

Anticancer Activity of Mannose-Specific Lectin, BPL2, from Marine Green Alga Bryopsis plumosa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
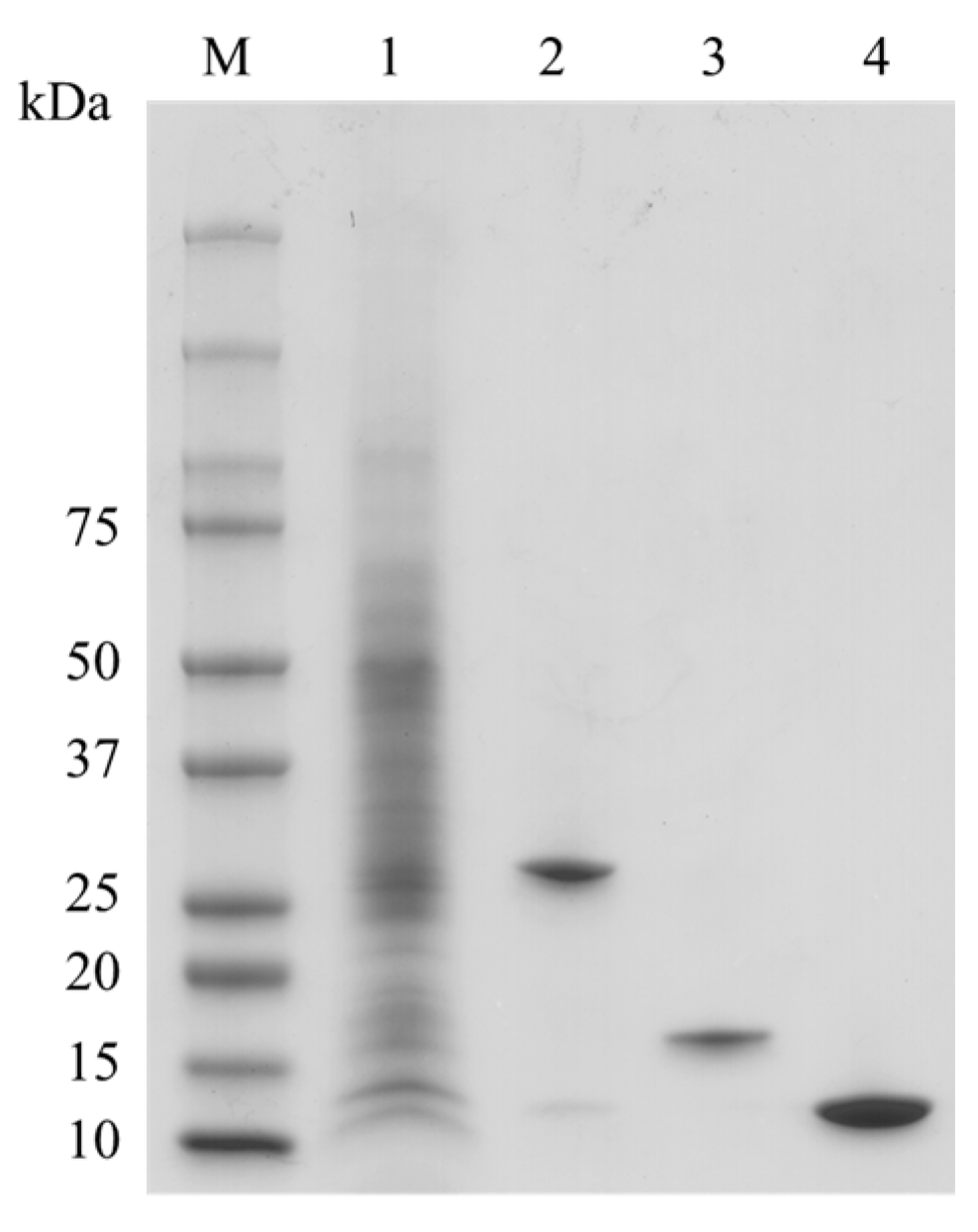
2.1. Preaparation of Bryopsis Plumosa Lectins
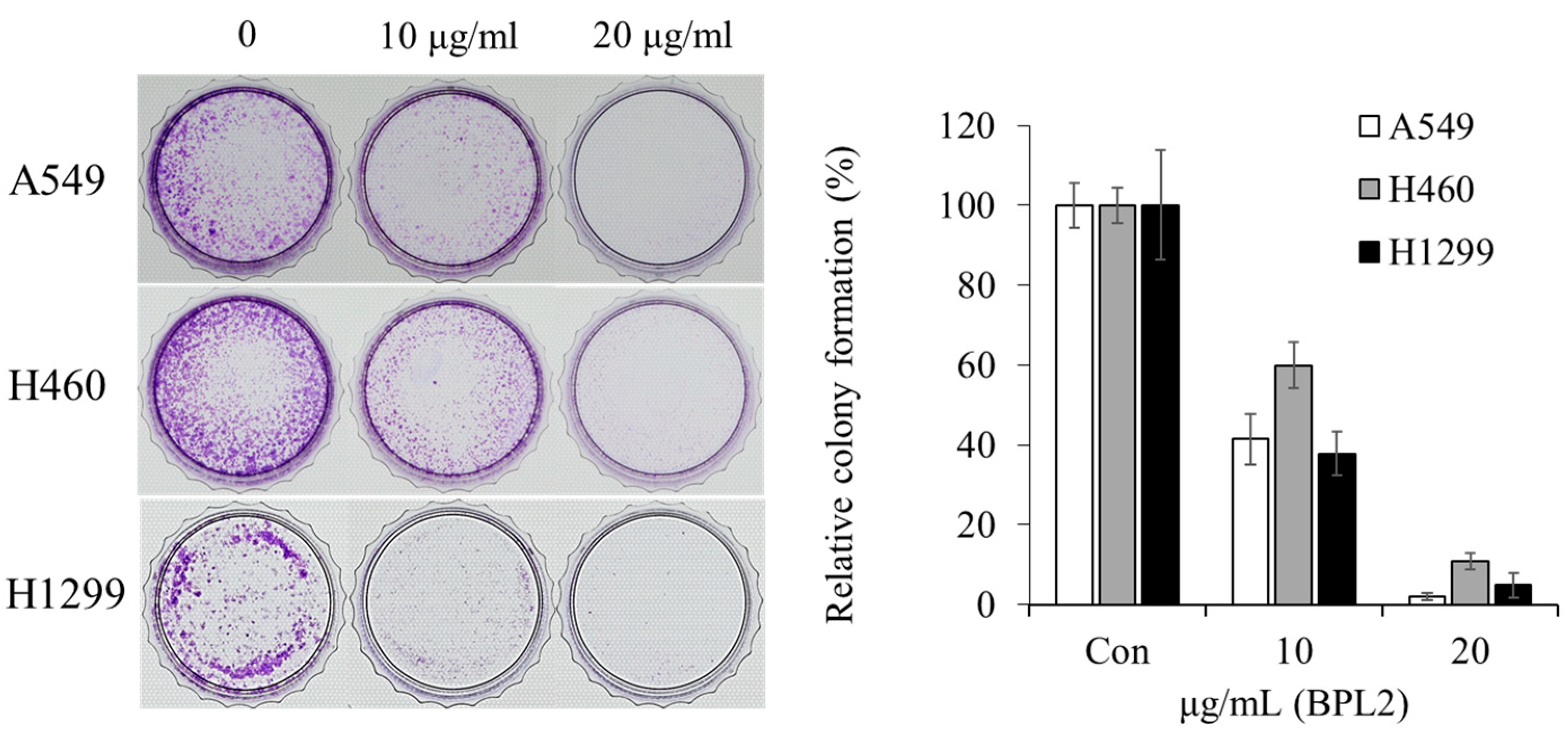
2.2. Cell Viability of Lung Cancer Cells and Non-Cancerous Cells against BPLs
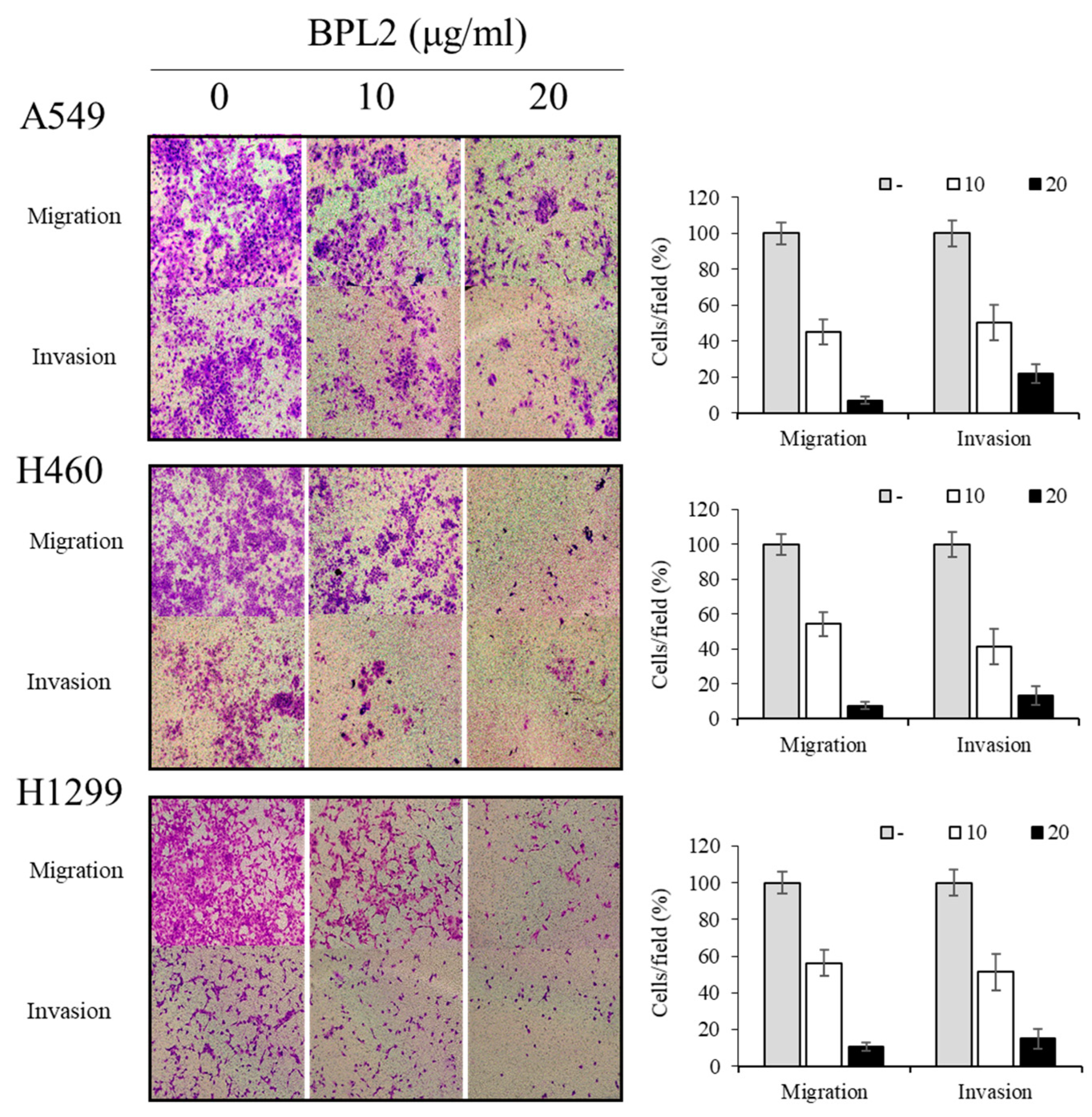
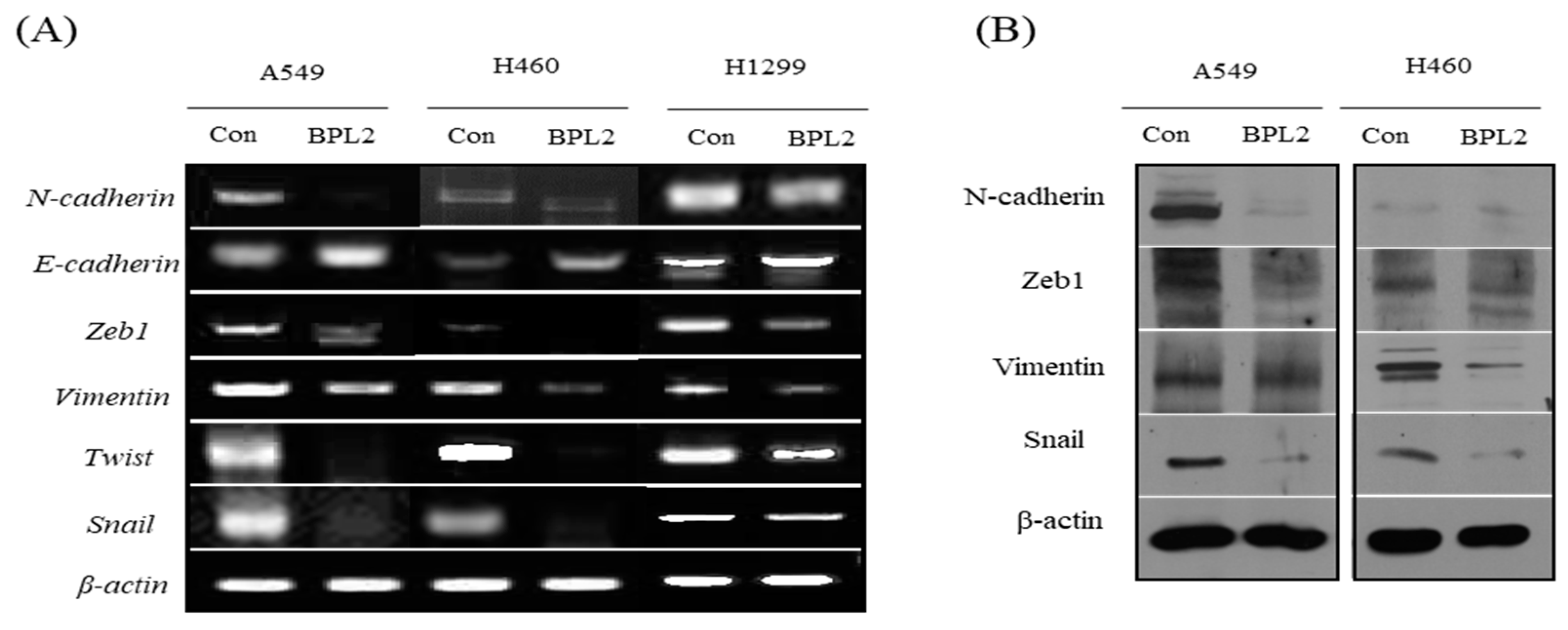
2.3. Determination of Migration and Invasion of Cancer Cells following BPL2 Treatment
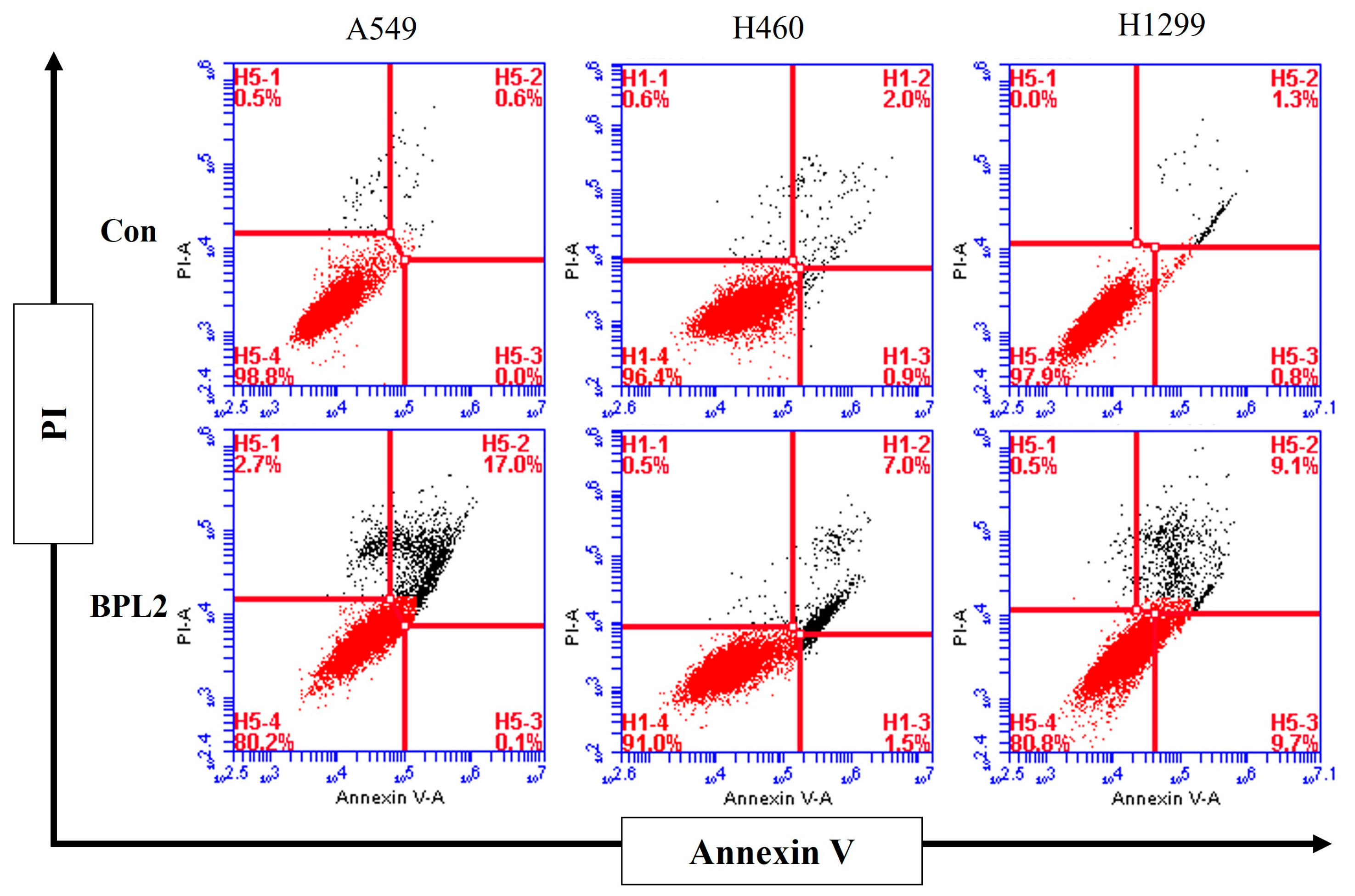
2.4. Analysis for Apoptosis in BPL2-Treated Lung Cancer Cell Lines
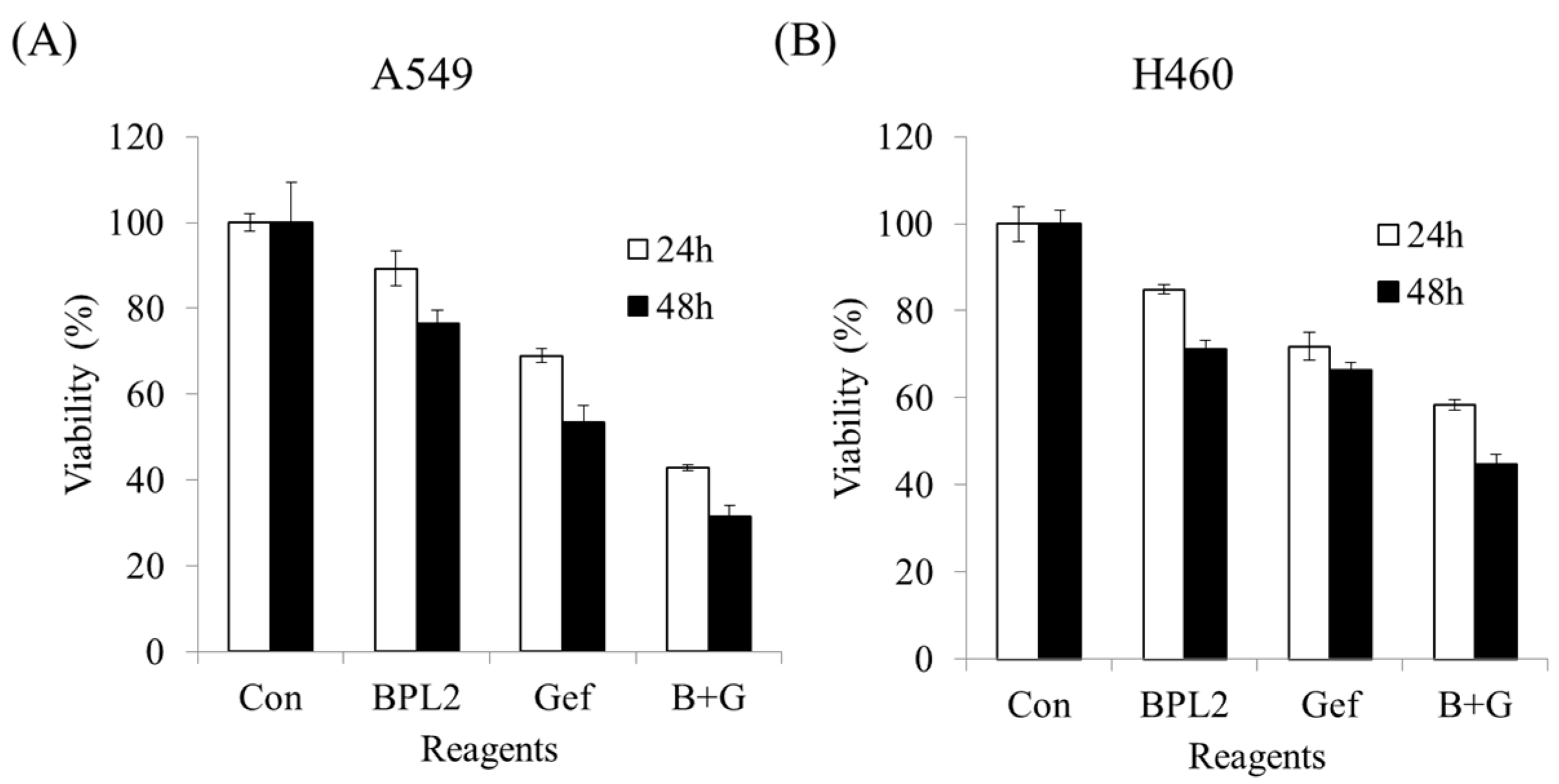
2.5. Effect of Concomitant Treatment with Gefitinib and BPL2 on Cell Viability
3. Discussion
4. Materials and Methods
4.1. Preparation of Bryopsis plumosa Lectins (BPLs)
4.2. Cell Culture and Viability Assay (Determination of Viability of Tumo Cells)
4.3. Determination of Cell Growth Rate Based on BPL Treatments
4.4. Determination of Migration and Invasion of Cancer Cells
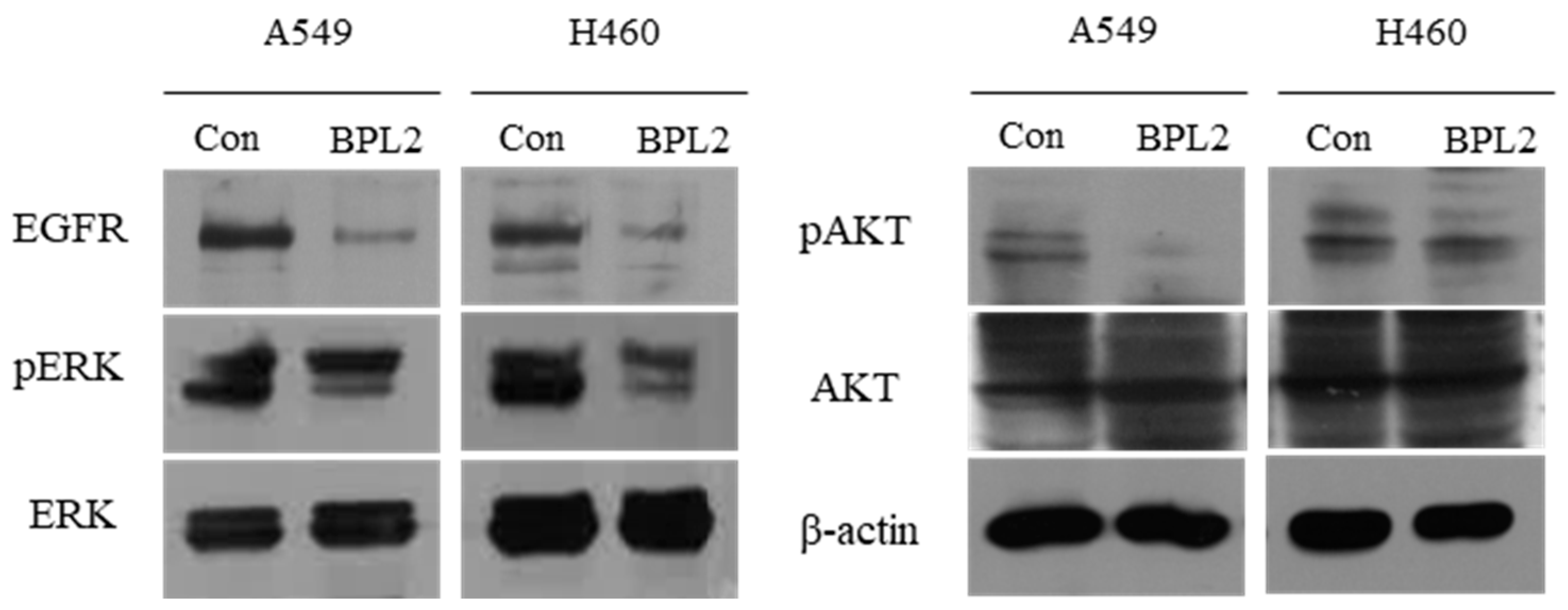
4.5. Comparison of the Expression Level of Cell Migration and Invasion-Related Genes, and EGFR-Related Proteins
4.6. The Effect of Concurrent Treatment (Gefitinib and BPL2) on Lung Cancer Cell Viability
4.7. Flow Cytometric Analysis for Apoptosis in BPL2-Treated Lung Cancer Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sharon, N.; Lis, H. Lectins: Cell-agglutinating and sugar-specific proteins. Science 1972, 177, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Nabi-Afjadi, M.; Heydari, M.; Zalpoor, H.; Arman, I.; Sadoughi, A.; Sahami, P.; Aghazadeh, S. Lectins and lectibodies: Potential promising antiviral agents. Cell. Mol. Biol. Lett. 2022, 27, 37. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Walia, A.K. Lectins from red algae and their biomedical potential. J. Appl. Phycol. 2018, 30, 1833–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Jung, M.; Shim, E.; Shim, J.; Kim, Y.; Kim, G. Functional recombinants designed from a fetuin/asialofetuin-specific marine algal lectin, Rhodobindin. Mar. Drugs 2015, 13, 2183–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Attarwala, H. Role of antibodies in cancer targeting. J. Nat. Sci. Biol. Med. 2010, 1, 53. [Google Scholar] [CrossRef] [Green Version]
- Seca, A.; Pinto, D. Plant secondary metabolites as anticancer agents: Successes in clinical trials and therapeutic application. Int. J. Mol. Sci. 2018, 19, 263. [Google Scholar] [CrossRef] [Green Version]
- Karpiński, T.; Adamczak, A. Anticancer activity of bacterial proteins and peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Gewirtz, D.A.; Bristol, M.L.; Yalowich, J.C. Toxicity issues in cancer drug development. Curr. Opin. Investig. Drugs 2010, 11, 612–614. [Google Scholar]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Rodrigues, J.G.; Balmaña, M.; Macedo, J.A.; Poças, J.; Fernandes, Â.; De-Freitas-Junior, J.C.M.; Pinho, S.S.; Gomes, J.; Magalhães, A.; Gomes, C.; et al. Glycosylation in cancer: Selected roles in tumour progression, immune modulation and metastasis. Cell. Immunol. 2018, 333, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yu, Y.; Wang, Y.; Zhu, B.; Han, B. COLGALT1 is a potential biomarker for predicting prognosis and immune responses for kidney renal clear cell carcinoma and its mechanisms of ceRNA networks. Eur. J. Med. Res. 2022, 27, 122. [Google Scholar] [CrossRef]
- Rodrigues Mantuano, N.; Natoli, M.; Zippelius, A.; Läubli, H. Tumor-associated carbohydrates and immunomodulatory lectins as targets for cancer immunotherapy. J. Immunother. Cancer 2020, 8, e001222. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.; Dan, X.; Ng, C.; Ng, T. Lectins with Potential for Anti-Cancer Therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Chen, J.; Li, C.; An, N.; Wang, Z.; Yang, S.; Huang, K.; Bao, J. Antitumor effects of concanavalin A and Sophora flavescens lectin in vitro and in vivo. Acta Pharmacol. Sin. 2014, 35, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, C.; Das, M.; Stephen-Victor, E.; Friboulet, A.; Bayry, J.; Kaveri, S. Differential effects of Viscum album preparations on the maturation and activation of human dendritic cells and CD4+ T cell responses. Molecules 2016, 21, 912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twardziok, M.; Kleinsimon, S.; Rolff, J.; Jäger, S.; Eggert, A.; Seifert, G.; Delebinski, C.I. Multiple active compounds from Viscum album L. synergistically converge to promote apoptosis in Ewing sarcoma. PLoS ONE 2016, 11, e0159749. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.-M.; Cheng, L.; Guo, S.-J.; Wang, Y.; Czajkowsky, D.M.; Gao, H.; Hu, X.-F.; Tao, S.-C. Lectin RCA-I specifically binds to metastasis-associated cell surface glycans in triple-negative breast cancer. Breast Cancer Res. 2015, 17, 36. [Google Scholar] [CrossRef] [Green Version]
- Maliki, I.M.; Misson, M.; Teoh, P.L.; Rodrigues, K.F.; Yong, W.T.L. Production of lectins from marine algae: Current status, challenges, and opportunities for non-destructive extraction. Mar. Drugs 2022, 20, 102. [Google Scholar] [CrossRef]
- Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A novel high-mannose specific lectin from the green alga Halimeda renschii exhibits a potent anti-influenza virus activity through high-affinity binding to the viral hemagglutinin. Mar. Drugs 2017, 15, 255. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.S.; Thakur, S.R.; Bansal, P. Algal lectins as promising biomolecules for biomedical research. Crit. Rev. Microbiol. 2015, 41, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Klochkova, T.A.; Yoon, K.-S.; Song, Y.-S.; Lee, K.P. Purification and characterization of a lectin, bryohealin, involved in the protoplast formation of a marine green alga Bryopsis plumosa (Chlorophyta) 1. J. Phycol. 2006, 42, 86–95. [Google Scholar] [CrossRef]
- Han, J.W.; Yoon, K.S.; Klochkova, T.A.; Hwang, M.S.; Kim, G.H. Purification and characterization of a lectin, BPL-3, from the marine green alga Bryopsis plumosa. J. Appl. Phycol. 2011, 23, 745–753. [Google Scholar] [CrossRef]
- Han, J.-W.; Yoon, K.-S.; Jung, M.-G.; Chah, K.-H.; Kim, G.-H. Molecular characterization of a lectin, BPL-4, from the marine green alga Bryopsis plumosa (Chlorophyta). Algae 2012, 27, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Han, J.W.; Jung, M.G.; Kim, M.J.; Yoon, K.S.; Lee, K.P.; Kim, G.H. Purification and characterization of a D-mannose specific lectin from the green marine alga, Bryopsis plumosa. Phycol. Res. 2010, 58, 143–150. [Google Scholar] [CrossRef]
- Jiang, Q.-L.; Zhang, S.; Tian, M.; Zhang, S.-Y.; Xie, T.; Chen, D.-Y.; Chen, Y.-J.; He, J.; Liu, J.; Ouyang, L.; et al. Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell Prolif. 2015, 48, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yin, D.; Wang, W.; Shen, X.; Zhu, J.-J.; Chen, H.-Y.; Zhen, L. Targeting and imaging of cancer cells via monosaccharide-imprinted fluorescent nanoparticles. Sci. Rep. 2016, 6, 22757. [Google Scholar] [CrossRef] [Green Version]
- Shan, M.; Yang, D.; Dou, H.; Zhang, L. Fucosylation in cancer biology and its clinical applications. Prog. Mol. Biol. Transl. Sci. 2019, 162, 93–119. [Google Scholar]
- Barre, A.; Simplicien, M.; Benoist, H.; Van Damme, E.J.; Rougé, P. Mannose-specific lectins from marine algae: Diverse structural scaffolds associated to common virucidal and anti-cancer properties. Mar. Drugs 2019, 17, 440. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.-J.; Han, J.-W.; Kim, G.H.; Han, J.W. Functional expression and characterization of the recombinant N-acetyl-glucosamine/N-acetyl-galactosamine-specific marine algal lectin BPL3. Mar. Drugs 2018, 16, 13. [Google Scholar] [CrossRef] [Green Version]
- Tian, R.; Zhang, H.; Chen, H.; Liu, G.; Wang, Z. Uncovering the binding specificities of lectins with cells for precision colorectal cancer diagnosis based on multimodal imaging. Adv. Sci. 2018, 5, 1800214. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.D.; Trinh, P.T.H. Structure and anticancer activity of a new lectin from the cultivated red alga, Kappaphycus striatus. J. Nat. Med. 2021, 75, 223–231. [Google Scholar] [CrossRef]
- Sindhura, B.R.; Hegde, P.; Chachadi, V.B.; Inamdar, S.R.; Swamy, B.M. High mannose N-glycan binding lectin from Remusatia vivipara (RVL) limits cell growth, motility and invasiveness of human breast cancer cells. Biomed. Pharmacother. 2017, 93, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Panda, P.K.; Mukhopadhyay, S.; Behera, B.; Bhol, C.S.; Dey, S.; Das, D.N.; Sinha, N.; Bissoyi, A.; Pramanik, K.; Maiti, T.K.; et al. Antitumor effect of soybean lectin mediated through reactive oxygen species-dependent pathway. Life Sci. 2014, 111, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, C.; Bian, H.; Min, M.; Chen, L.; Bao, J. Antiproliferative activity and apoptosis-inducing mechanism of Concanavalin A on human melanoma A375 cells. Arch. Biochem. Biophys. 2009, 482, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Christofori, G.; Lehembre, F. Distinct mechanisms of tumor invasion and metastasis. Trends Mol. Med. 2007, 13, 535–541. [Google Scholar] [CrossRef]
- Jian, Q.; Yang, Z.; Shu, J.; Liu, X.; Zhang, J.; Li, Z. Lectin BS-I inhibits cell migration and invasion via AKT/GSK-3β/β-catenin pathway in hepatocellular carcinoma. J. Cell. Mol. Med. 2018, 22, 315–329. [Google Scholar] [CrossRef]
- Song, N.; Zhong, J.; Hu, Q.; Gu, T.; Yang, B.; Zhang, J.; Yu, J.; Ma, X.; Chen, Q.; Qi, J.; et al. FGF18 enhances migration and the epithelial-mesenchymal transition in breast cancer by regulating Akt/GSK3β/Β-catenin signaling. Cell. Physiol. Biochem. 2018, 49, 1060–1073. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, Z.; Zhou, C.; Liu, L.; Huang, C. Epithelial–mesenchymal transition: The history, regulatory mechanism, and cancer therapeutic opportunities. MedComm 2022, 3, e144. [Google Scholar] [CrossRef]
- Jung, A.R.; Jung, C.-H.; Noh, J.K.; Lee, Y.C.; Eun, Y.-G. Epithelial-mesenchymal transition gene signature is associated with prognosis and tumor microenvironment in head and neck squamous cell carcinoma. Sci. Rep. 2020, 10, 3652. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.E.; Nathan, V.; Osborne, J.K.; Farrow, R.K.; Deb, D.; Sullivan, J.P.; Dospoy, P.D.; Augustyn, A.; Hight, S.K.; Sato, M.; et al. ZEB1 drives epithelial-to-mesenchymal transition in lung cancer. J. Clin. Investig. 2016, 126, 3219–3235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drápela, S.; Bouchal, J.; Jolly, M.K.; Culig, Z.; Souček, K. ZEB1: A Critical Regulator of Cell Plasticity, DNA Damage Response, and Therapy Resistance. Front. Mol. Biosci. 2020, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Sun, Y.; Ma, L. ZEB1: At the crossroads of epithelial-mesenchymal transition, metastasis and therapy resistance. Cell Cycle 2015, 14, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Fu, R.; Li, Y.; Jiang, N.; Ren, B.-X.; Zang, C.-Z.; Liu, L.-J.; Lv, W.-C.; Li, H.-M.; Weiss, S.; Li, Z.-Y.; et al. Inactivation of endothelial ZEB1 impedes tumor progression and sensitizes tumors to conventional therapies. J. Clin. Investig. 2020, 130, 1252–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Chen, H.; Zhang, D.; Fu, J. Twist: A molecular target in cancer therapeutics. Tumor Biol. 2013, 34, 2497–2506. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Wang, Y.; Zhuo, X.; Zhang, Y.; Ao, X.; Chen, Z. Knockdown of Snail, a novel zinc finger transcription factor, via RNA interference increases A549 cell sensitivity to cisplatin via JNK/mitochondrial pathway. Lung Cancer 2008, 62, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Dou, J.; He, X.; Liu, Y.; Wang, Y.; Zhao, F.; Wang, X.; Chen, D.; Shi, F.; Wang, J. Effect of downregulation of ZEB1 on vimentin expression, tumour migration and tumourigenicity of melanoma B16F10 cells and CSCs. Cell Biol. Int. 2014, 38, 452–461. [Google Scholar] [CrossRef]
- Caramel, J.; Ligier, M.; Puisieux, A. Pleiotropic roles for ZEB1 in cancer. Cancer Res. 2018, 78, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Fry, E.A. Novel Molecular Markers for Breast Cancer. Biomark. Cancer 2016, 8, BIC.S38394. [Google Scholar] [CrossRef]
- Loh, C.-Y.; Chai, J.; Tang, T.; Wong, W.; Sethi, G.; Shanmugam, M.; Chong, P.; Looi, C. The E-cadherin and N-cadherin switch in epithelial-to-mesenchymal transition: Signaling, therapeutic implications, and challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Mo, C.; Peng, Q.; Kang, X.; Sun, C.; Jiang, K.; Huang, L.; Lu, Y.; Sui, J.; Qin, X.; et al. Cell surface glycan alterations in epithelial mesenchymal transition process of Huh7 hepatocellular carcinoma cell. PLoS ONE 2013, 8, e71273. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xie, X.; Wang, P.; Luo, J.; Chen, Y.; Xu, Q.; Zhou, J.; Lu, X.; Zhao, J.; Chen, Z.; et al. Mannan-binding lectin reduces epithelial-mesenchymal transition in pulmonary fibrosis via inactivating the store-operated calcium entry machinery. J. Innate Immun. 2022, 1–13. [Google Scholar] [CrossRef]
- Cho, S.; Yee, J.; Kim, J.Y.; Jeong Rhie, S.; Gwak, H.S. Effects of concomitant medication use on gefitinib-induced hepatotoxicity. J. Clin. Pharmacol. 2018, 58, 263–268. [Google Scholar] [CrossRef]
- Sato, Y.; Kubo, T.; Morimoto, K.; Yanagihara, K.; Seyama, T. High mannose-binding Pseudomonas fluorescens lectin (PFL) downregulates cell surface integrin/EGFR and induces autophagy in gastric cancer cells. BMC Cancer 2016, 16, 63. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, L.; Chen, Y.; Wang, X.Y.; Lu, R.F.; Zhang, S.Y.; Tian, M.; Xie, T.; Liu, B.; He, G. Polygonatum odoratum lectin induces apoptosis and autophagy via targeting EGFR-mediated Ras-Raf-MEK-ERK pathway in human MCF-7 breast cancer cells. Phytomedicine 2014, 21, 1658–1665. [Google Scholar] [CrossRef]
- Wu, M.; Yuan, Y.; Pan, Y.-Y.; Zhang, Y. Antitumor activity of combination treatment with gefitinib and docetaxel in EGFR-TKI-sensitive, primary resistant and acquired resistant human non-small cell lung cancer cells. Mol. Med. Rep. 2014, 9, 2417–2422. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Chen, J.; Lu, B.; Shi, Z.; Wang, H.; Zhang, B.; Zhao, K.; Qi, W.; Bao, J.; Wang, Y. Molecular switch role of Akt in Polygonatum odoratum lectin-induced apoptosis and autophagy in human non-small cell lung cancer A549 cells. PLoS ONE. 2014, 9, e101526. [Google Scholar] [CrossRef]
- Jotte, R.M.; Spigel, D.R. Advances in molecular-based personalized non-small-cell lung cancer therapy: Targeting epidermal growth factor receptor and mechanisms of resistance. Cancer Med. 2015, 4, 1621–1632. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.Q.; Yu, Z.Y.; Li, J.; Ouyang, X.N. Gefitinib induces lung cancer cell autophagy and apoptosis via blockade of the PI3K/AKT/mTOR pathway. Oncol Lett. 2016, 12, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Khil, L.Y.; Kim, W.; Lyu, S.; Park, W.B.; Yoon, J.W.; Jun, H.S. Mechanisms involved in Korean mistletoe lectin-induced apoptosis of cancer cells. World J. Gastroenterol. 2007, 13, 2811–2818. [Google Scholar] [CrossRef]
- Gondim, A.C.S.; Romero-Canelón, I.; Sousa, E.H.S.; Blindauer, C.A.; Butler, J.S.; Romero, M.J.; Sanchez-Cano, C.; Sousa, B.L.; Chaves, R.P.; Nagano, C.S.; et al. The potent anti-cancer activity of Dioclea lasiocarpa lectin. J. Inorg. Biochem. 2017, 175, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, B.; Zhang, Z.T.; Zhou, T.T.; Bian, H.J.; Min, M.W.; Liu, Y.H.; Chen, J.; Bao, J.K. A mannose-binding lectin from Sophora flavescens induces apoptosis in HeLa cells. Phytomedicine 2008, 15, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.E.; Park, A.K.; Lyu, S.Y. Synergistic anticancer effects of lectin and doxorubicin in breast cancer cells. Mol. Cell. Biochem. 2014, 394, 225–235. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Lee, S.B.; Kim, H.; Shin, J.M.; Yoon, M.; An, H.S.; Han, J.W. Anticancer Activity of Mannose-Specific Lectin, BPL2, from Marine Green Alga Bryopsis plumosa. Mar. Drugs 2022, 20, 776. https://doi.org/10.3390/md20120776
Lee JH, Lee SB, Kim H, Shin JM, Yoon M, An HS, Han JW. Anticancer Activity of Mannose-Specific Lectin, BPL2, from Marine Green Alga Bryopsis plumosa. Marine Drugs. 2022; 20(12):776. https://doi.org/10.3390/md20120776
Chicago/Turabian StyleLee, Jei Ha, Set Byul Lee, Heabin Kim, Jae Min Shin, Moongeun Yoon, Hye Suck An, and Jong Won Han. 2022. "Anticancer Activity of Mannose-Specific Lectin, BPL2, from Marine Green Alga Bryopsis plumosa" Marine Drugs 20, no. 12: 776. https://doi.org/10.3390/md20120776