
Novel β-Hairpin Peptide from Marine Polychaeta with a High Efficacy against Gram-Negative Pathogens

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Novel β-Hairpin AMPs
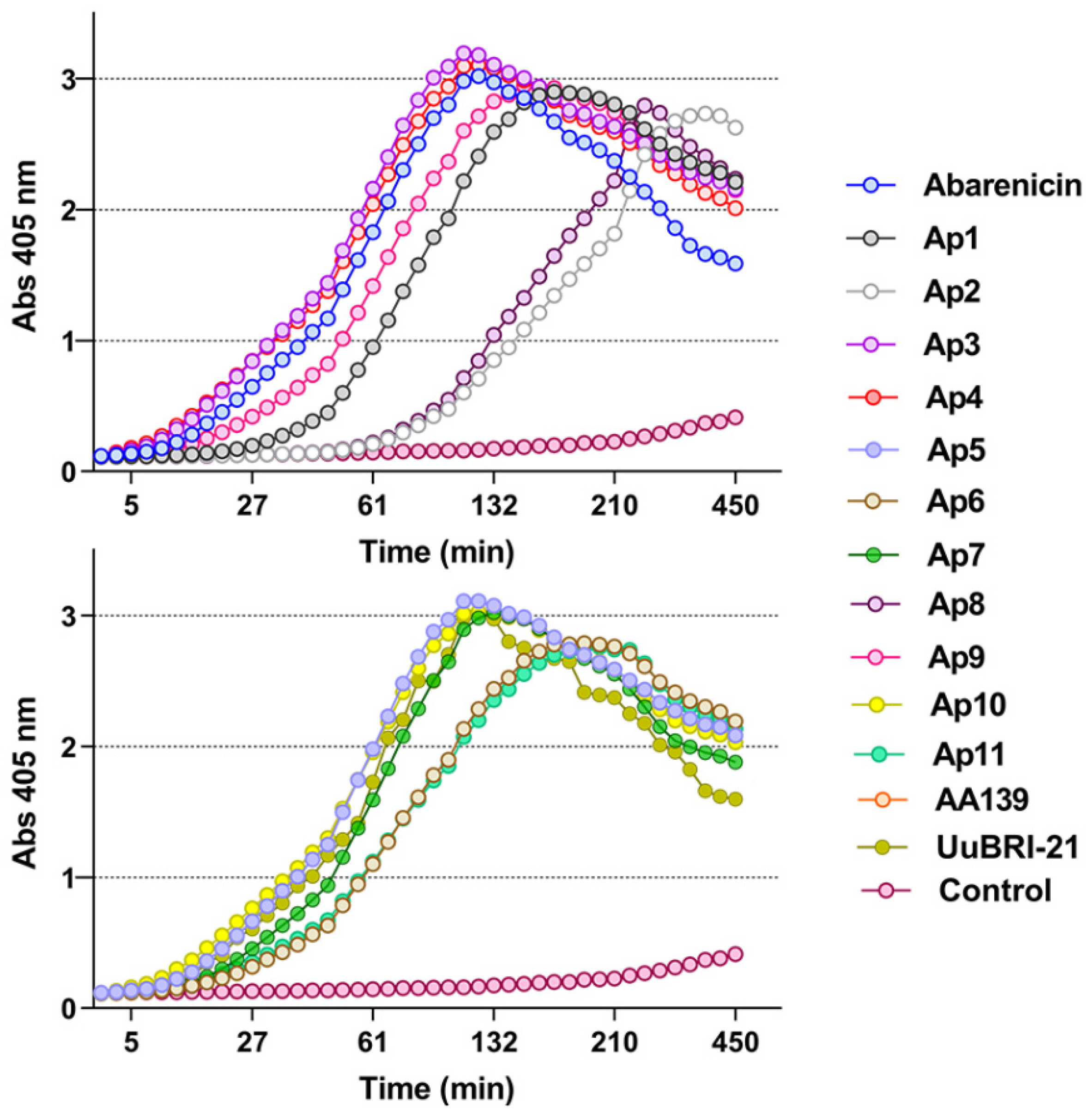
2.2. Recombinant Expression and Biological Activity of the Novel β-Hairpin AMPs
2.3. Abarenicin Analogs Display a Potent Antibacterial Activity against Gram-Negative ESKAPE Pathogens
2.4. The Analogs of Abarenicin Selectively Target Bacterial Membranes
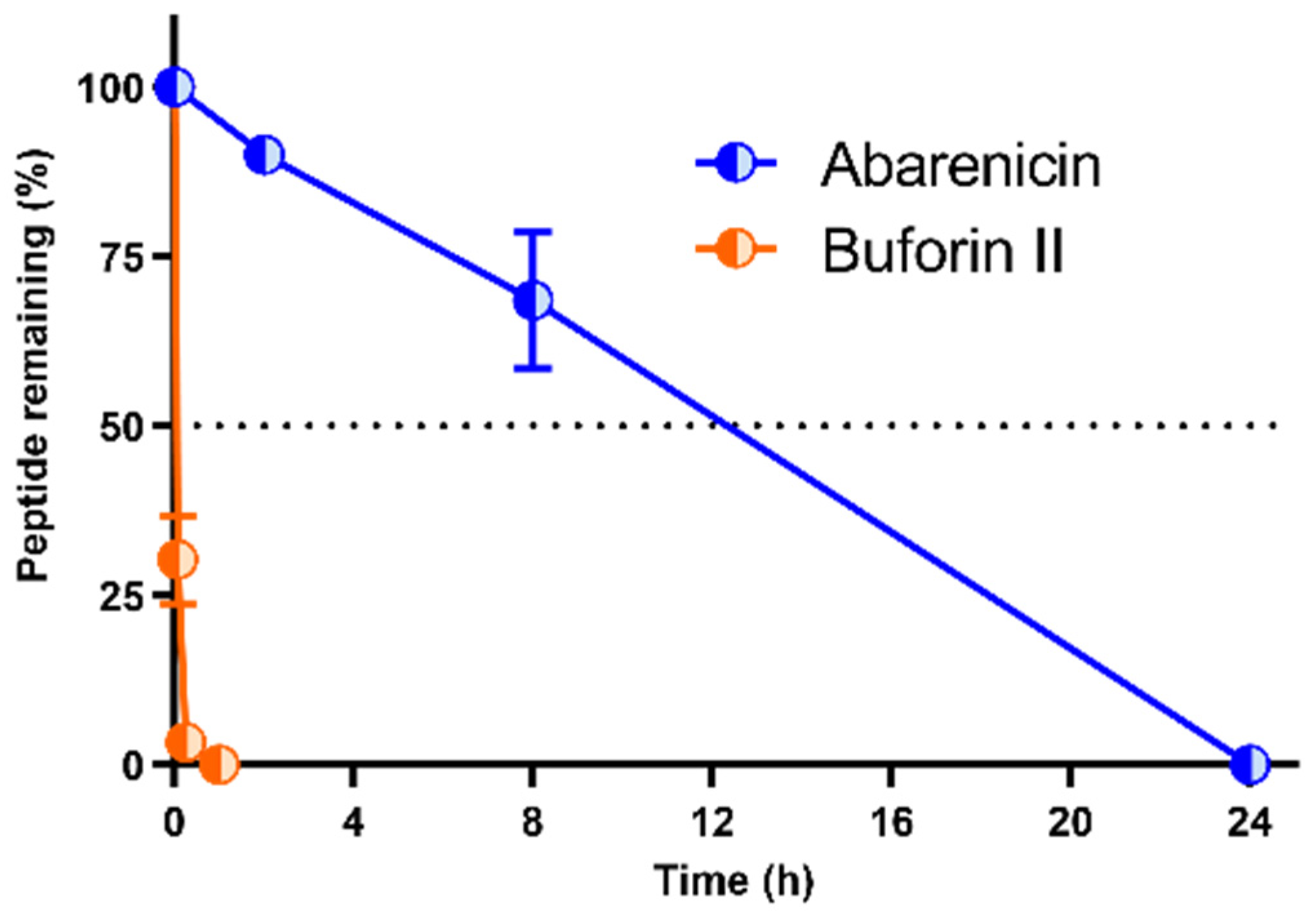
2.5. Stability of Abarenicin to Degradation by Serum Proteases
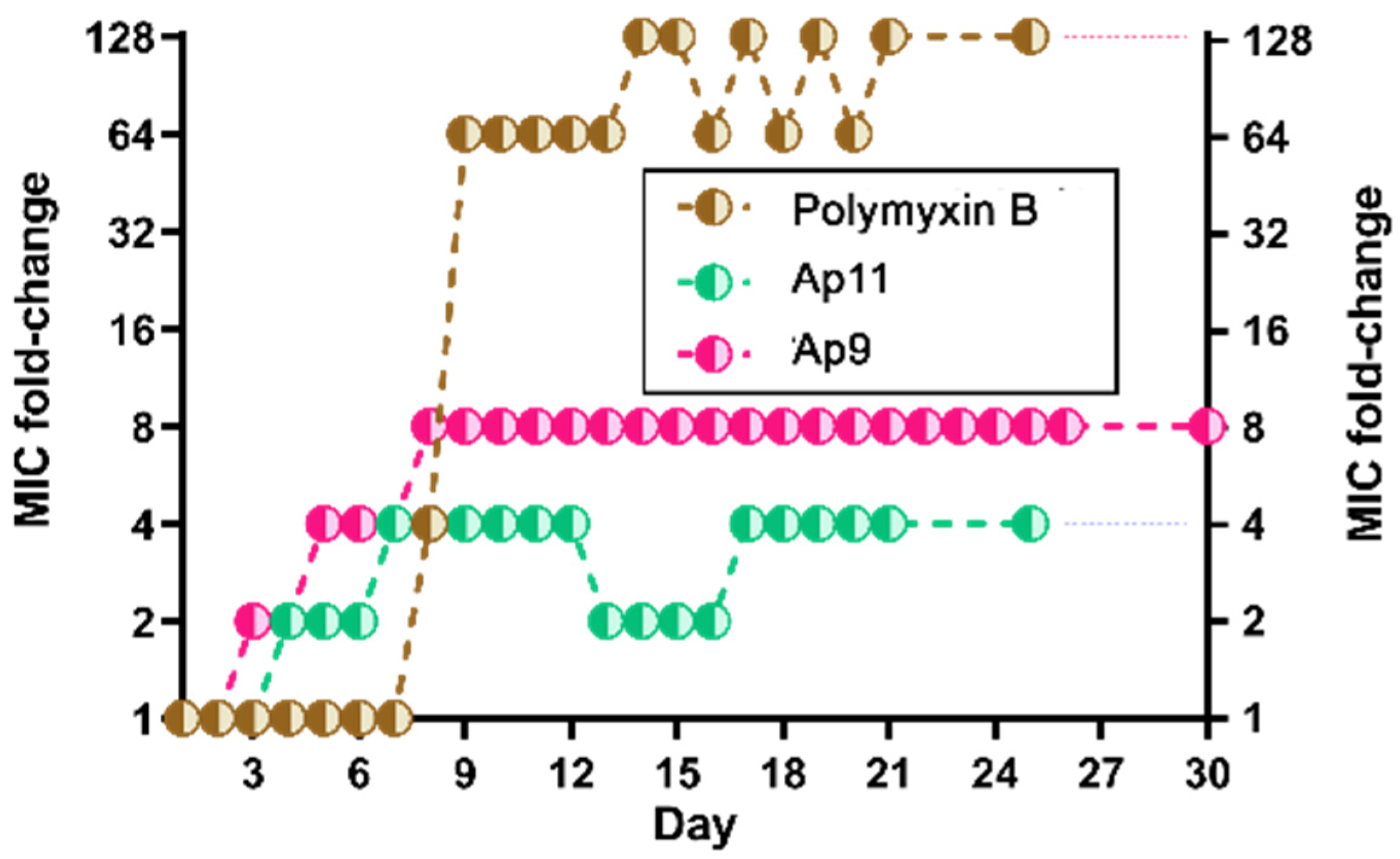
2.6. Abarenicins Do Not Induce a Strong Bacterial Resistance
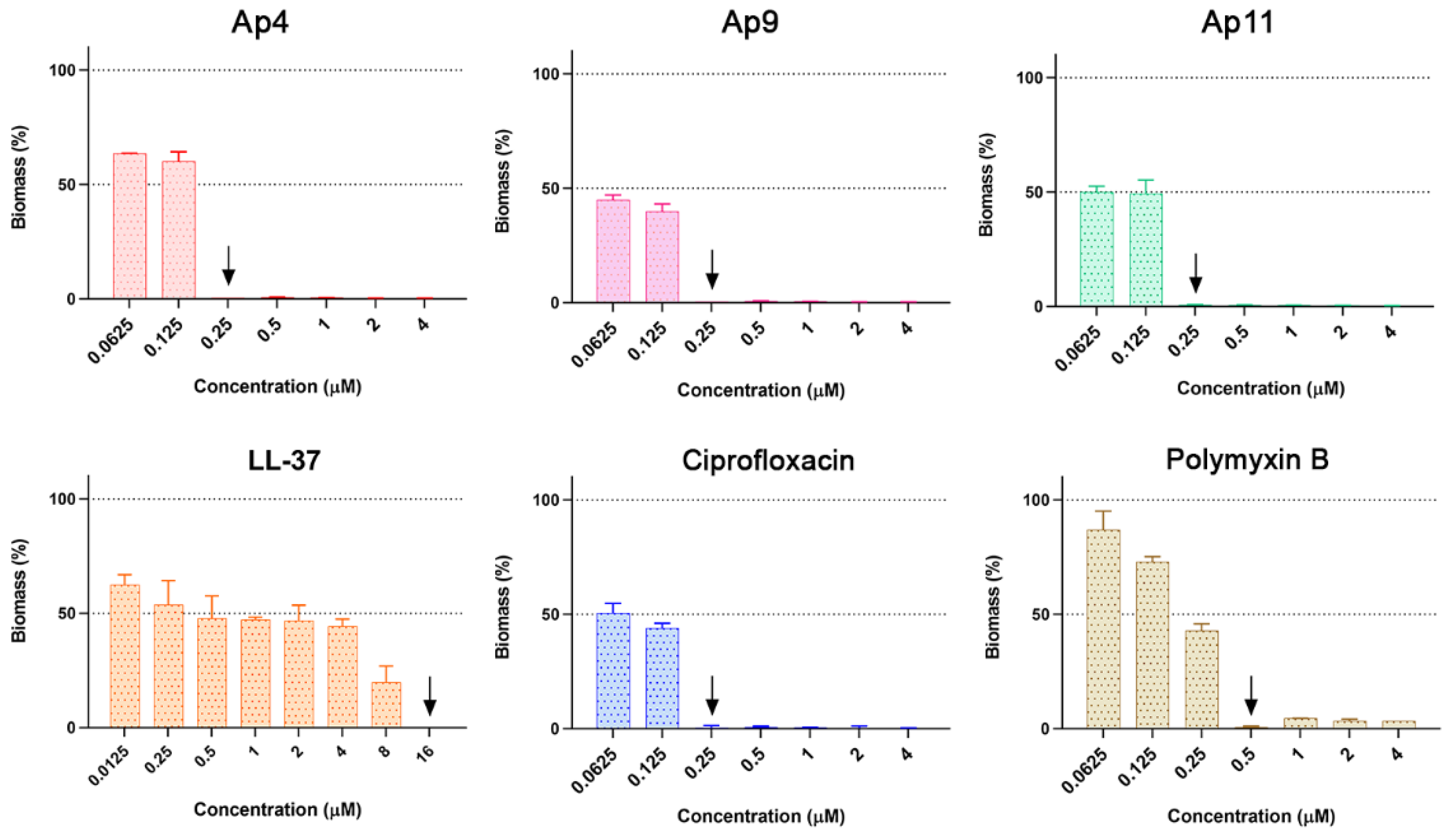
2.7. Abarenicins Prevent Biofilm Formation and Kill Embedded Cells Inside Biofilms Formed by P. aeruginosa including Its Drug-Resistance Clinical Isolates
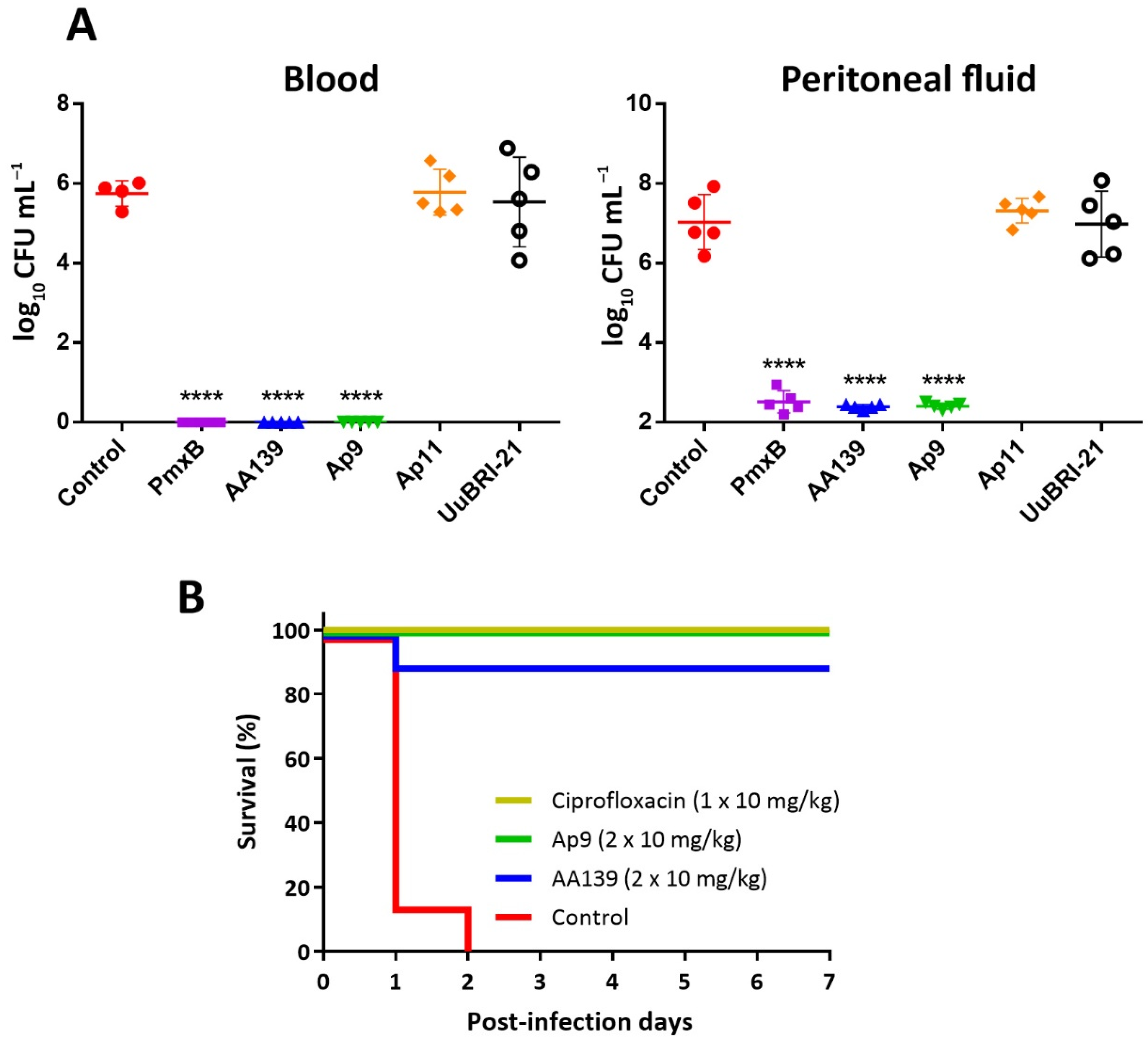
2.8. In Vivo Efficiency of the Novel Peptides
3. Materials and Methods
3.1. Transcriptome Assembly
3.2. Expression and Purification of the Antimicrobial Peptides
3.3. Antimicrobial Assay
3.4. Hemolytic Assay
3.5. Cytotoxic Assay
3.6. Assessment of Bacterial Membrane Permeabilization
3.7. Resistance Induction Assay
3.8. Stability in Serum
3.9. Measurement of Anti-Biofilm Activity
3.10. The Effect of Peptides on Established Biofilms
3.11. Animal Studies
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nagvekar, V.; Sawant, S.; Amey, S. Prevalence of Multidrug-Resistant Gram-Negative Bacteria Cases at Admission in a Multispeciality Hospital. J. Glob. Antimicrob. Resist. 2020, 22, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Aleshina, G.M.; Balandin, S.V.; Krasnosdembskaya, A.D.; Markelov, M.L.; Frolova, E.I.; Leonova, Y.F.; Tagaev, A.A.; Krasnodembsky, E.G.; Kokryakov, V.N. Purification and Primary Structure of Two Isoforms of Arenicin, a Novel Antimicrobial Peptide from Marine Polychaeta Arenicola marina. FEBS Lett. 2004, 577, 209–214. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Balandin, S.V.; Ovchinnikova, T.V. Design of Antimicrobial Peptide Arenicin Analogs with Improved Therapeutic Indices: Design of Arenicin Analogs with Improved Therapeutic Indices. J. Pept. Sci. 2015, 21, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Myshkin, M.Y.; Shenkarev, Z.O.; Ovchinnikova, T.V. Dimerization of the Antimicrobial Peptide Arenicin Plays a Key Role in the Cytotoxicity but Not in the Antibacterial Activity. Biochem. Biophys. Res. Commun. 2017, 482, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Krenev, I.A.; Umnyakova, E.S.; Eliseev, I.E.; Dubrovskii, Y.A.; Gorbunov, N.P.; Pozolotin, V.A.; Komlev, A.S.; Panteleev, P.V.; Balandin, S.V.; Ovchinnikova, T.V.; et al. Antimicrobial Peptide Arenicin-1 Derivative Ar-1-(C/A) as Complement System Modulator. Mar. Drugs 2020, 18, 631. [Google Scholar] [CrossRef] [PubMed]
- Edwards, I.A.; Elliott, A.G.; Kavanagh, A.M.; Zuegg, J.; Blaskovich, M.A.T.; Cooper, M.A. Contribution of Amphipathicity and Hydrophobicity to the Antimicrobial Activity and Cytotoxicity of β-Hairpin Peptides. ACS Infect. Dis. 2016, 2, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An Amphipathic Peptide with Antibiotic Activity against Multidrug-Resistant Gram-Negative Bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef] [PubMed]
- Stanovova, M.V.; Gazizova, G.R.; Gorbushin, A.M. Transcriptomic Profiling of Immune-associated Molecules in the Coelomocytes of Lugworm Arenicola marina (Linnaeus, 1758). J. Exp. Zool. Part B Mol. Dev. Evol. 2022, jez.b.23135. [Google Scholar] [CrossRef] [PubMed]
- Tilic, E.; Lehrke, J.; Bartolomaeus, T. Homology and Evolution of the Chaetae in Echiura (Annelida). PLoS ONE 2015, 10, e0120002. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Tsarev, A.V.; Safronova, V.N.; Reznikova, O.V.; Bolosov, I.A.; Sychev, S.V.; Shenkarev, Z.O.; Ovchinnikova, T.V. Structure Elucidation and Functional Studies of a Novel β-Hairpin Antimicrobial Peptide from the Marine Polychaeta Capitella Teleta. Mar. Drugs 2020, 18, 620. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Bolosov, I.A.; Kalashnikov, A.À.; Kokryakov, V.N.; Shamova, O.V.; Emelianova, A.A.; Balandin, S.V.; Ovchinnikova, T.V. Combined Antibacterial Effects of Goat Cathelicidins With Different Mechanisms of Action. Front. Microbiol. 2018, 9, 2983. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.; Straus, S. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef]
- Edwards, I.A.; Elliott, A.G.; Kavanagh, A.M.; Blaskovich, M.A.T.; Cooper, M.A. Structure–Activity and –Toxicity Relationships of the Antimicrobial Peptide Tachyplesin-1. ACS Infect. Dis. 2017, 3, 917–926. [Google Scholar] [CrossRef]
- Malmsten, M. Antimicrobial Peptides. Upsala J. Med. Sci. 2014, 119, 199–204. [Google Scholar] [CrossRef]
- Yin, L.M.; Edwards, M.A.; Li, J.; Yip, C.M.; Deber, C.M. Roles of Hydrophobicity and Charge Distribution of Cationic Antimicrobial Peptides in Peptide-Membrane Interactions. J. Biol. Chem. 2012, 287, 7738–7745. [Google Scholar] [CrossRef]
- Jenssen, H.; Aspmo, S.I. Serum Stability of Peptides. In Peptide-Based Drug Design; Otvos, L., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2008; Volume 494, pp. 177–186. ISBN 978-1-58829-990-1. [Google Scholar]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and Consequences of Bacterial Resistance to Antimicrobial Peptides. Drug Resist. Updates 2016, 26, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Komp Lindgren, P.; Karlsson, Å.; Hughes, D. Mutation Rate and Evolution of Fluoroquinolone Resistance in Escherichia Coli Isolates from Patients with Urinary Tract Infections. Antimicrob. Agents Chemother. 2003, 47, 3222–3232. [Google Scholar] [CrossRef]
- Vimberg, V.; Buriánková, K.; Mazumdar, A.; Branny, P.; Novotná, G.B. Role of Membrane Proteins in Bacterial Resistance to Antimicrobial Peptides. Med. Res. Rev. 2022, 42, 1023–1036. [Google Scholar] [CrossRef]
- Safronova, V.N.; Panteleev, P.V.; Sukhanov, S.V.; Toropygin, I.Y.; Bolosov, I.A.; Ovchinnikova, T.V. Mechanism of Action and Therapeutic Potential of the β-Hairpin Antimicrobial Peptide Capitellacin from the Marine Polychaeta Capitella Teleta. Mar. Drugs 2022, 20, 167. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, P.; Singh, S.; Agarwal, V. Microbial Biofilms. In Bacterial Biofilms; Dincer, S., Sümengen Özdenefe, M., Arkut, A., Eds.; IntechOpen: London, UK, 2020; ISBN 978-1-78985-899-0. [Google Scholar]
- de la Fuente-Núñez, C.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.M.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E.W. Inhibition of Bacterial Biofilm Formation and Swarming Motility by a Small Synthetic Cationic Peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef]
- Segev-Zarko, L.; Saar-Dover, R.; Brumfeld, V.; Mangoni, M.L.; Shai, Y. Mechanisms of Biofilm Inhibition and Degradation by Antimicrobial Peptides. Biochem. J. 2015, 468, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, S.; Nair, H.A.S.; Lee, K.W.K.; Ong, J.; Goh, J.Q.J.; Kjelleberg, S.; Rice, S.A. Pseudomonas Aeruginosa PAO1 Exopolysaccharides Are Important for Mixed Species Biofilm Community Development and Stress Tolerance. Front. Microbiol. 2015, 6, 851. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Balandin, S.V.; Ovchinnikova, T.V. Effect of Arenicins and Other β-Hairpin Antimicrobial Peptides on Pseudomonas Aeruginosa PAO1 Biofilms. Pharm. Chem. J. 2017, 50, 715–720. [Google Scholar] [CrossRef]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.W.; Rehm, B.H.A.; Hancock, R.E.W. Human Host Defense Peptide LL-37 Prevents Bacterial Biofilm Formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as Intermicrobial Signaling Agents Instead of Weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside Antibiotics Induce Bacterial Biofilm Formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Paunova-Krasteva, T.; Haladjova, E.; Petrov, P.; Forys, A.; Trzebicka, B.; Topouzova-Hristova, T.; Stoitsova, S.R. Destruction of Pseudomonas aeruginosa Pre-Formed Biofilms by Cationic Polymer Micelles Bearing Silver Nanoparticles. Biofouling 2020, 36, 679–695. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Ovchinnikova, T.V. Bioengineering and Functional Characterization of Arenicin Shortened Analogs with Enhanced Antibacterial Activity and Cell Selectivity: Bioengineering of Arenicin Shortened Analogs with Enhanced Selectivity. J. Pept. Sci. 2016, 22, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Minimum Inhibitory Concentration, µM | |
|---|---|---|
| MHB + 0.9% NaCl | MHB + 0.9% NaCl + 25% FBS | |
| Abarenicin | 0.25 | 1 |
| AA139 | 0.25 | 1 |
| UuBRI-21 | 0.25 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safronova, V.N.; Bolosov, I.A.; Kruglikov, R.N.; Korobova, O.V.; Pereskokova, E.S.; Borzilov, A.I.; Panteleev, P.V.; Ovchinnikova, T.V. Novel β-Hairpin Peptide from Marine Polychaeta with a High Efficacy against Gram-Negative Pathogens. Mar. Drugs 2022, 20, 517. https://doi.org/10.3390/md20080517
Safronova VN, Bolosov IA, Kruglikov RN, Korobova OV, Pereskokova ES, Borzilov AI, Panteleev PV, Ovchinnikova TV. Novel β-Hairpin Peptide from Marine Polychaeta with a High Efficacy against Gram-Negative Pathogens. Marine Drugs. 2022; 20(8):517. https://doi.org/10.3390/md20080517
Chicago/Turabian StyleSafronova, Victoria N., Ilia A. Bolosov, Roman N. Kruglikov, Olga V. Korobova, Eugenia S. Pereskokova, Alexander I. Borzilov, Pavel V. Panteleev, and Tatiana V. Ovchinnikova. 2022. "Novel β-Hairpin Peptide from Marine Polychaeta with a High Efficacy against Gram-Negative Pathogens" Marine Drugs 20, no. 8: 517. https://doi.org/10.3390/md20080517