R.A. Transgenerational Epigenetic Inheritance: Myths and Mechanisms. Cell 2014, 157, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Sommer, R.J. Phenotypic Plasticity: From Theory and Genetics to Current and Future Challenges. Genetics 2020, 215, 1–13. [Google Scholar] [CrossRef]
- World Porifera Database—Species. Available online: https://www.marinespecies.org/porifera/porifera.php?p=stats (accessed on 4 July 2022).
- Amina, M.; Musayeib, N.M. Biological and medicinal importance of sponge. In Biological Resources of Water; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar] [CrossRef] [Green Version]
- Malve, H. Exploring the Ocean for New Drug Developments: Marine Pharmacology. J. Pharm. Bioallied. Sci. 2016, 8, 83. [Google Scholar] [CrossRef]
- Privat De Garilhe, M.; De Rudder, J. Effect of 2 Arabinose Nucleosides on the multiplication of Herpes Virus and vaccine in cell culture. Comptes Rendus Hebd. Seances Acad. Sci. 1964, 259, 2725–2728. [Google Scholar]
- Van Soest, R.W.M.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; de Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.A. Global Diversity of Sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef]
- Hirata, Y.U.D. Halichondrins-Antitumor Polyether Macrolides from a Marine Sponge. Pure Appl. Chem. 1986, 58, 701–710. [Google Scholar]
- Bai, R.; Paull, K.; Herald, C.; Malspeis, L.; Pettit, G.; Hamel, E. Halichondrin B and Homohalichondrin B, Marine Natural Products Binding in the Vinca Domain of Tubulin. Discovery of Tubulin-Based Mechanism of Action by Analysis of differential cytotoxic data. J. Biol. Chem. 1991, 266, 15882–15889. [Google Scholar] [CrossRef]
- Kaufman, P.A.; Awada, A.; Twelves, C.; Yelle, L.; Perez, E.A.; Velikova, G.; Olivo, M.S.; He, Y.; Dutcus, C.E.; Cortes, J. Phase III Open-Label Randomized Study of Eribulin Mesylate Versus Capecitabine in Patients with Locally Advanced or Metastatic Breast Cancer Previously Treated with an Anthracycline and a Taxane. J. Clin. Oncol. 2015, 33, 594. [Google Scholar] [CrossRef]
- Schöffski, P.; Chawla, S.; Maki, R.G.; Italiano, A.; Gelderblom, H.; Choy, E.; Grignani, G.; Camargo, V.; Bauer, S.; Rha, S.Y.; et al. Eribulin versus Dacarbazine in Previously Treated Patients with Advanced Liposarcoma or Leiomyosarcoma: A Randomised, Open-Label, Multicentre, Phase 3 Trial. Lancet 2016, 387, 1629–1637. [Google Scholar] [CrossRef]
- Barni, S.; Livraghi, L.; Morritti, M.; Vici, P.; Michelotti, A.; Cinieri, S.; Fontanella, C.; Porcu, L.; del Mastro, L.; Puglisi, F. Eribulin in the Treatment of Advanced Breast Cancer: Real-World Scenario from 39 Italian Centers—ESEMPiO Study. Future Oncol. 2019, 15, 3247–3258. [Google Scholar] [CrossRef]
- Kuznetsov, G.; Towle, M.; Cheng, H. Induction of Morphological and Biochemical Apoptosis Following Prolonged Mitotic Blockage by Halichondrin B Macrocyclic Ketone Analog E7389. Cancer Res. 2004, 64, 5760–5764. [Google Scholar] [CrossRef] [Green Version]
- Barreca, M.; Spanò, V.; Montalbano, A.; Cueto, M.; Díaz Marrero, A.R.; Deniz, I.; Erdoğan, A.; Bilela, L.L.; Moulin, C.; Taffin-De-Givenchy, E.; et al. Marine Anticancer Agents: An Overview with a Particular Focus on Their Chemical Classes. Mar. Drugs 2020, 18, 619. [Google Scholar] [CrossRef]
- Dybdal-Hargreaves, N.F.; Risinger, A.L.; Mooberry, S.L. Eribulin Mesylate: Mechanism of Action of a Unique Microtubule-Targeting Agent. Clin. Cancer Res. 2015, 21, 2445–2452. [Google Scholar] [CrossRef] [Green Version]
- Quiñoà, E.; Crews, P. Phenolic Constituents of Psammaplysilla. Tetrahedron Lett. 1987, 28, 3229–3232. [Google Scholar] [CrossRef]
- Kim, D.; Lee, I.S.; Jung, J.H.; Yang, S.i. Psammaplin A, a Natural Bromotyrosine Derivative from a Sponge, Possesses the Antibacterial Activity against Methicillin-Resistant Staphylococcus Aureus and the DNA Gyrase-Inhibitory Activity. Arch. Pharm. Res. 1999, 22, 25–29. [Google Scholar] [CrossRef]
- Salam, K.A.; Furuta, A.; Noda, N.; Tsuneda, S.; Sekiguchi, Y.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tsubuki, M.; Tani, H.; et al. Psammaplin A Inhibits Hepatitis C Virus NS3 Helicase. J. Nat. Med. 2013, 67, 765–772. [Google Scholar] [CrossRef]
- Saguez, J.; Dubois, F.; Vincent, C.; Laberche, J.C.; Sangwan-Norreel, B.S.; Giordanengo, P. Differential Aphicidal Effects of Chitinase Inhibitors on the Polyphagous Homopteran Myzus Persicae (Sulzer). Pest Manag. Sci. 2006, 62, 1150–1154. [Google Scholar] [CrossRef]
- Jing, Q.; Hu, X.; Ma, Y.; Mu, J.; Liu, W.; Xu, F.; Li, Z.; Bai, J.; Hua, H.; Li, D. Marine-Derived Natural Lead Compound Disulfide-Linked Dimer Psammaplin A: Biological Activity and Structural Modification. Mar. Drugs 2019, 17, 384. [Google Scholar] [CrossRef] [Green Version]
- Tabudravu, J.N.; Eijsink, V.G.H.; Gooday, G.W.; Jaspars, M.; Komander, D.; Legg, M.; Synstad, B.; van Aalten, D.M.F. Psammaplin A, a Chitinase Inhibitor Isolated from the Fijian Marine Sponge Aplysinella Rhax. Bioorg. Med. Chem. 2002, 10, 1123–1128. [Google Scholar] [CrossRef]
- Kim, D.; Lee, I.S.; Jung, J.H.; Lee, C.O.; Choi, S.U. Psammaplin A, a Natural Phenolic Compound, Has Inhibitory Effect on Human Topoisomerase II and Is Cytotoxic to Cancer Cells. Anticancer Res. 1999, 19, 4085–4090. [Google Scholar] [PubMed]
- Jiménez, C.; Crews, P. Novel Marine Sponge Derived Amino Acids 13. Additional Psammaplin Derivatives from Psammaplysilla Purpurea. Tetrahedron 1991, 47, 2097–2102. [Google Scholar] [CrossRef]
- Piña, I.C.; Gautschi, J.T.; Wang, G.Y.S.; Sanders, M.L.; Schmitz, F.J.; France, D.; Cornell-Kennon, S.; Sambucetti, L.C.; Remiszewski, S.W.; Perez, L.B.; et al. Psammaplins from the Sponge Pseudoceratina purpurea: Inhibition of Both Histone Deacetylase and DNA Methyltransferase. J. Org. Chem. 2003, 68, 3866–3873. [Google Scholar] [CrossRef] [PubMed]
- Dixon, G.; Liao, Y.; Bay, L.K.; Matz, M.v. Role of Gene Body Methylation in Acclimatization and Adaptation in a Basal Metazoan. Proc. Natl. Acad. Sci. USA 2018, 115, 13342–13346. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.D.; Li, J.; Du, L.; Mahdi, F.; Le, T.P.; Chen, W.L.; Swanson, S.M.; Watabe, K.; Nagle, D.G. Biochemical and Anti-Triple Negative Metastatic Breast Tumor Cell Properties of Psammaplins. Mar. Drugs 2018, 16, 442. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.Y.; Jung, J.H.; Na, Y.J.; Kim, H.S. A Natural Histone Deacetylase Inhibitor, Psammaplin A, Induces Cell Cycle Arrest and Apoptosis in Human Endometrial Cancer Cells. Gynecol. Oncol. 2008, 108, 27–33. [Google Scholar] [CrossRef]
- Anne, M.; Sammartino, D.; Barginear, M.F.; Budman, D. Profile of Panobinostat and Its Potential for Treatment in Solid Tumors: An Update. Onco Targets Ther. 2013, 6, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- Wahaib, K.; Beggs, A.E.; Campbell, H.; Kodali, L.; Ford, P.D. Panobinostat: A Histone Deacetylase Inhibitor for the Treatment of Relapsed or Refractory Multiple Myeloma. Am. J. Health Syst. Pharm. 2016, 73, 441–450. [Google Scholar] [CrossRef]
- Qian, D.Z.; Kato, Y.; Shabbeer, S.; Wei, Y.; Verheul, H.M.W.; Salumbides, B.; Sanni, T.; Atadja, P.; Pili, R. Targeting Tumor Angiogenesis with Histone Deacetylase Inhibitors: The Hydroxamic Acid Derivative LBH589. Clin. Cancer Res. 2006, 12, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Bali, P.; Pranpat, M.; Bradner, J.; Balasis, M.; Fiskus, W.; Guo, F.; Rocha, K.; Kumaraswamy, S.; Boyapalle, S.; Atadja, P.; et al. Inhibition of Histone Deacetylase 6 Acetylates and Disrupts the Chaperone Function of Heat Shock Protein 90: A novel basis for antileukemia activity of histone deacetylase inhibitors. J. Biol. Chem. 2005, 280, 26729–26734. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Patel, V.K.; Jain, D.K.; Patel, P.; Rajak, H. Panobinostat as Pan-Deacetylase Inhibitor for the Treatment of Pancreatic Cancer: Recent Progress and Future Prospects. Oncol. Ther. 2016, 4, 73. [Google Scholar] [CrossRef]
- Shim, J.S.; Lee, H.S.; Shin, J.; Kwon, H.J. Psammaplin A, a Marine Natural Product, Inhibits Aminopeptidase N and Suppresses Angiogenesis in Vitro. Cancer Lett. 2004, 203, 163–169. [Google Scholar] [CrossRef]
- San-Miguel, J.F.; Hungria, V.T.M.; Yoon, S.S.; Beksac, M.; Dimopoulos, M.A.; Elghandour, A.; Jedrzejczak, W.W.; Günther, A.; Nakorn, T.N.; Siritanaratkul, N.; et al. Overall Survival of Patients with Relapsed Multiple Myeloma Treated with Panobinostat or Placebo plus Bortezomib and Dexamethasone (the PANORAMA 1 Trial): A Randomised, Placebo-Controlled, Phase 3 Trial. Lancet Haematol. 2016, 3, e506–e515. [Google Scholar] [CrossRef]
- O’Shaughnessy, J.; Kaklamani, V.; Kalinsky, K. Perspectives on the Mechanism of Action and Clinical Application of Eribulin for Metastatic Breast Cancer. Future Oncol. 2019, 15, 1641–1653. [Google Scholar] [CrossRef] [Green Version]
- Tate, C.R.; Rhodes, L.V.; Segar, H.C.; Driver, J.L.; Pounder, F.N.; Burow, M.E.; Collins-Burow, B.M. Targeting Triple-Negative Breast Cancer Cells with the Histone Deacetylase Inhibitor Panobinostat. Breast Cancer Res. 2012, 14, R79. [Google Scholar] [CrossRef] [Green Version]
- Dias, J.; Aguiar, S.; Pereira, D.; André, A.; Gano, L.; Correia, J.; Carrapiço, B.; Rütgen, B.; Malhó, R.; Peleteiro, C.; et al. The Histone Deacetylase Inhibitor Panobinostat Is a Potent Antitumor Agent in Canine Diffuse Large B-Cell Lymphoma. Oncotarget 2018, 9, 28586. [Google Scholar] [CrossRef] [Green Version]
- Shenkar, N.; Swalla, B.J. Global Diversity of Ascidiacea. PLoS ONE 2011, 6, e20657. [Google Scholar] [CrossRef]
- Dou, X.; Dong, B. Origins and Bioactivities of Natural Compounds Derived from Marine Ascidians and Their Symbionts. Mar. Drugs 2019, 17, 670. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, K.; Lithgow-Bertelloni, A.M. Novel Antiviral and Cytotoxic Agent, dehydrodidemnin B. PCT Int. Pat. Appl. 1991, 15, 248086q. [Google Scholar]
- Alonso-Álvarez, S.; Pardal, E.; Sánchez-Nieto, D.; Navarro, M.; Caballero, M.; Mateos, M.; Martín, A. Plitidepsin: Design, Development, and Potential Place in Therapy. Drug Des. Dev. Ther. 2017, 11, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Urdiales, J.; Morata, P.; de Castro, I.; Sánchez-Jiménez, F. Antiproliferative Effect of Dehydrodidemnin B (DDB), a Depsipeptide Isolated from Mediterranean Tunicates. Cancer Lett. 1996, 102, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Losada, A.; Muñoz-Alonso, M.; García, C.; Sánchez-Murcia, P.; Martínez-Leal, J.; Domínguez, J.; Lillo, M.; Gago, F.; Galmarini, C. Translation Elongation Factor EEF1A2 Is a Novel Anticancer Target for the Marine Natural Product Plitidepsin. Sci. Rep. 2016, 6, 35100. [Google Scholar] [CrossRef] [PubMed]
- Leisch, M.; Egle, A.; Greil, R. Plitidepsin: A Potential New Treatment for Relapsed/Refractory Multiple Myeloma. Future Oncol. 2018, 15, 109–120. [Google Scholar] [CrossRef]
- Capalbo, A.; Lauritano, C. Multiple Myeloma: Possible Cure from the Sea. Cancers 2022, 14, 2965. [Google Scholar] [CrossRef] [PubMed]
- Spicka, I.; Ocio, E.; Oakervee, H.; Greil, R.; Banh, R.; Huang, S.; D’Rozario, J.; Dimopoulos, M.; Martínez, S.; Extremera, S.; et al. Randomized Phase III Study (ADMYRE) of Plitidepsin in Combination with Dexamethasone vs. Dexamethasone Alone in Patients with Relapsed/Refractory Multiple Myeloma. Ann. Hematol. 2019, 98, 2139–2150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency. Aplidin. Available online: https://www.ema.europa.eu/en/aplidin (accessed on 23 October 2022).
- Varona, J.; Landete, P.; Lopez-Martin, J.; Estrada, V.; Paredes, R.; Guisado-Vasco, P.; de Orueta, L.; Torralba, M.; Fortun, J.; Vates, R.; et al. Preclinical and Randomized Phase I Studies of Plitidepsin in Adults Hospitalized with COVID-19. Life Sci Alliance 2022, 5, e202101200. [Google Scholar] [CrossRef] [PubMed]
- Papapanou, M.; Papoutsi, E.; Giannakas, T.; Katsaounou, P. Plitidepsin: Mechanisms and Clinical Profile of a Promising Antiviral Agent against COVID-19. J. Pers. Med. 2021, 11, 668. [Google Scholar] [CrossRef]
- Guisado-Vasco, P.; Carralón-González, M.; Aguareles-Gorines, J.; Martí-Ballesteros, E.; Sánchez-Manzano, M.; Carnevali-Ruiz, D.; García-Coca, M.; Barrena-Puertas, R.; de Viedma, R.; Luque-Pinilla, J.; et al. Plitidepsin as a Successful Rescue Treatment for Prolonged Viral SARS-CoV-2 Replication in a Patient with Previous Anti-CD20 Monoclonal Antibody-Mediated B Cell Depletion and Chronic Lymphocytic Leukemia. J. Hematol. Oncol. 2022, 15, 4. [Google Scholar] [CrossRef]
- Carter, N.; Keam, S. Trabectedin. Drugs 2007, 67, 2257–2276. [Google Scholar] [CrossRef]
- Rinehart, K.L. Antitumor Compounds from Tunicates. Med. Res. Rev. 2000, 20, 1–27. [Google Scholar] [CrossRef]
- Larsen, A.K.; Galmarini, C.M.; D’Incalci, M. Unique Features of Trabectedin Mechanism of Action. Cancer Chemother. Pharmacol. 2016, 77, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Aune, G.J.; Takagi, K.; Sordet, O.; Guirouilh-Barbat, J.; Antony, S.; Bohr, V.; Pommier, Y. Von Hippel-Lindau Coupled and Transcription-Coupled Nucleotide Excision Repair Dependent Degradation of RNA Polymerase II in Response to Trabectedin. Clin. Cancer Res. 2008, 14, 6449. [Google Scholar] [CrossRef] [PubMed]
- Feuerhahn, S.; Giraudon, C.; Martínez-Díez, M.; Bueren-Calabuig, J.A.; Galmarini, C.M.; Gago, F.; Egly, J.M. XPF-Dependent DNA Breaks and RNA Polymerase II Arrest Induced by Antitumor DNA Interstrand Crosslinking-Mimetic Alkaloids. Chem. Biol. 2011, 18, 988–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minuzzo, M.; Marchini, S.; Broggini, M.; Faircloth, G.; D’Incalci, M.; Mantovani, R. Interference of Transcriptional Activation by the Antineoplastic Drug Ecteinascidin-743. Proc. Natl. Acad. Sci. USA 2000, 97, 6780–6784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Gorfajn, B.; Faircloth, G.; Scotto, K.W. Ecteinascidin 743, a Transcription-Targeted Chemotherapeutic That Inhibits MDR1 Activation. Proc. Natl. Acad. Sci. USA 2000, 97, 6775–6779. [Google Scholar] [CrossRef] [Green Version]
- Bonfanti, M.; la Valle, E.; Fernandez Sousa Faro, J.M.; Faircloth, G.; Caretti, G.; Mantovani, R.; D’Incalci, M. Effect of Ecteinascidin-743 on the Interaction between DNA Binding Proteins and DNA. Anticancer Drug Des. 1999, 14, 179–186. [Google Scholar]
- Di Giandomenico, S.; Frapolli, R.; Bello, E.; Uboldi, S.; Licandro, S.A.; Marchini, S.; Beltrame, L.; Brich, S.; Mauro, V.; Tamborini, E.; et al. Mode of Action of Trabectedin in Myxoid Liposarcomas. Oncogene 2014, 33, 5201–5210. [Google Scholar] [CrossRef] [Green Version]
- Demetri, G.; von Mehren, M.; Jones, R.; Hensley, M.; Schuetze, S.; Staddon, A.; Milhem, M.; Elias, A.; Ganjoo, K.; Tawbi, H.; et al. Efficacy and Safety of Trabectedin or Dacarbazine for Metastatic Liposarcoma or Leiomyosarcoma After Failure of Conventional Chemotherapy: Results of a Phase III Randomized Multicenter Clinical Trial. J. Clin. Oncol. 2016, 34, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Germano, G.; Frapolli, R.; Belgiovine, C.; Anselmo, A.; Pesce, S.; Liguori, M.; Erba, E.; Uboldi, S.; Zucchetti, M.; Pasqualini, F.; et al. Role of Macrophage Targeting in the Antitumor Activity of Trabectedin. Cancer Cell 2013, 23, 249–262. [Google Scholar] [CrossRef] [Green Version]
- D’Incalci, M.; Badri, N.; Galmarini, C.M.; Allavena, P. Trabectedin, a Drug Acting on Both Cancer Cells and the Tumour Microenvironment. Br. J. Cancer 2014, 111, 646. [Google Scholar] [CrossRef] [Green Version]
- Allavena, P.; Signorelli, M.; Chieppa, M.; Erba, E.; Bianchi, G.; Marchesi, F.; Olimpio, C.; Bonardi, C.; Garbi, A.; Lissoni, A.; et al. Anti-Inflammatory Properties of the Novel Antitumor Agent Yondelis (Trabectedin): Inhibition of Macrophage Differentiation and Cytokine Production. Cancer Res. 2005, 65, 2964–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trigo, J.; Subbiah, V.; Besse, B.; Moreno, V.; López, R.; Sala, M.A.; Peters, S.; Ponce, S.; Fernández, C.; Alfaro, V.; et al. Lurbinectedin as Second-Line Treatment for Patients with Small-Cell Lung Cancer: A Single-Arm, Open-Label, Phase 2 Basket Trial. Lancet Oncol. 2020, 21, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Jaigirdar, A.; Mulkey, F.; Cheng, J.; Hamed, S.; Li, Y.; Liu, J.; Zhao, H.; Goheer, A.; Helms, W.; et al. FDA Approval Summary: Lurbinectedin for the Treatment of Metastatic Small Cell Lung Cancer. Clin. Cancer Res. 2021, 27, 2378–2382. [Google Scholar] [CrossRef] [PubMed]
- Harlow, M.; Maloney, N.; Roland, J.; Guillen Navarro, M.; Easton, M.; Kitchen-Goosen, S.; Boguslawski, E.; Madaj, Z.; Johnson, B.; Bowman, M.; et al. Lurbinectedin Inactivates the Ewing Sarcoma Oncoprotein EWS-FLI1 by Redistributing It within the Nucleus. Cancer Res. 2016, 76, 6657–6668. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.; Jambon, S.; Depauw, S.; David-Cordonnier, M. Targeting Transcription Factors for Cancer Treatment. Molecules 2018, 23, 1479. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Alonso, M.; González-Santiago, L.; Zarich, N.; Martínez, T.; Alvarez, E.; Rojas, J.M.; Muñoz, A. Plitidepsin Has a Dual Effect Inhibiting Cell Cycle and Inducing Apoptosis via Rac1/c-Jun NH2-Terminal Kinase Activation in Human Melanoma Cells. J. Pharmacol. Exp. Ther. 2008, 324, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Erba, E.; Serafini, M.; Gaipa, G.; Tognon, G.; Marchini, S.; Celli, N.; Rotilio, O.; Broggini, M.; Jimeno, J.; Faircloth, G.; et al. Effect of Aplidin in Acute Lymphoblastic Leukaemia Cells. Br. J. Cancer 2003, 89, 763–773. [Google Scholar] [CrossRef]
- Cuadrado, A.; González, L.; Suárez, Y.; Martínez, T.; Muñoz, A. JNK Activation Is Critical for AplidinTM-Induced Apoptosis. Oncogene 2004, 23, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Guiry, M. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Molluscabase. Available online: https://www.molluscabase.org/ (accessed on 5 July 2022).
- Hrouzek, P. Secondary metabolites produced by cyanobacteria in symbiotic associations. In Algal and Cyanobacteria Symbioses; World Scientific: Singapore, 2017; pp. 611–626. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tuinman, A.A.; Boettner, F.E.; Kizu, H.; Schmidt, J.M.; Baczynskyj, L.; Tomer, K.B.; Bontems, R.J. The Isolation and Structure of a Remarkable Marine Animal Antineoplastic Constituent: Dolastatin 10. J. Am. Chem. Soc. 1987, 109, 6883–6885. [Google Scholar] [CrossRef]
- Pettit, G.; Kamano, Y.; Fujii, Y.; Herald, C.; Inoue, M.; Brown, P.; Gust, D.; Kitahara, K.; Schmidt, J.; Doubek, D.; et al. Marine Animal Biosynthetic Constituents for Cancer Chemotherapy. J. Nat. Prod. 1981, 44, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, G.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H.; Valeriote, F.A. Symplostatin 1: A Dolastatin 10 Analogue from the Marine Cyanobacterium Symploca Hydnoides. J. Nat. Prod. 1998, 61, 1075–1077. [Google Scholar] [CrossRef] [PubMed]
- Robles-Bañuelos, B.; Durán-Riveroll, L.; Rangel-López, E.; Pérez-López, H.; González-Maya, L. Marine Cyanobacteria as Sources of Lead Anticancer Compounds: A Review of Families of Metabolites with Cytotoxic, Antiproliferative, and Antineoplastic Effects. Molecules 2022, 27, 4814. [Google Scholar] [CrossRef] [PubMed]
- Maderna, A.; Leverett, C. Recent Advances in the Development of New Auristatins: Structural Modifications and Application in Antibody Drug Conjugates. Mol. Pharm. 2015, 12, 1798–1812. [Google Scholar] [CrossRef]
- Maderna, A.; Doroski, M.; Subramanyam, C.; Porte, A.; Leverett, C.; Vetelino, B.; Chen, Z.; Risley, H.; Parris, K.; Pandit, J.; et al. Discovery of Cytotoxic Dolastatin 10 Analogues with N-Terminal Modifications. J. Med. Chem. 2014, 57, 10527–10543. [Google Scholar] [CrossRef]
- Akaiwa, M.; Dugal-Tessier, J.; Mendelsohn, B. Antibody-Drug Conjugate Payloads; Study of Auristatin Derivatives. Chem. Pharm. Bull. 2020, 68, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Liu, D. Antibody-Drug Conjugates in Clinical Trials for Lymphoid Malignancies and Multiple Myeloma. J. Hematol. Oncol. 2019, 12, 94. [Google Scholar] [CrossRef]
- Van de Donk, N.W.C.J.; Dhimolea, E. Brentuximab Vedotin. MAbs 2012, 4, 458. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, R. Immune compromise associated with biologics. In Stiehm’s Immune Deficiencies; Academic Press: Cambridge, MA, USA, 2014; pp. 889–906. [Google Scholar] [CrossRef]
- Jiang, J.; Li, S.; Shan, X.; Wang, L.; Ma, J.; Huang, M.; Dong, L.; Chen, F. Preclinical Safety Profile of Disitamab Vedotin: A Novel Anti-HER2 Antibody Conjugated with MMAE. Toxicol. Lett. 2020, 324, 30–37. [Google Scholar] [CrossRef]
- Young, R.; Shaffer, A.; Phelan, J.; Staudt, L. B-Cell Receptor Signaling in Diffuse Large B-Cell Lymphoma. Semin. Hematol. 2015, 52, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Diefenbach, C. Polatuzumab Vedotin: A New Target for B Cell Malignancies. Curr. Hematol. Malig. Rep. 2020, 15, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Tilly, H.; Morschhauser, F.; Bartlett, N.L.; Mehta, A.; Salles, G.; Haioun, C.; Munoz, J.; Chen, A.I.; Kolibaba, K.; Lu, D.; et al. Polatuzumab Vedotin in Combination with Immunochemotherapy in Patients with Previously Untreated Diffuse Large B-Cell Lymphoma: An Open-Label, Non-Randomised, Phase 1b–2 Study. Lancet Oncol. 2019, 20, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Challita-Eid, P.; Satpayev, D.; Yang, P.; An, Z.; Morrison, K.; Shostak, Y.; Raitano, A.; Nadell, R.; Liu, W.; Lortie, D.; et al. Enfortumab Vedotin Antibody-Drug Conjugate Targeting Nectin-4 Is a Highly Potent Therapeutic Agent in Multiple Preclinical Cancer Models. Cancer Res. 2016, 76, 3003–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powles, T.; Rosenberg, J.; Sonpavde, G.; Loriot, Y.; Durán, I.; Lee, J.; Matsubara, N.; Vulsteke, C.; Castellano, D.; Wu, C.; et al. Enfortumab Vedotin in Previously Treated Advanced Urothelial Carcinoma. N. Engl. J. Med. 2021, 384, 1125–1135. [Google Scholar] [CrossRef]
- Shi, F.; Liu, Y.; Zhou, X.; Shen, P.; Xue, R.; Zhang, M. Disitamab Vedotin: A Novel Antibody-Drug Conjugates for Cancer Therapy. Drug Deliv. 2022, 29, 1335. [Google Scholar] [CrossRef]
- Deeks, E.D. Disitamab Vedotin: First Approval. Drugs 2021, 81, 1929–1935. [Google Scholar] [CrossRef]
- ClinicalTrials.Gov. A Study of RC48-ADC in Subjects with Advanced Breast Cancer—Full Text View. Available online: https://clinicaltrials.gov/ct2/show/NCT03052634 (accessed on 22 June 2022).
- Patelli, G.; Zeppellini, A.; Spina, F.; Righetti, E.; Stabile, S.; Amatu, A.; Tosi, F.; Ghezzi, S.; Siena, S.; Sartore-Bianchi, A. The Evolving Panorama of HER2-Targeted Treatments in Metastatic Urothelial Cancer: A Systematic Review and Future Perspectives. Cancer Treat. Rev. 2022, 104, 102351. [Google Scholar] [CrossRef]
- Sheng, X.; Yan, X.; Wang, L.; Shi, Y.; Yao, X.; Luo, H.; Shi, B.; Liu, J.; He, Z.; Yu, G.; et al. Open-Label, Multicenter, Phase II Study of RC48-ADC, a HER2-Targeting Antibody-Drug Conjugate, in Patients with Locally Advanced or Metastatic Urothelial Carcinoma. Clin. Cancer Res. 2021, 27, 43–51. [Google Scholar] [CrossRef]
- Zhao, X.; Cheng, C.; Gou, J.; Yi, T.; Qian, Y.; Du, X.; Zhao, X. Expression of Tissue Factor in Human Cervical Carcinoma Tissue. Exp. Ther. Med. 2018, 16, 4075–4081. [Google Scholar] [CrossRef] [Green Version]
- Cocco, E.; Varughese, J.; Buza, N.; Bellone, S.; Lin, K.Y.; Bellone, M.; Todeschini, P.; Silasi, D.A.; Azodi, M.; Schwartz, P.E.; et al. Tissue Factor Expression in Ovarian Cancer: Implications for Immunotherapy with HI-Con1, a Factor VII-IgGF(c) Chimeric Protein Targeting Tissue Factor. Clin. Exp. Metastasis 2011, 28, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Patry, G.; Hovington, H.; Larue, H.; Harel, F.; Fradet, Y.; Lacombe, L. Tissue Factor Expression Correlates with Disease-Specific Survival in Patients with Node-Negative Muscle-Invasive Bladder Cancer. Int. J. Cancer 2008, 122, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- de Bono, J.; Concin, N.; Hong, D.; Thistlethwaite, F.; Machiels, J.; Arkenau, H.; Plummer, R.; Jones, R.; Nielsen, D.; Windfeld, K.; et al. Tisotumab Vedotin in Patients with Advanced or Metastatic Solid Tumours (InnovaTV 201): A First-in-Human, Multicentre, Phase 1–2 Trial. Lancet Oncol. 2019, 20, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, Y.W.; Osanto, S.; Reitsma, P.H.; Versteeg, H.H. The Relationship between Tissue Factor and Cancer Progression: Insights from Bench and Bedside. Blood 2012, 119, 924–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasthuri, R.S.; Taubman, M.B.; Mackman, N. Role of Tissue Factor in Cancer. J. Clin. Oncol. 2009, 27, 4834–4838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, U.; Schlegel, P.; Lang, P. Natural Killer Cell Mediated Antibody-Dependent Cellular Cytotoxicity in Tumor Immunotherapy with Therapeutic Antibodies. Front. Immunol. 2013, 4, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, R.; Lorusso, D.; Gennigens, C.; González-Martín, A.; Randall, L.; Cibula, D.; Lund, B.; Woelber, L.; Pignata, S.; Forget, F.; et al. Efficacy and Safety of Tisotumab Vedotin in Previously Treated Recurrent or Metastatic Cervical Cancer (InnovaTV 204/GOG-3023/ENGOT-Cx6): A Multicentre, Open-Label, Single-Arm, Phase 2 Study. Lancet Oncol. 2021, 22, 609–619. [Google Scholar] [CrossRef]
- FDA. FDA Grants Accelerated Approval to Tisotumab Vedotin-Tftv for Recurrent or Metastatic Cervical Cancer. Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-grants-accelerated-approval-tisotumab-vedotin-tftv-recurrent-or-metastatic-cervical-cancer (accessed on 23 June 2022).
- Offidani, M.; Corvatta, L.; Morè, S.; Olivieri, A. Belantamab Mafodotin for the Treatment of Multiple Myeloma: An Overview of the Clinical Efficacy and Safety. Drug Des. Devel. Ther. 2021, 15, 2401–2415. [Google Scholar] [CrossRef]
- Condorelli, A.; Garibaldi, B.; Gagliano, C.; Romano, A.; del Fabro, V.; Parrinello, N.L.; Longo, A.; Cosentino, S.; di Raimondo, F.; Conticello, C. Belantamab Mafodotin and Relapsed/Refractory Multiple Myeloma: This Is Not Game Over. Acta Haematol. 2021, abstract. [Google Scholar] [CrossRef]
- Markham, A. Belantamab Mafodotin: First Approval. Drugs 2020, 80, 1607–1613. [Google Scholar] [CrossRef]
- Li, D.; Lee, D.; Dere, R.C.; Zheng, B.; Yu, S.F.; Fuh, F.K.; Kozak, K.R.; Chung, S.; Bumbaca Yadav, D.; Nazzal, D.; et al. Evaluation and Use of an Anti-cynomolgus Monkey CD79b Surrogate Antibody–Drug Conjugate to Enable Clinical Development of Polatuzumab Vedotin. Br. J. Pharmacol. 2019, 176, 3805. [Google Scholar] [CrossRef] [Green Version]
- Lagunin, A.; Dubovskaja, V.; Rudik, A.; Pogodin, P.; Druzhilovskiy, D.; Gloriozova, T.; Filimonov, D.; Sastry, N.; Poroikov, V. CLC-Pred: A Freely Available Web-Service for in Silico Prediction of Human Cell Line Cytotoxicity for Drug-like Compounds. PLoS ONE 2018, 13, e0191838. [Google Scholar] [CrossRef] [PubMed]
- Lagunin, A.; Stepanchikova, A.; Filimonov, D.; Poroikov, V. PASS: Prediction of Activity Spectra for Biologically Active Substances. Bioinformatics 2000, 16, 747–748. [Google Scholar] [CrossRef] [Green Version]
- Lagunin, A.; Ivanov, S.; Rudik, A.; Filimonov, D.; Poroikov, V. DIGEP-Pred: Web Service for in Silico Prediction of Drug-Induced Gene Expression Profiles Based on Structural Formula. Bioinformatics 2013, 29, 2062–2063. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Jung, D.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Bilbao, M.; Katz, C.; Kass, S.; Smith, D.; Hunter, K.; Warshal, D.; Aikins, J.; Ostrovsky, O. Epigenetic Therapy Augments Classic Chemotherapy in Suppressing the Growth of 3D High-Grade Serous Ovarian Cancer Spheroids over an Extended Period of Time. Biomolecules 2021, 11, 1711. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Cheong, H.; Kim, S.; Yoon, J.; Kim, H.; Kim, K.; Kim, S.; Kim, H.; Bae, S.; Kim, C. Low-Dose Combinations of LBH589 and TRAIL Can Overcome TRAIL-Resistance in Colon Cancer Cell Lines. Anticancer. Res. 2011, 31, 3385–3394. [Google Scholar] [PubMed]
- Huang, H.y.; Wang, Y.; Wang, W.d.; Wei, X.l.; Gale, R.P.; Li, J.y.; Zhang, Q.y.; Shu, L.l.; Li, L.; Li, J.; et al. A Prognostic Survival Model Based on Metabolism-Related Gene Expression in Plasma Cell Myeloma. Leukemia 2021, 35, 3212–3222. [Google Scholar] [CrossRef]
- Fedeli, E.; Lancelot, A.; Dominguez, J.; Serrano, J.; Calvo, P.; Sierra, T. Self-Assembling Hybrid Linear-Dendritic Block Copolymers: The Design of Nano-Carriers for Lipophilic Antitumoral Drugs. Nanomaterials 2019, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, H.H.; Schneider, W.M.; Rozen-Gagnon, K.; Miles, L.A.; Schuster, F.; Razooky, B.; Jacobson, E.; Wu, X.; Yi, S.; Rudin, C.M.; et al. TMEM41B Is a Pan-Flavivirus Host Factor. Cell 2021, 184, 133–148.e20. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, Y.; Gu, Z.; Wang, H.; Xia, J.; Wu, X.; Zhan, X.; Levasseur, D.; Zhou, Y.; Janz, S.; et al. ALDH1 Activity Identifies Tumor-Initiating Cells and Links to Chromosomal Instability Signatures in Multiple Myeloma. Leukemia 2014, 28, 1155–1158. [Google Scholar] [CrossRef]
- Conte, M.; Fontana, E.; Nebbioso, A.; Altucci, L. Marine-Derived Secondary Metabolites as Promising Epigenetic Bio-Compounds for Anticancer Therapy. Mar. Drugs 2020, 19, 15. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ye, K.; Jiang, S.; Zhou, G. Marine Power on Cancer: Drugs, Lead Compounds, and Mechanisms. Mar. Drugs 2021, 19, 488. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xue, Y.; Wang, X.; Smith, L.S.; He, B.; Liu, S.; Zhu, H.J. Tissue- and Cell-Expression of Druggable Host Proteins Provide Insights into Repurposing Drugs for COVID-19. Clin. Transl. Sci. 2022, 15, 2796–2811. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M. Plitidepsin: A Repurposed Drug for the Treatment of COVID-19. Antimicrob. Agents Chemother. 2021, 65, e00200-21. [Google Scholar] [CrossRef]





| Activity | Pa | Pi |
|---|---|---|
| HDAC 1 inhibitor | 0.843 | 0.001 |
| HDAC class I inhibitor | 0.842 | 0.001 |
| HDAC inhibitor | 0.792 | 0.002 |
| HDAC 2 inhibitor | 0.760 | 0.001 |
| HDAC 4 inhibitor | 0.706 | 0.001 |
| HDAC IIa inhibitor | 0.700 | 0.001 |
| HDAC 8 inhibitor | 0.530 | 0.001 |
| Chemosensitizer | 0.523 | 0.016 |
| Calcium channel (voltage-sensitive) activator | 0.521 | 0.064 |
| Possible Adverse and Toxic Effects | Pa | Pi |
|---|---|---|
| Ulcer, gastric | 0.362 | 0.057 |
| Occult bleeding Ulcer, peptic | 0.421 0.346 | 0.131 0.088 |
| Cell Line Full Name and Code | Tissue | Pa | Pi |
|---|---|---|---|
| Colon Carcinoma HCT-116 | Colon | 0.784 | 0.0070 |
| Non-small cell lung carcinoma NCI-H1299 | Lung | 0.543 | 0.004 |
| Down-Regulated Genes | Pa | Pi | Up-Regulated Genes | Pa | Pi |
|---|---|---|---|---|---|
| CTPS1 INCENP KEAP1 AKR1C3 EIF4G2 RCC2 ABL1 PHF11 TOB1 DHFR FABP4 MAPK4 NPM | 0.875 0.863 0.795 0.802 0.700 0.697 0.676 0.683 0.715 0.701 0.664 0.563 0.568 | 0.001 0.006 0.008 0.022 0.003 0.002 0.002 0.013 0.050 0.048 0.049 0.114 0.137 | TACC1 ITGAM H1F0 HPGD FGF21 TMSB4X OCLN SAT1 | 0.833 0.819 0.759 0.748 0.686 0.659 0.573 0.592 | 0.004 0.021 0.005 0.032 0.01 0.034 0.039 0.129 |
| Activity | Pa | Pi |
|---|---|---|
| Immunosuppressant | 0.788 | 0.006 |
| Antibiotic glycopeptide-like | 0.738 | 0.003 |
| General pump inhibitor | 0.649 | 0.014 |
| Antineoplastic | 0.657 | 0.034 |
| CYP2H substrate | 0.650 | 0.050 |
| Antifungal | 0.585 | 0.020 |
| Antineoplastic (colorectal cancer) | 0.548 | 0.010 |
| Antineoplastic (colon cancer) | 0.541 | 0.010 |
| Xenobiotic-transporting ATPase inhibitor | 0.524 | 0.009 |
| Antibacterial | 0.453 | 0.021 |
| Possible Adverse and Toxic Effects | Pa | Pi |
|---|---|---|
| Dyskinesia Sleep disturbance Dyspnea Ataxia | 0.960 0.898 0.892 0.776 | 0.004 0.012 0.006 0.013 |
| Cell Line Full Name and Code | Tissue | Pa | Pi |
|---|---|---|---|
| Lung carcinoma A549 Colon adenocarcinoma HT-29 Breast adenocarcinoma MDA-MB-231 Lung carcinoma DMS-114 | Lung Colon Breast Lung | 0.808 0.801 0.554 0.501 | 0.011 0.005 0.020 0.038 |
| Down-Regulated Genes | Pa | Pi | Up-Regulated Genes | Pa | Pi |
|---|---|---|---|---|---|
| NSF ALDH18A1 H6PD SLC15A1 MYBL1 BACE1 SLC14A1 TOB1 VTN BARD1 FKBP5 CTPS1 HSPB11 TAGLN | 0.907 0.879 0.782 0.765 0.739 0.697 0.675 0.682 0.605 0.636 0.599 0.566 0.501 0.516 | 0.012 0.015 0.019 0.036 0.040 0.016 0.053 0.063 0.036 0.120 0.086 0.090 0.053 0.088 | TMEM41B C10ORF118 FAM49A PLXNA2 HMGCR PSAP TXNDC9 PLK3 FGF21 C8ORF4 WIPI1 GPRC5A NUCB2 | 0.823 0.826 0.756 0.751 0.637 0.635 0.542 0.595 0.517 0.552 0.530 0.507 0.523 | 0.009 0.012 0.041 0.047 0.035 0.055 0.025 0.092 0.034 0.083 0.106 0.087 0.105 |
| Activity | Pa | Pi |
|---|---|---|
| Immunostimulant | 0.823 | 0.008 |
| Proteasome ATPase inhibitor | 0.774 | 0.007 |
| Antineoplastic (non-Hodgkin’s lymphoma) | 0.729 | 0.003 |
| Muramoyltetrapeptide carboxypeptidase inhibitor | 0.636 | 0.021 |
| Antineoplastic (solid tumors) | 0.537 | 0.010 |
| Peptide agonist | 0.537 | 0.039 |
| Neuropeptide Y4 antagonist | 0.461 | 0.027 |
| Antineoplastic (pancreatic cancer) | 0.437 | 0.010 |
| CYP2H substrate | 0.502 | 0.122 |
| Fibroblast growth factor agonist | 0.419 | 0.056 |
| Cell Line Full Name and Code | Tissue | Pa | Pi |
|---|---|---|---|
| Breast adenocarcinoma MDA-MB-231 Breast carcinoma MCF7 | Breast Breast | 0.501 0.513 | 0.028 0.049 |
| Down-Regulated Genes | Pa | Pi | Up-Regulated Genes | Pa | Pi |
|---|---|---|---|---|---|
| ALDH18A1 SHC1 NSF H6PD AKR1C3 | 0.698 0.550 0.546 0.524 0.511 | 0.098 0.040 0.117 0.127 0.211 | C10ORF118 TMEM41B | 0.631 0.606 | 0.063 0.051 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santaniello, G.; Nebbioso, A.; Altucci, L.; Conte, M. Recent Advancement in Anticancer Compounds from Marine Organisms: Approval, Use and Bioinformatic Approaches to Predict New Targets. Mar. Drugs 2023, 21, 24. https://doi.org/10.3390/md21010024
Santaniello G, Nebbioso A, Altucci L, Conte M. Recent Advancement in Anticancer Compounds from Marine Organisms: Approval, Use and Bioinformatic Approaches to Predict New Targets. Marine Drugs. 2023; 21(1):24. https://doi.org/10.3390/md21010024
Chicago/Turabian StyleSantaniello, Giovanna, Angela Nebbioso, Lucia Altucci, and Mariarosaria Conte. 2023. "Recent Advancement in Anticancer Compounds from Marine Organisms: Approval, Use and Bioinformatic Approaches to Predict New Targets" Marine Drugs 21, no. 1: 24. https://doi.org/10.3390/md21010024