
Immunomodulatory Activity In Vitro and In Vivo of a Sulfated Polysaccharide with Novel Structure from the Green Alga Ulvaconglobata Kjellman

Abstract
:
1. Introduction
2. Results and Discussion
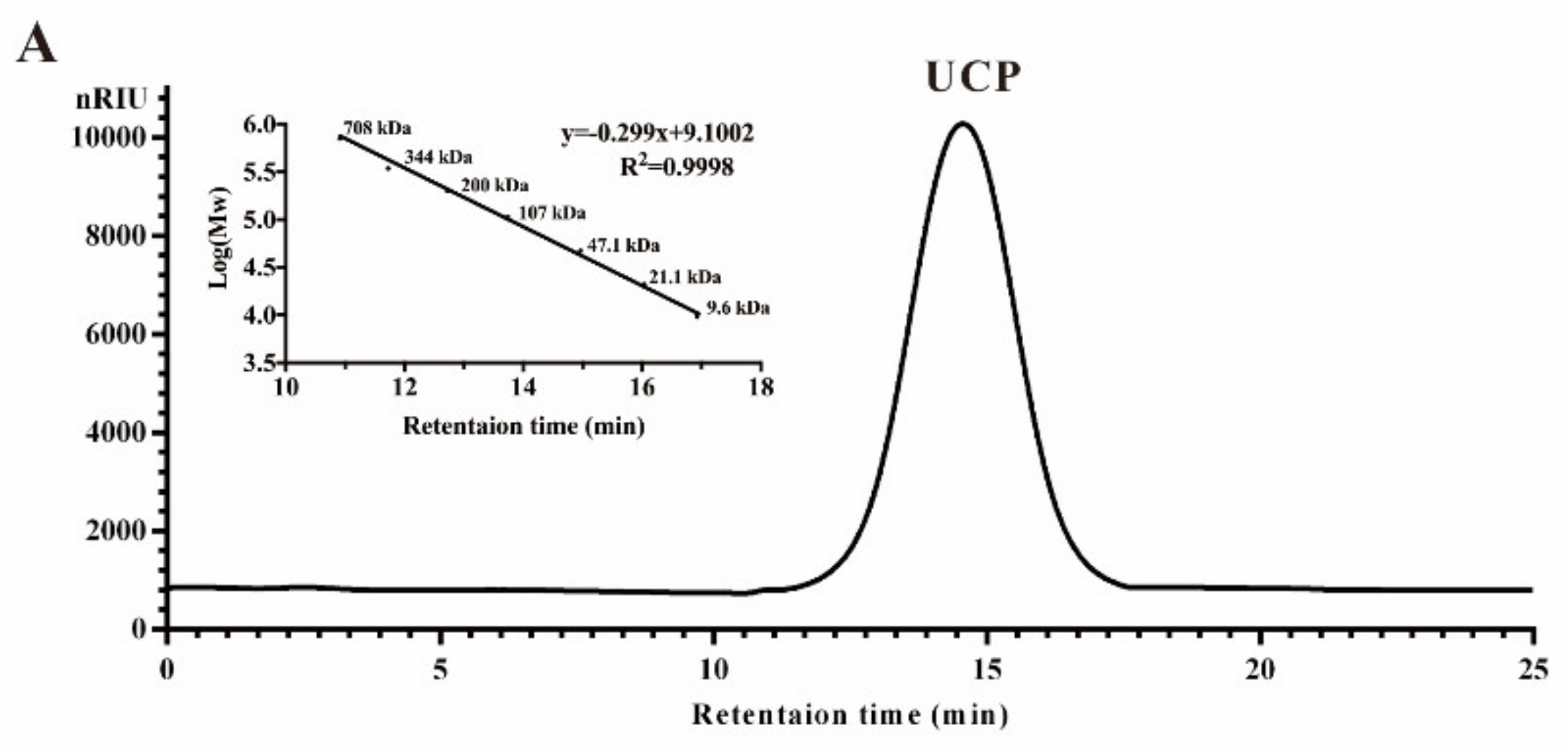
2.1. Chemical Composition of UCP
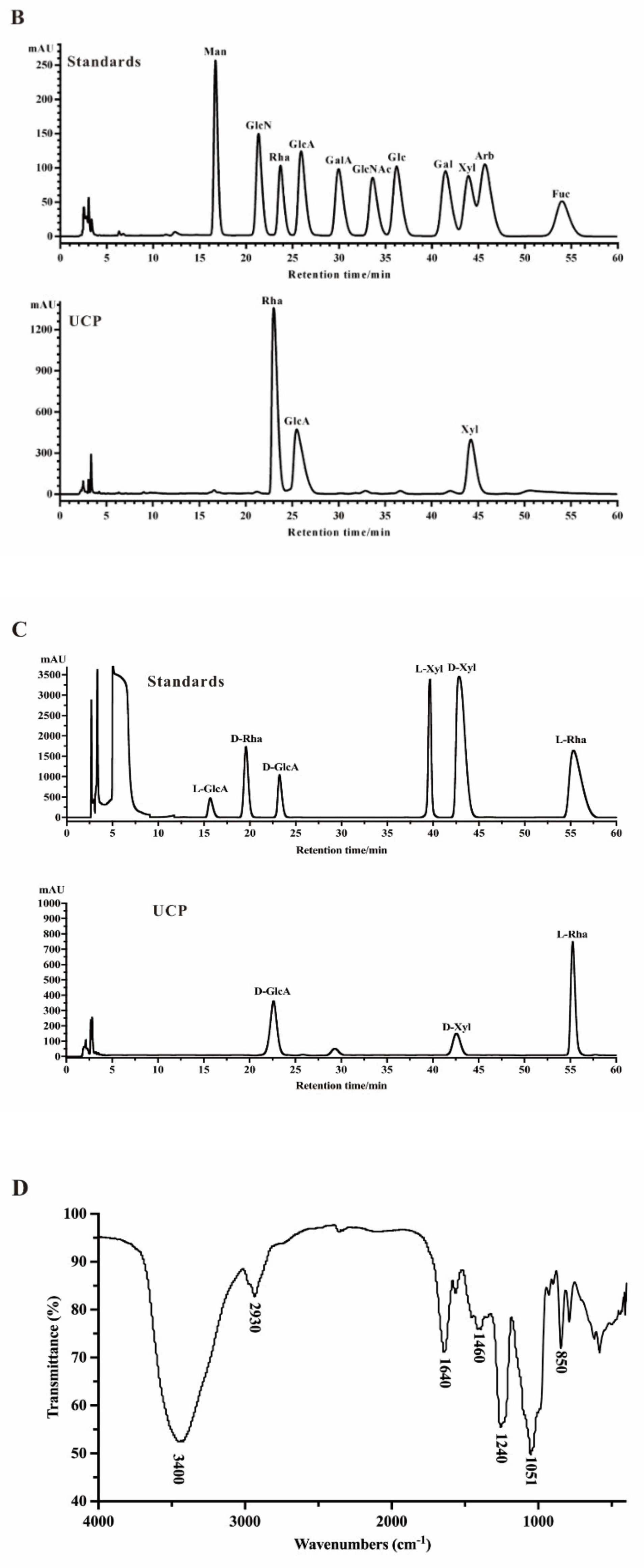
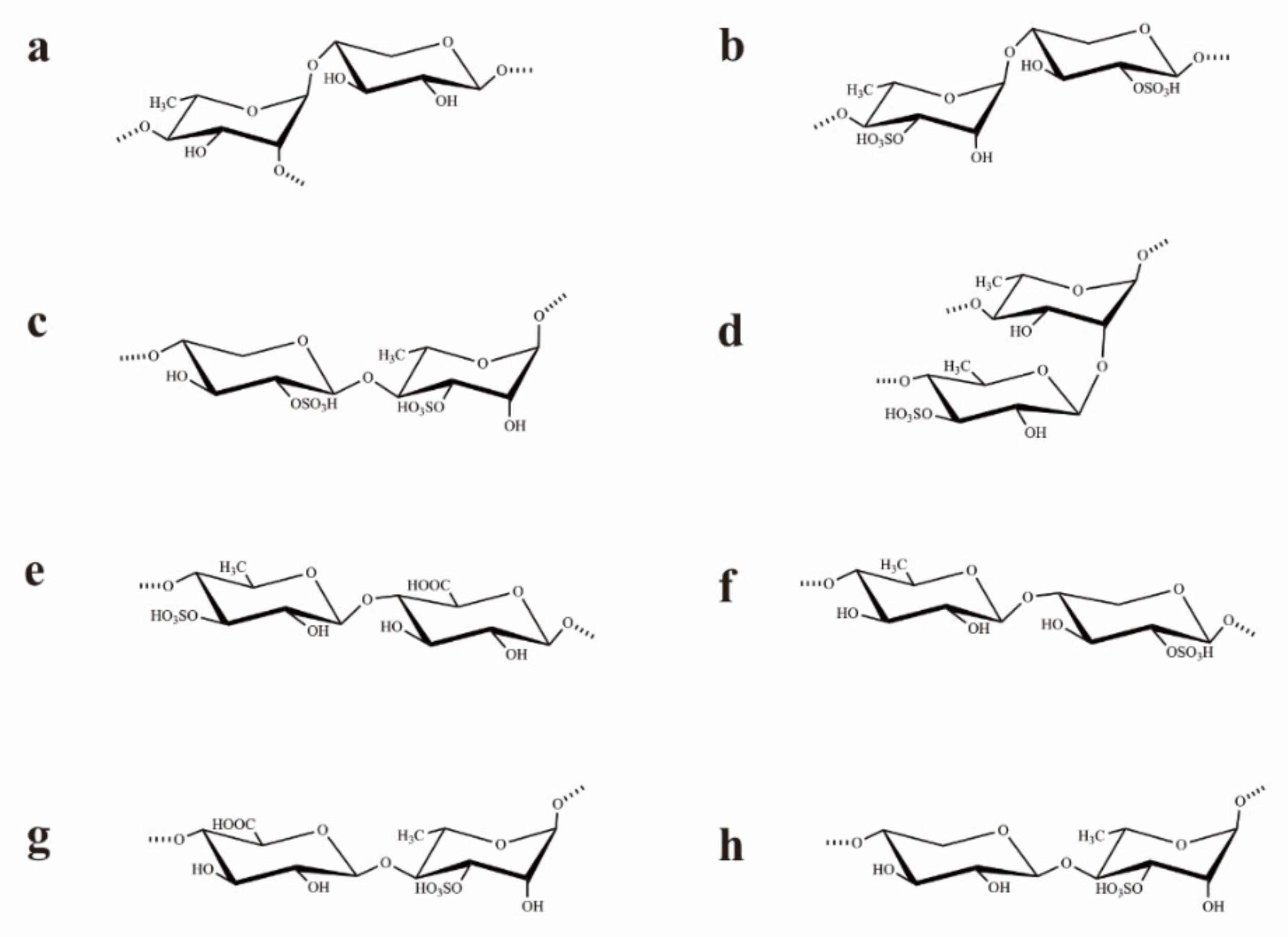
2.2. Structural Characteristics of UCP
2.2.1. Methylation Analysis
2.2.2. NMR Analysis of UCP
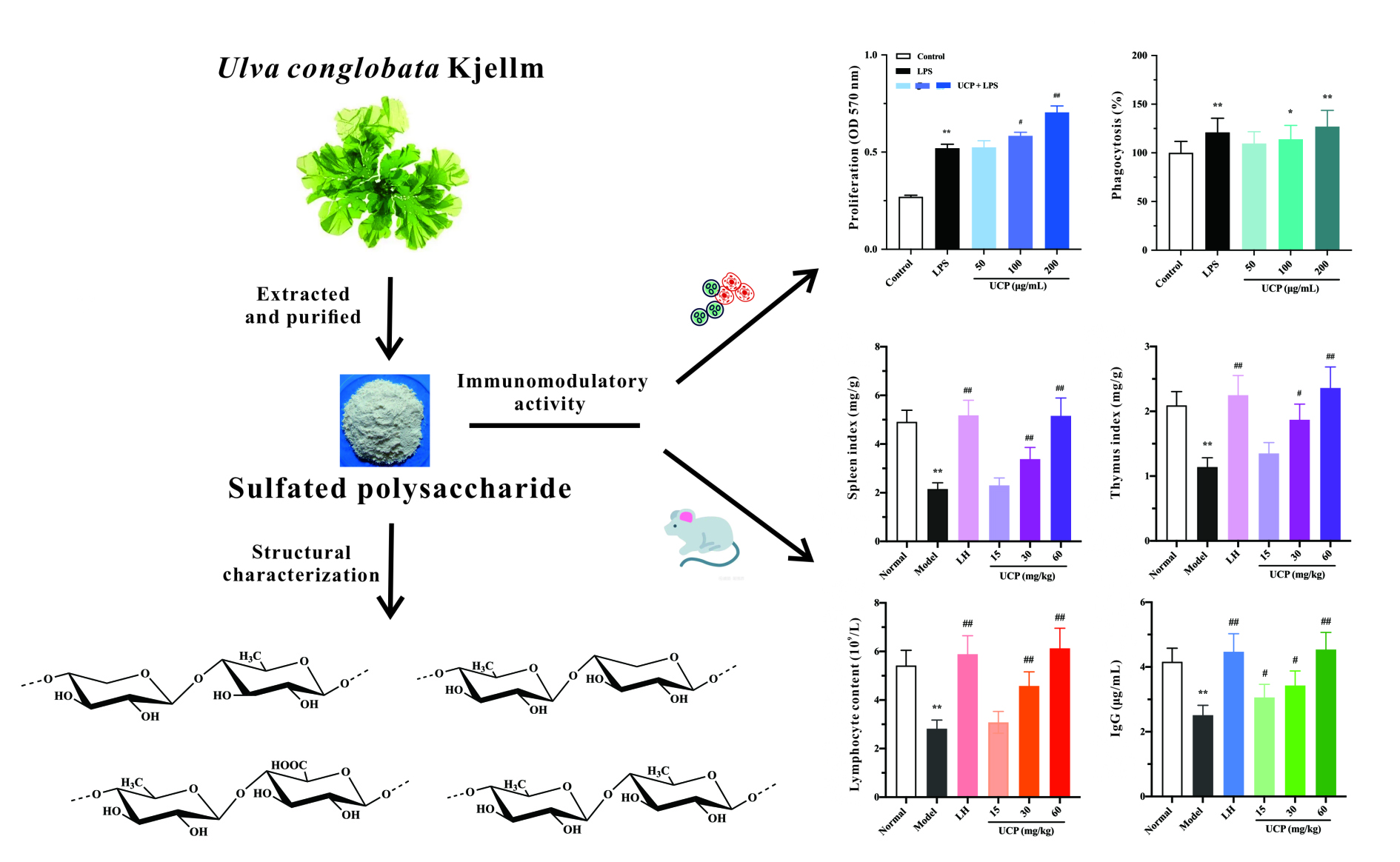
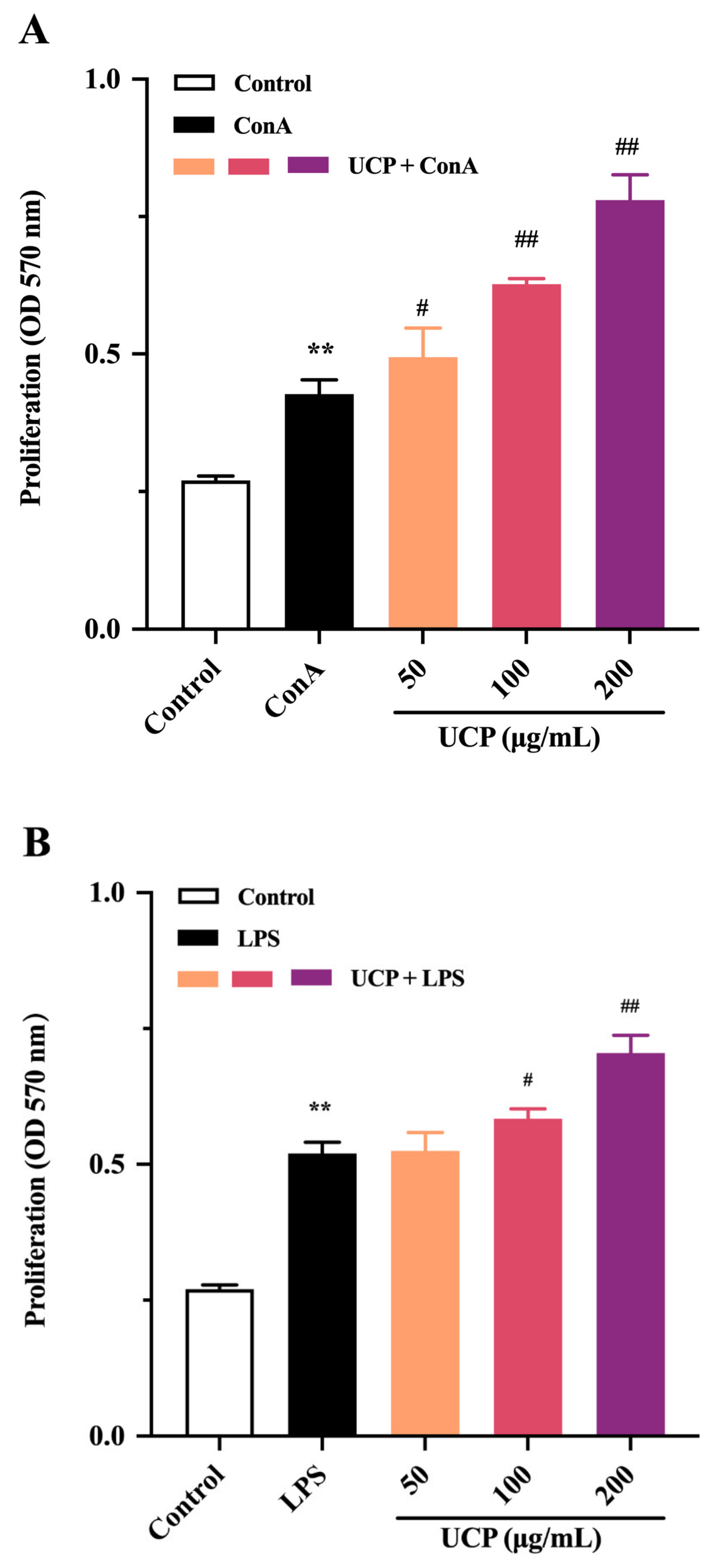
2.3. Effect of UCP on Lymphocyte Proliferation
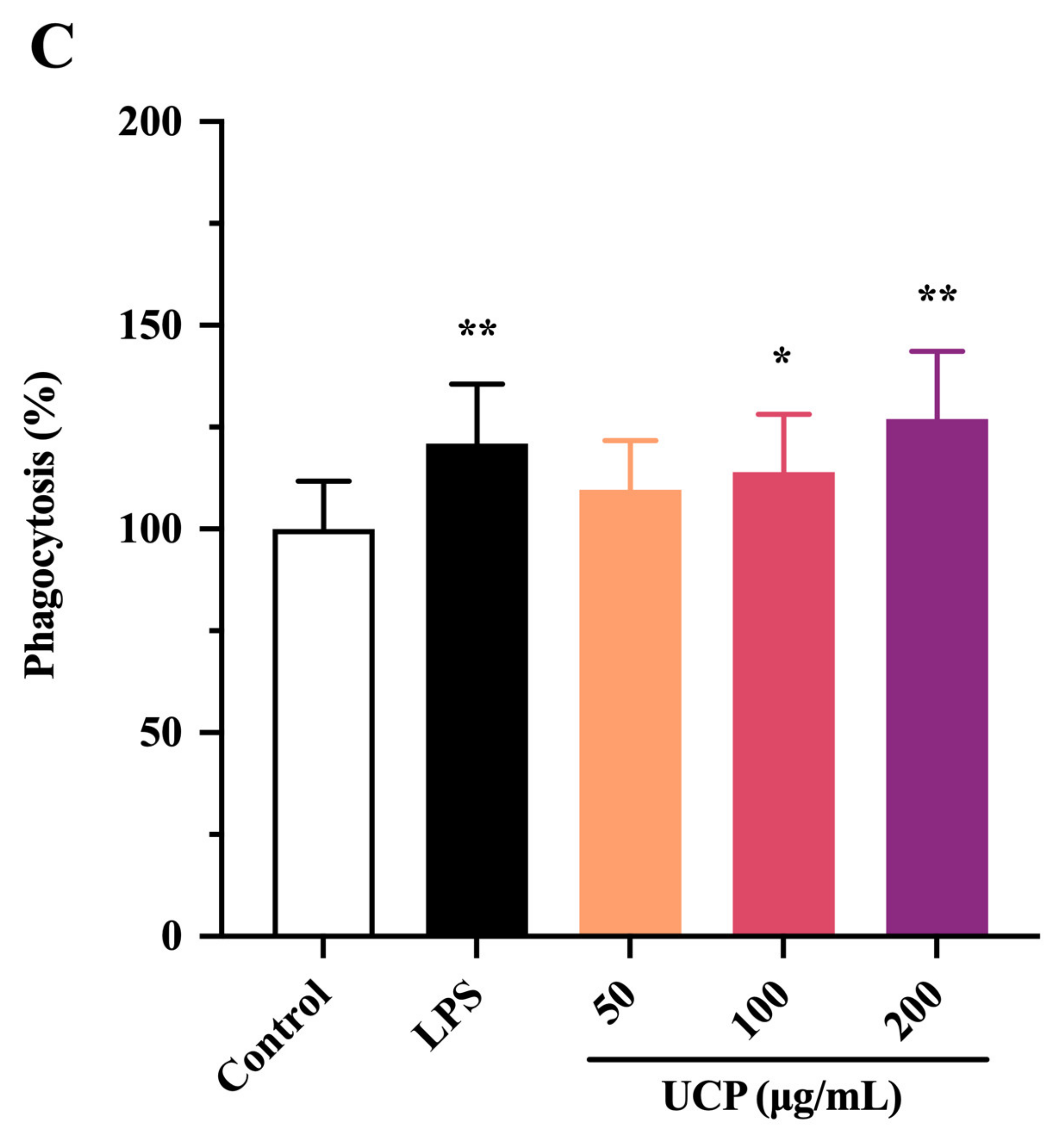
2.4. Effect of UCP on Phagocytic Activity of RAW246.7 Cells
2.5. Immunomodulatory Activity In Vivo of UCP
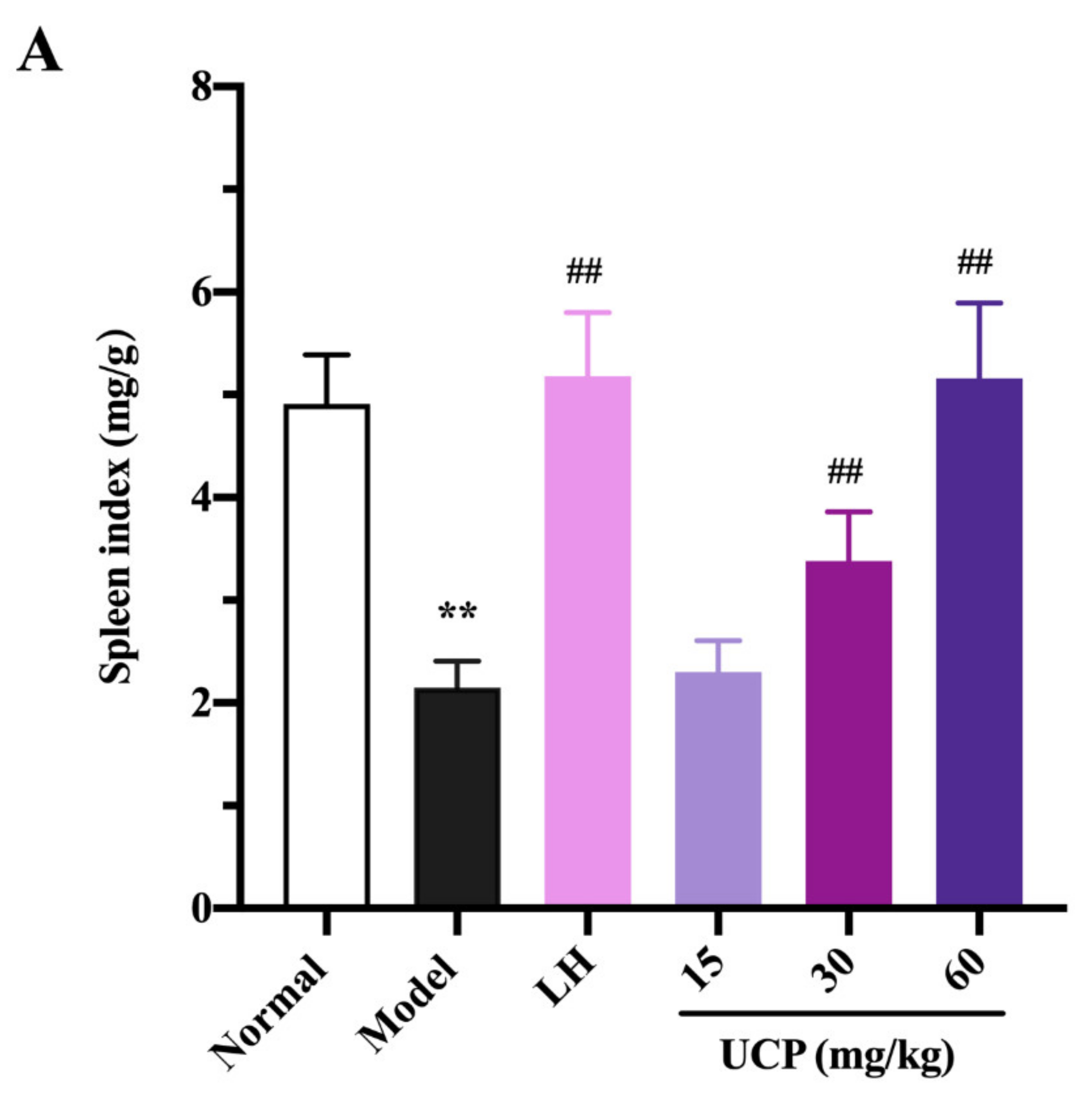
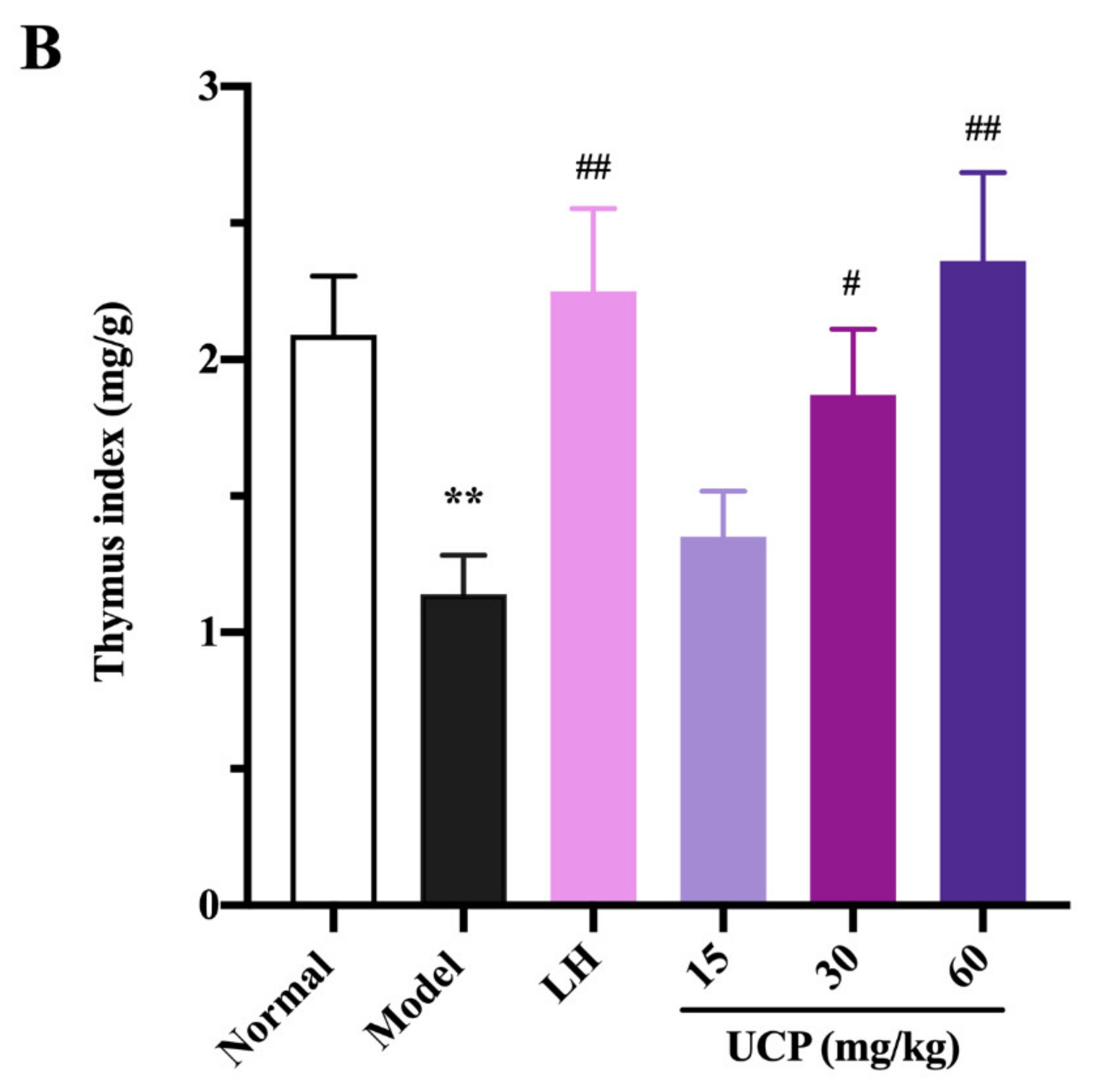
2.5.1. Effect of UCP on Thymus and Spleen Indices
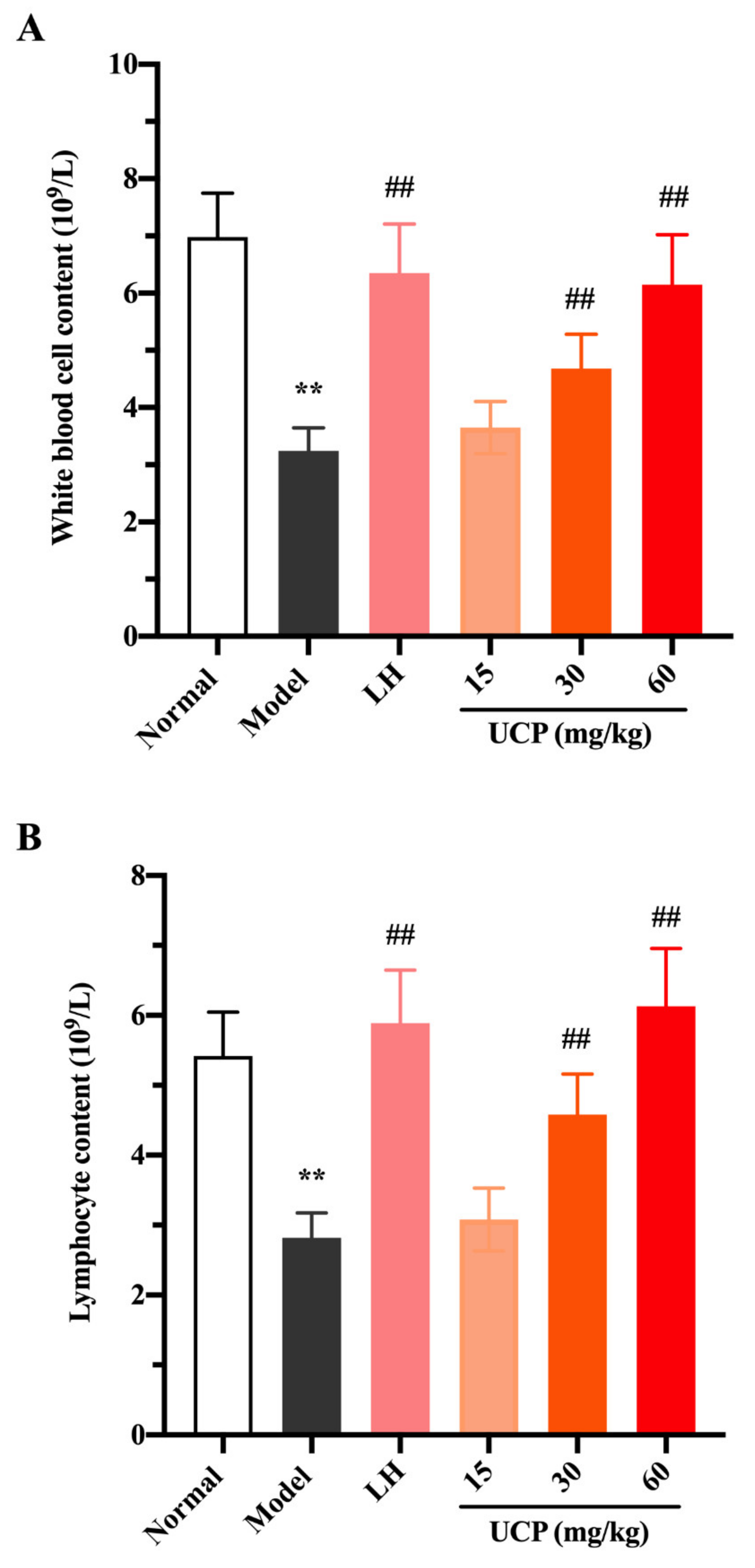
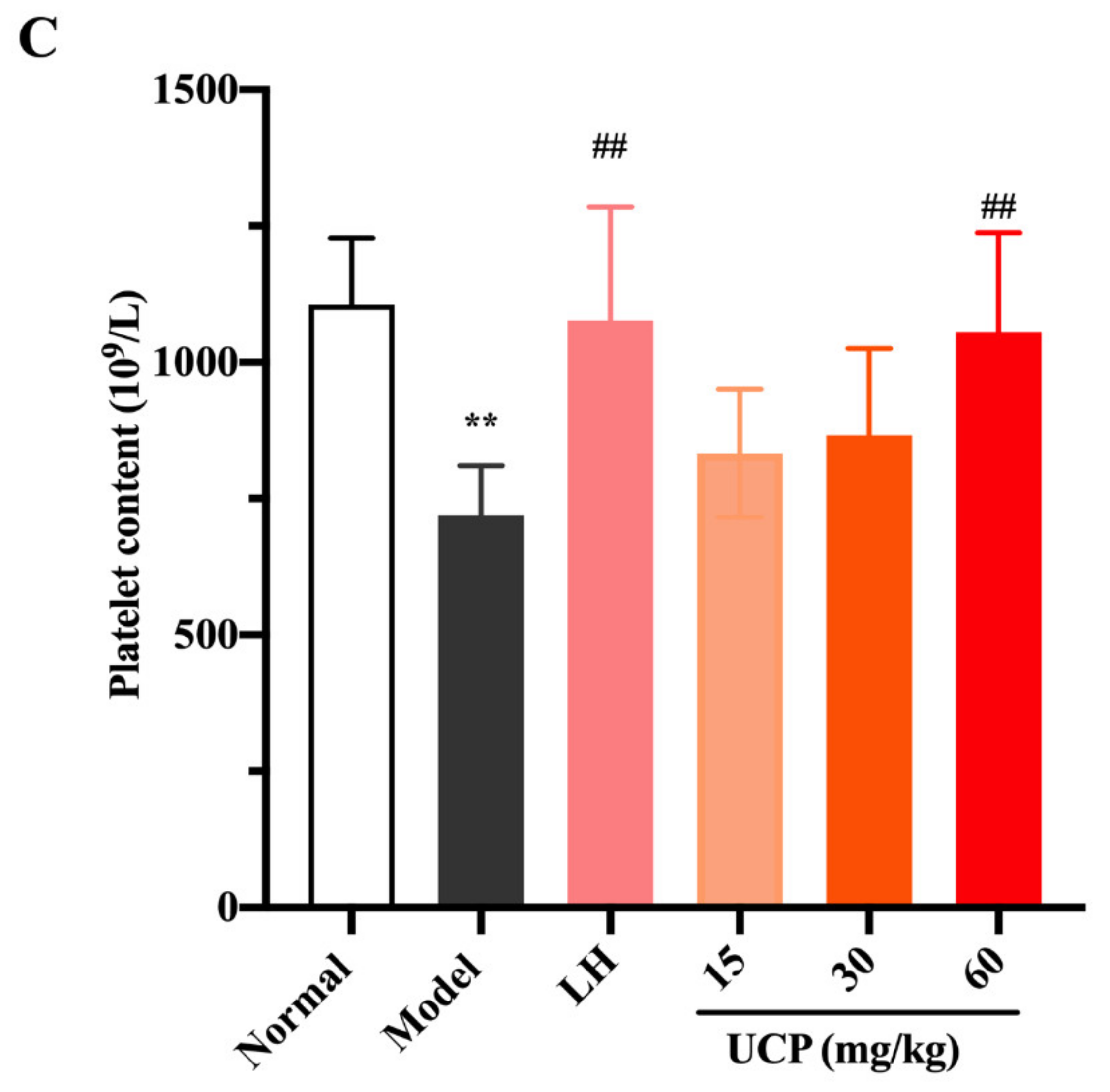
2.5.2. Effect of UCP on Peripheral Blood Cells
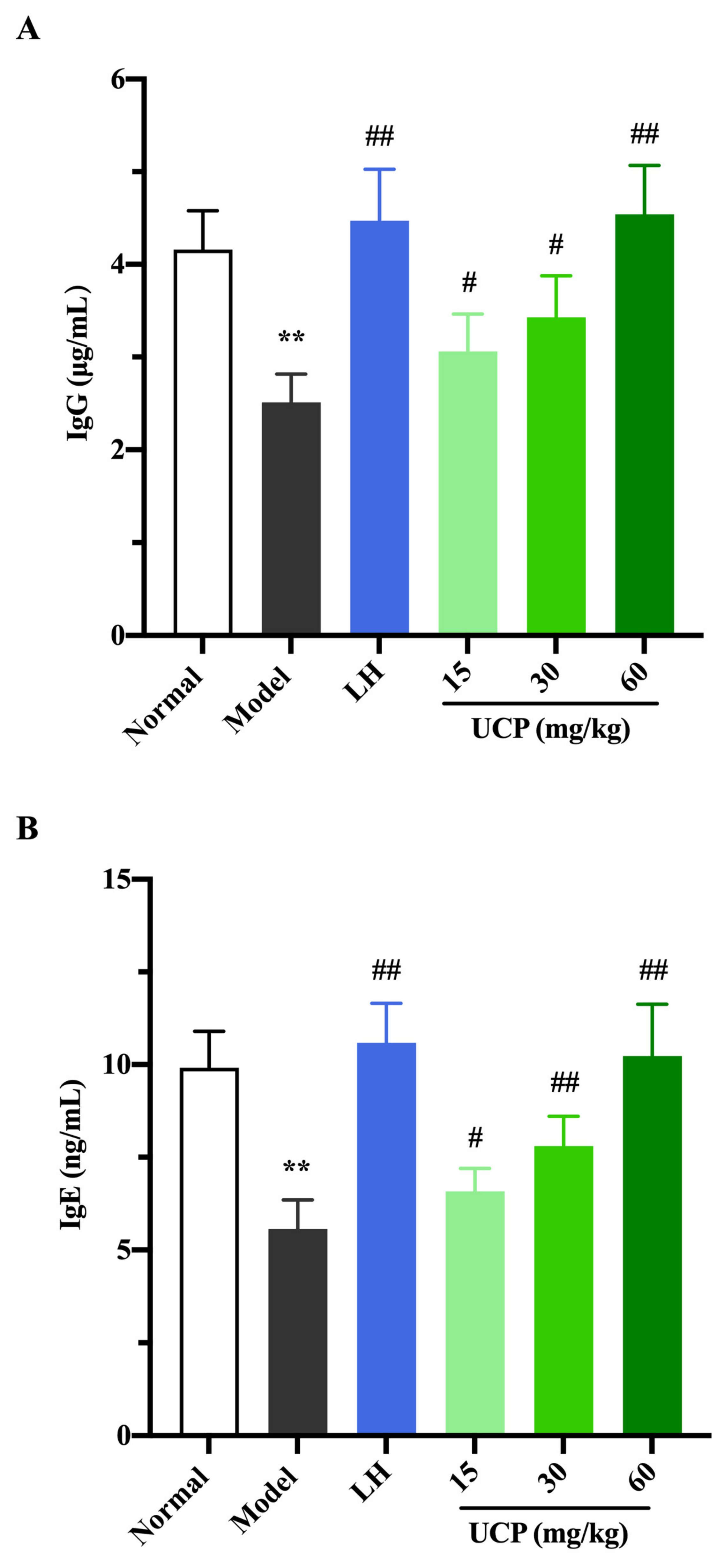
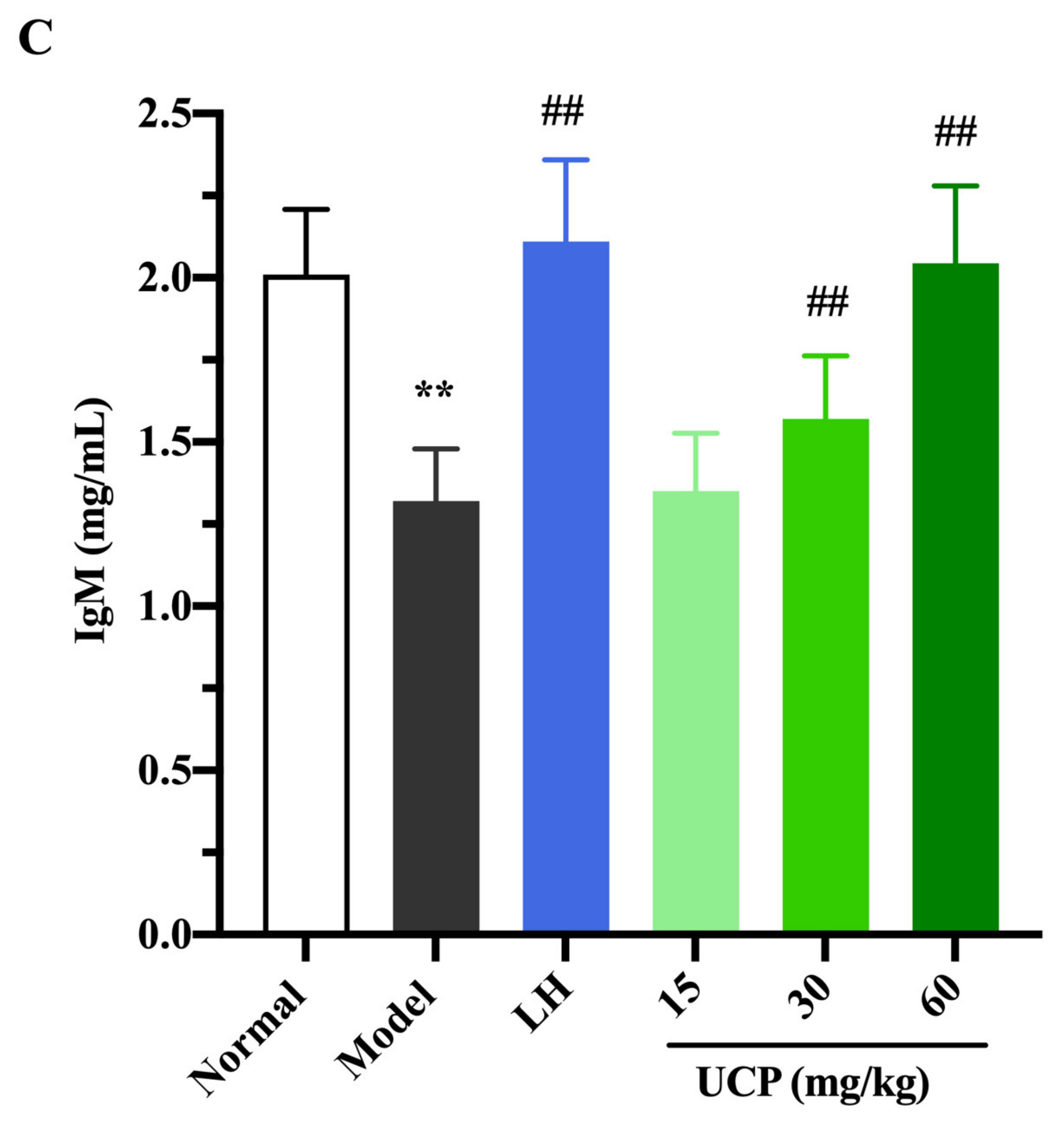
2.5.3. Influence of UCP on Serum Antibody Level
3. Materials and Methods
3.1. Materials
3.2. Cell Culture
3.3. Animals
3.4. Isolation and Purification of the Sulfated Polysaccharide
3.5. Structural Characterization of Polysaccharide
3.6. Spectroscopy Analysis
3.7. Preparation of Spleen Lymphocytes
3.8. Assay of Spleen Lymphocyte Proliferation
3.9. Phagocytic Activity of RAW264.7 Cells
3.10. Animal Experimental Design
3.11. Analysis of Spleen and Thymus Indices
3.12. Peripheral Blood Cell Counts
3.13. Analysis of Immunoglobulins in Serum
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fernández, P.V.; Arata, P.X.; Ciancia, M. Polysaccharides from Codium species. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71, pp. 253–278. [Google Scholar]
- Fischer, A.M.; Tapon-Bretaudiere, J.; Boisson, C.; Durand, P.; Jozefonvicz, J. Anticoagulant properties of a fucoidan fraction. Thromb. Res. 1991, 64, 143–154. [Google Scholar]
- Synytsya, A.; Kim, W.J.; Kim, S.M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Il Park, Y. Structure and antitumor activity of fucoidan isolated from sporophyll of korean brown seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Na, Y.S.; Kim, W.J.; Kim, S.M.; Park, J.K.; Lee, S.M.; Kim, S.O.; Synytsya, A.; Park, Y.I. Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens. Int. Immunopharmacol. 2010, 10, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Artan, M.; Li, Y.; Karadeniz, F.; Lee, S.H.; Kim, M.M.; Kim, S.K. Anti-HIV-1 activity of phloroglucinol derivative, 6,6′-bieckol from Ecklonia cava. Bioorg. Med. Chem. 2008, 16, 7921–7926. [Google Scholar] [CrossRef]
- Sun, L.Q.; Wang, C.H.; Shi, Q.J.; Ma, C.H. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Kim, J.; Cho, M.; Karnjanapratum, S.; Shin, I.; You, S. In vitro and in vivo immunomodulatory activity of sulfated polysaccharides from Enteromorpha prolifera. Int. J. Biol. Macromol. 2011, 49, 1051–1058. [Google Scholar] [CrossRef]
- Liu, X.; Wang, S.Y.; Cao, S.J.; He, X.X.; Qin, L.; He, M.J.; Yang, Y.J.; Hao, J.J.; Mao, W.J. Structural characteristics and anticoagulant property in vitro and in vivo of a seaweed sulfated rhamnan. Mar. Drugs 2018, 16, 243. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Du, P.; Liu, X.; Cao, S.J.; Qin, L.; He, M.J.; He, X.X.; Mao, W.J. Anticoagulant properties of a green algal rhamnan-type sulfated polysaccharide and its low-molecular-weight fragments prepared by mild acid degradation. Mar. Drugs 2018, 16, 445. [Google Scholar] [CrossRef] [Green Version]
- Tabarsa, M.; Shin, I.-S.; Lee, J.H.; Surayot, U.; Park, W.; You, S. An immune-enhancing water-soluble α-glucan from Chlorella vulgaris and structural characteristics. Food Sci. Biotechnol. 2015, 24, 1933–1941. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Jeyashoke, N.; Yeh, C.H.; Song, Y.J.; Hua, K.F.; Chao, L.K. Immunostimulatory bioactivity of algal polysaccharides from Chlorella pyrenoidosa activates macrophages via toll-like receptor 4. J. Agric. Food Chem. 2010, 58, 927–936. [Google Scholar] [CrossRef]
- Suárez, E.R.; Kralovec, J.A.; Bruce Grindley, T. Isolation of phosphorylated polysaccharides from algae: The immunostimulatory principle of Chlorella pyrenoidosa. Carbohydr. Res. 2010, 345, 1190–1204. [Google Scholar] [CrossRef]
- Guo, Q.; Shao, Q.; Xu, W.; Rui, L.; Sumi, R.; Eguchi, F.; Li, Z. Immunomodulatory and anti-IBDV activities of the polysaccharide AEX from Coccomyxa gloeobotrydiformis. Mar. Drugs 2017, 15, 36. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.B.; Ohta, Y.; Hayashi, K.; Hayashi, T. Immunostimulating effects of a sulfated galactan from Codium fragile. Carbohydr Res. 2010, 345, 1452–1454. [Google Scholar] [CrossRef]
- Hao, H.; Han, Y.; Yang, L.; Hu, L.; Duan, X.; Yang, X.; Huang, R. Structural characterization and immunostimulatory activity of a novel polysaccharide from green alga Caulerpa racemosa var peltata. Int. J. Biol. Macromol. 2019, 134, 891–900. [Google Scholar] [CrossRef]
- Glasson, C.R.K.; Sims, I.M.; Carnachan, S.M.; de Nys, R.; Magnusson, M. A cascading biorefinery process targeting sulfated polysaccharides (Ulvan) from Ulva ohnoi. Algal Res. 2017, 27, 383–391. [Google Scholar] [CrossRef]
- Abd-elrahman, G.E.; Ahmed, O.; Abdel-Reheim, E.; Abdel-Hamid, A.-H. Ulva lactuca polysaccharides prevent wistar wat breast carcinogenesis through the augmentation of apoptosis, enhancement of antioxidant defense system, and suppression of inflammation. Breast Cancer Targets Ther. 2017, 9, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Leiro, J.M.; Castro, R.; Arranz, J.A.; Lamas, J. Immunomodulating activities of acidic sulphated polysaccharides obtained from the seaweed Ulva rigida C. Agardh. Int. Immunopharmacol. 2007, 7, 879–888. [Google Scholar] [CrossRef]
- Tabarsa, M.; Han, J.H.; Kim, C.Y.; You, S.G. Molecular characteristics and immunomodulatory activities of water-soluble sulfated polysaccharides from Ulva pertusa. J. Med. Food 2012, 15, 135–144. [Google Scholar] [CrossRef]
- Kidgell, J.T.; Glasson, C.R.K.; Magnusson, M.; Vamvounis, G.; Sims, I.M.; Carnachan, S.M.; Hinkley, S.F.R.; Lopata, A.L.; de Nys, R.; Taki, A.C. The molecular weight of Ulvan affects the in vitro inflammatory response of a murine macrophage. Int. J. Biol. Macromol. 2020, 150, 839–848. [Google Scholar] [CrossRef]
- Peasura, N.; Laohakunjit, N.; Kerdchoechuen, O.; Vongsawasdi, P.; Chao, L.K. Assessment of biochemical and immunomodulatory activity of sulphated polysaccharides from Ulva intestinalis. Int. J. Biol. Macromol. 2016, 91, 269–277. [Google Scholar] [CrossRef]
- Cassolato, J.E.F.; Noseda, M.D.; Pujol, C.A.; Pellizzari, F.M.; Damonte, E.B.; Duarte, M.E.R. Chemical structure and antiviral activity of the sulfated heterorhamnan isolated from the green seaweed Gayralia oxysperma. Carbohydr. Res. 2008, 343, 3085–3095. [Google Scholar] [CrossRef]
- Wu, D.S.; Chen, Y.H.; Wan, X.Z.; Liu, D.; Wen, Y.X.; Chen, X.H.; Zhao, C. Structural characterization and hypoglycemic effect of green alga Ulva lactuca oligosaccharide by regulating microRNAs in Caenorhabditis elegans. Algal Res. 2020, 51, 102083. [Google Scholar] [CrossRef]
- Hu, P.; Li, Z.X.; Chen, M.C.; Sun, Z.L.; Ling, Y.; Jiang, J.; Huang, C.G. Structural elucidation and protective role of a polysaccharide from Sargassum fusiforme on ameliorating learning and memory deficiencies in mice. Carbohydr. Polym. 2016, 139, 150–158. [Google Scholar] [CrossRef]
- Ropellato, J.; Carvalho, M.M.; Ferreira, L.G.; Noseda, M.D.; Zuconelli, C.R.; Gonçalves, A.G.; Ducatti, D.R.B.; Kenski, J.C.N.; Nasato, P.L.; Winnischofer, S.M.B.; et al. Sulfated heterorhamnans from the green seaweed Gayralia oxysperma: Partial depolymerization, chemical structure and antitumor activity. Carbohydr. Polym. 2015, 117, 476–485. [Google Scholar] [CrossRef]
- Lahaye, M. NMR spectroscopic characterisation of oligosaccharides from two Ulva rigida ulvan samples (Ulvales, Chlorophyta) degraded by a lyase. Carbohydr. Res. 1998, 12, 1355–1363. [Google Scholar] [CrossRef]
- Ray, B.; Lahaye, M. Cell-wall polysaccharides from the marine green alga Ulva “Rigida” (Ulvales, Chlorophyta). Extraction and chemical composition. Carbohydr. Res. 1995, 274, 251–261. [Google Scholar] [CrossRef]
- Zuo, S.Q.; Li, F.Y.; Gu, X.; Wei, Z.P.; Qiao, L.K.; Du, C.Y.; Chi, Y.Z.; Liu, R.Z.; Wang, P. Effects of low molecular weight polysaccharides from Ulva prolifera on the tolerance of Triticum aestivum to osmotic stress. Int. J. Biol. Macromol. 2021, 183, 12–22. [Google Scholar] [CrossRef]
- Moawad, M.N.; El-Sayed, A.A.M.; Abd El Latif, H.H.; El-Naggar, N.A.; Shams El-Din, N.G.; Tadros, H.R.Z. Chemical characterization and biochemical activity of polysaccharides isolated from egyptian Ulva fasciata Delile. Oceanologia 2022, 64, 117–130. [Google Scholar] [CrossRef]
- Han, Y.; Wu, Y.L.; Li, G.Q.; Li, M.Y.; Yan, R.; Xu, Z.L.; Lei, H.T.; Sun, Y.M.; Duan, X.W.; Hu, L.M.; et al. Structural characterization and transcript-metabolite correlation network of immunostimulatory effects of sulfated polysaccharides from green alga Ulva pertusa. Food Chem. 2021, 342, 128537. [Google Scholar] [CrossRef]
- Ni, T.; Yang, W.; Han, M.; Zhang, Y.; Shen, T.; Nie, H.; Zhou, Z.; Dai, Y.; Yang, Y.; Liu, P.; et al. Global intron retention mediated gene regulation during CD4 + T cell activation. Nucleic Acids Res. 2016, 44, 6817–6829. [Google Scholar] [CrossRef] [Green Version]
- Sarangi, I.; Ghosh, D.; Bhutia, S.K.; Mallick, S.K.; Maiti, T.K. Anti-tumor and immunomodulating effects of Pleurotus ostreatus mycelia-derived proteoglycans. Int. Immunopharmacol. 2006, 6, 1287–1297. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Thambiraj, S.R.; Phillips, M.; Koyyalamudi, S.R.; Reddy, N. Antioxidant activities and characterization of polysaccharides isolated from the seeds of Lupinus angustifolius. Ind. Crops Prod. 2015, 74, 950–956. [Google Scholar] [CrossRef]
- Haubitz, M.; Bohnenstengel, F.; Brunkhorst, R.; Schwab, M.; Hofmann, U.; Busse, D. Cyclophosphamide pharmacokinetics and dose requirements in patients with renal insufficiency. Kidney Int. 2002, 61, 1495–1501. [Google Scholar] [CrossRef] [Green Version]
- Yun, L.; Wu, T.; Li, Q.; Zhang, M. Dietary supplementation with purified wheat germ glycoprotein improve immunostimulatory activity in cyclophosphamide induced Balb/c mice. Int. J. Biol. Macromol. 2018, 118, 1267–1275. [Google Scholar] [CrossRef]
- Qi, Q.; Dong, Z.; Sun, Y.; Li, S.; Zhao, Z. Protective effect of bergenin against cyclophosphamide-induced immunosuppression by immunomodulatory effect and antioxidation in Balb/c mice. Molecules 2018, 23, 2668. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.M.; Lee, H.Y.; Shin, D.Y.; Lee, Y.H.; Yang, Y.J.; Lee, H.S.; Lee, J.O.; Choi, K.S.; Kang, J.H.; Cho, Y.H.; et al. Immunostimulatory activity of black rice bran in cyclophosphamide-induced immunosuppressed Rats. Nat. Prod. Commun. 2020, 15, 1–12. [Google Scholar] [CrossRef]
- You, L.; Gao, Q.; Feng, M.; Yang, B.; Ren, J.; Gu, L.; Cui, C.; Zhao, M. Structural characterisation of polysaccharides from Tricholoma Matsutake and their antioxidant and antitumour activities. Food Chem. 2013, 138, 2242–2249. [Google Scholar] [CrossRef]
- Shabbir, A.; Butt, H.I.; Shahzad, M.; Arshad, H.M.; Waheed, I. Immunostimulatory effect of methanolic leaves extract of Psidium guajava (Guava) on humoral and cell-mediated immunity in mice. J. Anim. Plant Sci. 2016, 26, 1492–1500. [Google Scholar]
- Zhou, C.; Yang, B.; Tian, Y.; Jiao, H.; Zheng, W.; Wang, J.; Guan, F. Immunomodulatory effect of human umbilical cord wharton’s jelly-derived mesenchymal stem cells on lymphocytes. Cell. Immunol. 2011, 272, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Ahlmann, M.; Hempel, G. The effect of cyclophosphamide on the immune system: Implications for clinical cancer therapy. Cancer Chemother. Pharmacol. 2016, 78, 661–671. [Google Scholar] [CrossRef]
- Qi, S.; Li, X.; Dong, Q.; Lai, H.; Porter, D.; Tian, S.; Hou, L.; Chen, X.; Li, X.; Wang, K. Chinese herbal medicine (Xiaoaiping) injections for chemotherapy-induced thrombocytopenia: A randomized, controlled, multicenter clinical trial. J. Altern. Complement. Med. 2019, 25, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Dugas, B.; Renauld, J.C.; Pène, J.; Bonnefoy, J.Y.; Peti-Frère, C.; Braquet, P.; Bousquet, J.; Van Snick, J.; Mencia-Huerta, J.M. Interleukin-9 potentiates the interleukin-4-induced immunoglobulin (IgG, IgM and IgE) production by normal human B lymphocytes. Eur. J. Immunol. 1993, 23, 1687–1692. [Google Scholar] [CrossRef]
- Huang, Z.P.; Zhang, L.N.; Duan, X.B.; Liao, Z.Q.; Ding, H.; Cheung, P.C.K. Novel highly branched water-soluble heteropolysaccharides as immunopotentiators to inhibit S-180 tumor cell growth in BALB/c mice. Carbohydr. Polym. 2012, 87, 427–434. [Google Scholar] [CrossRef]
- Annick, A.J.M.; Alfaro, T.M.; Robinson, A.N.; Baumann, U.; Hurst, J.R. Managing granulomatous-lymphocytic interstitial lung disease in common variable immunodeficiency disorders: E-GLILDnet international clinicians survey. Front. Immunol. 2020, 11, 606333. [Google Scholar]
- Erniati; Rungkat, F.Z.; Prangdimurti, E.; Adawiyah, D.R.; Priosoeryanto, B.P.; Huda, N. Stimulating activity on human lymphocytes in vitro of nori like product (Geluring) made from Gelidium sp. and Ulva lactuca seaweeds. Int. J. Adv. Sci. Eng. Inf. Technol. 2020, 10, 1311. [Google Scholar] [CrossRef]
- Gao, X.; Qu, H.; Gao, Z.; Zeng, D.; Wang, J.; Baranenko, D.; Li, Y.; Lu, W. Protective effects of Ulva pertusa polysaccharide and polysaccharide-iron (III) complex on cyclophosphamide induced immunosuppression in mice. Int. J. Biol. Macromol. 2019, 133, 911–919. [Google Scholar] [CrossRef]
- Yamamoto, I.; Maruyama, H.; Takahashi, M.; Komiyama, K. The effect of dietary or intraperitoneally injected seaweed preparations on the growth of sarcoma-180 cells subcutaneously implanted into mice. Cancer Lett. 1986, 30, 125–131. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Therho, T.T.; Hartiala, K. Method for determination of the sulfate content of glycosaminoglycans. Anal. Biochem. 1971, 41, 471–476. [Google Scholar] [CrossRef]
- Cesaretti, M. A 96-well assay for uronic acid carbazole reaction. Carbohydr. Polym. 2003, 54, 59–61. [Google Scholar] [CrossRef]
- Cao, S.J.; He, X.X.; Qin, L.; He, M.J.; Yang, Y.J.; Liu, Z.C.; Mao, W.J. Anticoagulant and antithrombotic properties in vitro and in vivo of a novel sulfated polysaccharide from marine green alga Monostroma nitidum. Mar. Drugs 2019, 17, 247. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Nakashima, T.; Ueda, T.; Tomii, K.; Kouno, I. Facile discrimination of aldose enantiomers by reversed-phase HPLC. Chem. Pharm. Bull. 2007, 55, 899–901. [Google Scholar] [CrossRef] [Green Version]
- Falshaw, R.; Furneaux, R.H. Structural analysis of carrageenans from the tetrasporic stages of the red algae, Gigartina lanceata and Gigartina chapmanii (Gigartinaceae, Rhodophyta). Carbohydr. Res. 1998, 307, 325–331. [Google Scholar] [CrossRef]
- Taylor, R.L.; Conrad, H.E. Stoichiometric depolymerization of polyuronides and glycosaminoglycuronans to monosaccharides following reduction of their carbodiimide-activated carboxyl group. Biochemistry 1972, 11, 1383–1388. [Google Scholar] [CrossRef]
- Sims, I.M.; Carnachan, S.M.; Bell, T.J.; Hinkley, S.F.R. Methylation analysis of polysaccharides: Technical advice. Carbohydr. Polym. 2018, 188, 1–7. [Google Scholar] [CrossRef]
- Shingel, K.I. Determination of structural peculiarities of dexran, pullulan and γ-irradiated pullulan by Fourier-Transform IR spectroscopy. Carbohydr. Res. 2002, 337, 1445–1451. [Google Scholar] [CrossRef]
- Yi, J.E.; Obminska-Mrukowicz, B.; Yuan, L.Y.; Yuan, H. Immunomodulatory effects of betulinic acid from the bark of white birch on mice. J. Vet. Sci. 2010, 11, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.H.; Cong, L.X.; Wang, C.M.; Li, H.; Zhang, C.Y.; Guan, X.G.; Liu, P.; Xie, Y.; Chen, J.G.; Sun, J.H. Immunomodulatory effect of schisandra polysaccharides in cyclophosphamide-induced immunocompromised Mice. Exp. Ther. Med. 2018, 6, 4755–4762. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.L.; Wei, S.Q.; Lu, X.M.; Qiao, X.G.; Simal-Gandara, J.; Capanoglu, E.; Woźniak, Ł.; Zou, L.; Cao, H.; Xiao, J.B.; et al. A neutral polysaccharide with a triple helix structure from ginger: Characterization and immunomodulatory activity. Food Chem. 2021, 350, 129261. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylated Alditol Acetate | Molar Percent Ratio | Linkage Pattern | ||
|---|---|---|---|---|
| UCP | DsUCP | RdUCP | ||
| 1,5-Di-O-acetyl-2,3,4-tri-O-methyl-rhamnitol | 2.46 | 2.56 | 1.94 | Rhap-(1→ |
| 1,5-Di-O-acetyl-2,3,4-tri-O-methyl-xylitol | 1.69 | 1.98 | 1.51 | Xylp-(1→ |
| 1,5-Di-O-acetyl-2,3,4,6-tetro-O-methyl-glucitol | -- | -- | 2.43 | Glcp-(1→ |
| 1,4,5-Tri-O-acetyl-2,3-di-O-methyl-rhamnitol | 20.31 | 59.68 | 15.37 | →4)-Rhap-(1→ |
| 1,4,5-Tri-O-acetyl-2,3-di-O-methyl-xylitol | 11.08 | 24.83 | 8.38 | →4)-Xylp-(1→ |
| 1,2,4,5-Tetra-O-acetyl-3-O-methyl-xylitol | 13.99 | -- | 10.59 | →2,4)-Xylp-(1→ |
| 1,2,4,5-Tetra-O-acetyl-3-O-methyl-rhamnitol | 10.07 | 10.94 | 7.02 | →2,4)-Rhap-(1→ |
| 1,3,4,5-Tetra-O-acetyl-2-O-methyl-rhamnitol | 40.38 | -- | 30.56 | →3,4)-Rhap-(1→ |
| 1,4,5-Tri-O-acetyl-2,3,6-tri-O-metyl-glucitol | -- | -- | 22.17 | →4)-Glcp-(1→ |
| Residues | Chemical Shifts (ppm) | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| A →2,4)-α-l-Rhap-(1→ | 1H | 5.31 | 4.44 | 4.19 | 3.83 | 4.17 | 1.35 |
| 13C | 102.59 | 79.80 | 70.41 | 79.59 | 69.61 | 18.69 | |
| B →4)-α-l-Rhap(3SO4)-(1→ | 1H | 5.12 | 4.26 | 4.67 | 3.84 | 4.05 | 1.35 |
| 13C | 103.33 | 70.52 | 79.80 | 79.49 | 69.61 | 18.69 | |
| C →4)-β-d-Xylp-(2SO4)-(1→ | 1H | 5.02 | 4.16 | 3.90 | 3.70 | 3.52 | |
| 13C | 101.41 | 79.49 | 72.72 | 75.34 | 62.75 | ||
| D →4)-β-l-Rhap(3SO4)-(1→ | 1H | 4.95 | 4.30 | 4.62 | 3.80 | 3.63 | 1.35 |
| 13C | 99.77 | 70.51 | 79.80 | 79.49 | 71.62 | 18.69 | |
| E →4)-β-l-Rhap-(1→ | 1H | 4.91 | 4.21 | 4.00 | 3.77 | 3.60 | 1.35 |
| 13C | 98.93 | 70.4 | 69.59 | 79.49 | 71.62 | 18.69 | |
| F →4)-β-d-GlcAp-(1→ | 1H | 4.68 | 3.38 | 3.68 | 3.64 | 3.82 | |
| 13C | 104.67 | 75.13 | 75.16 | 80.29 | 77.34 | 176.46 | |
| G →4)-β-d-Xylp-(1→ | 1H | 4.55 | 3.36 | 3.67 | 3.68 | 3.38/4.17 | |
| 13C | 105.18 | 75.03 | 75.43 | 77.56 | 63.92 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, S.; Yang, Y.; Liu, S.; Shao, Z.; Chu, X.; Mao, W. Immunomodulatory Activity In Vitro and In Vivo of a Sulfated Polysaccharide with Novel Structure from the Green Alga Ulvaconglobata Kjellman. Mar. Drugs 2022, 20, 447. https://doi.org/10.3390/md20070447
Cao S, Yang Y, Liu S, Shao Z, Chu X, Mao W. Immunomodulatory Activity In Vitro and In Vivo of a Sulfated Polysaccharide with Novel Structure from the Green Alga Ulvaconglobata Kjellman. Marine Drugs. 2022; 20(7):447. https://doi.org/10.3390/md20070447
Chicago/Turabian StyleCao, Sujian, Yajing Yang, Shan Liu, Zhuling Shao, Xiao Chu, and Wenjun Mao. 2022. "Immunomodulatory Activity In Vitro and In Vivo of a Sulfated Polysaccharide with Novel Structure from the Green Alga Ulvaconglobata Kjellman" Marine Drugs 20, no. 7: 447. https://doi.org/10.3390/md20070447