


Carbon Source Influences Antioxidant, Antiglycemic, and Antilipidemic Activities of Haloferax mediterranei Carotenoid Extracts

Abstract
:1. Introduction
2. Results and Discussion
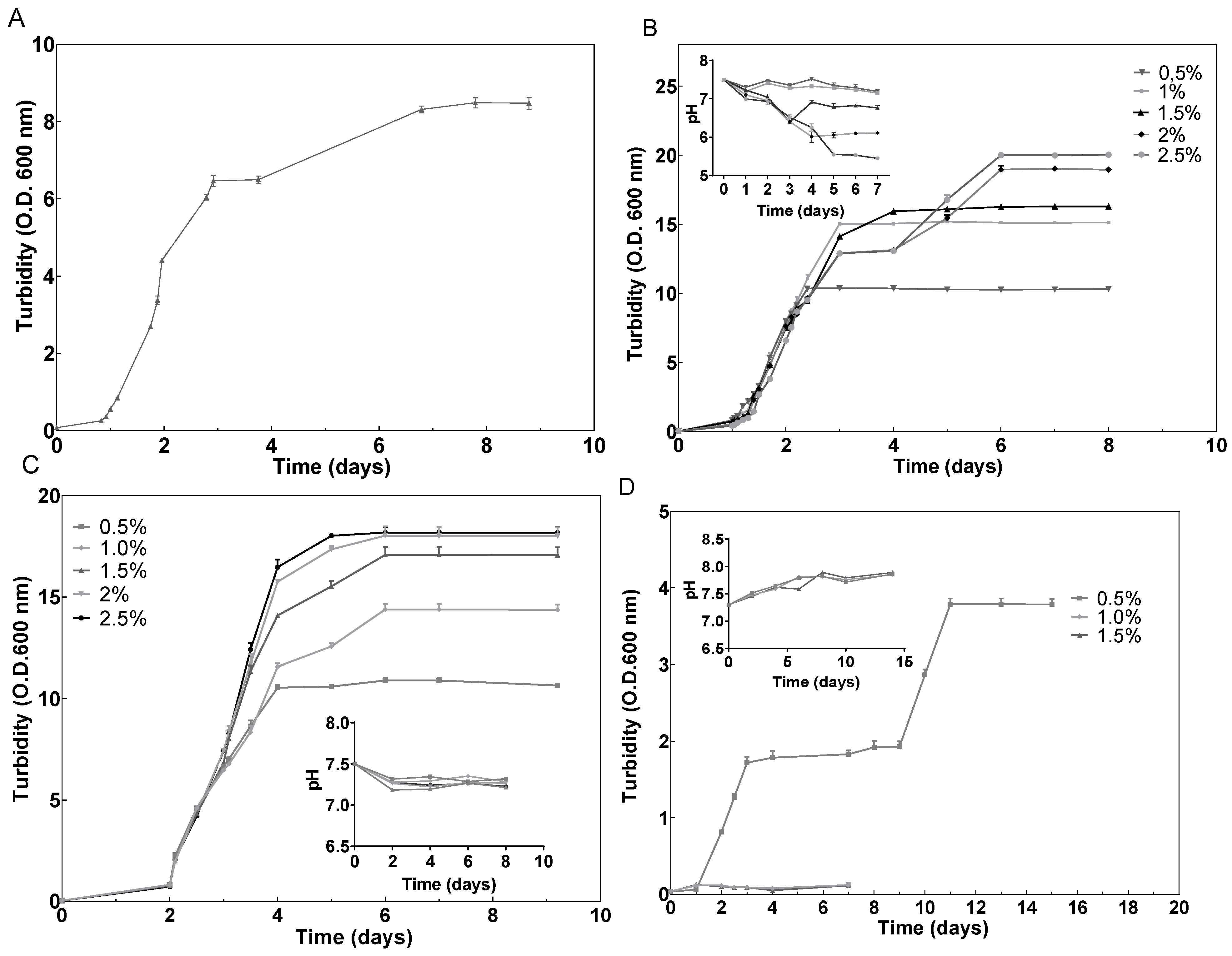
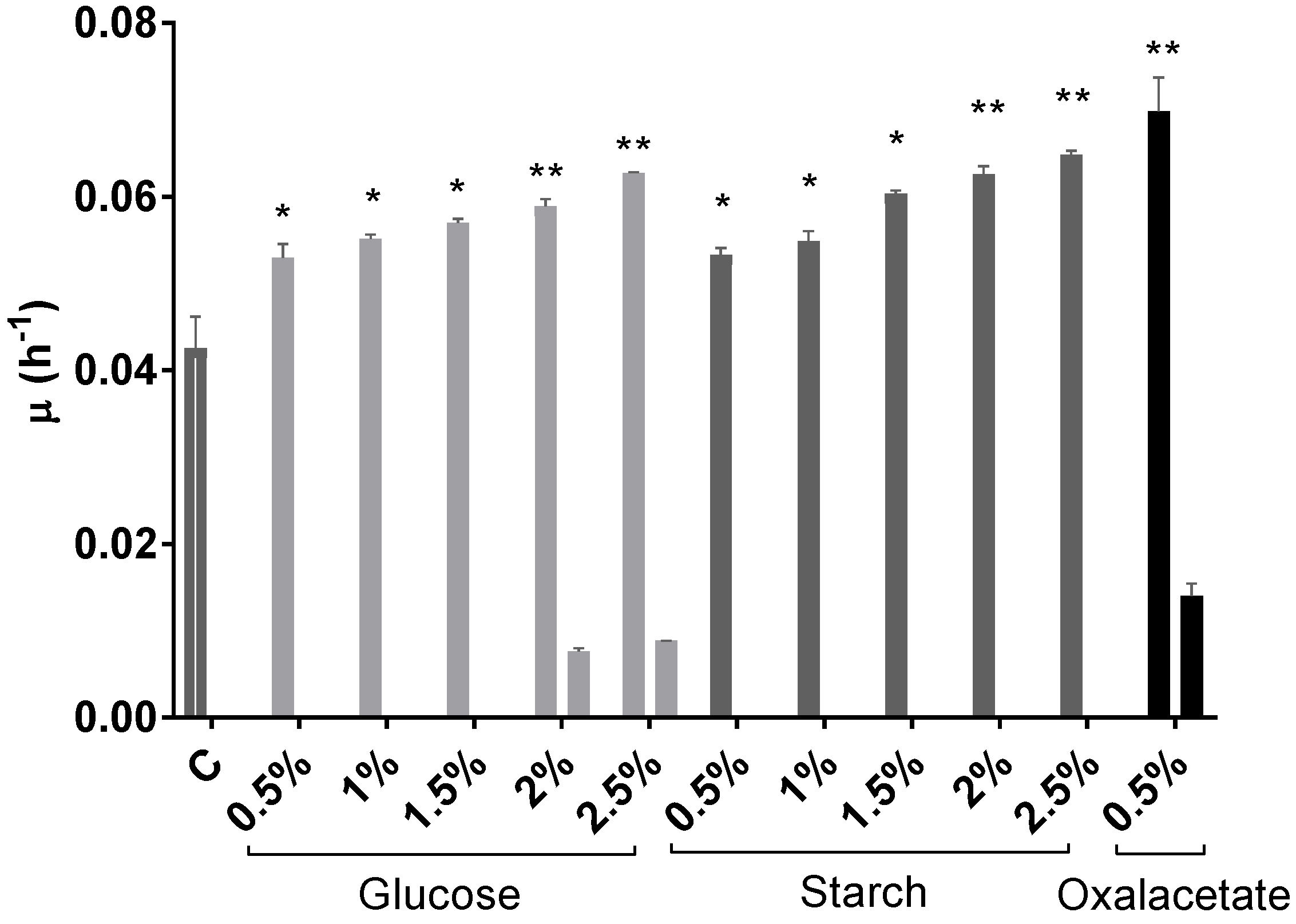
2.1. Effect of Different Carbon Sources on Growth
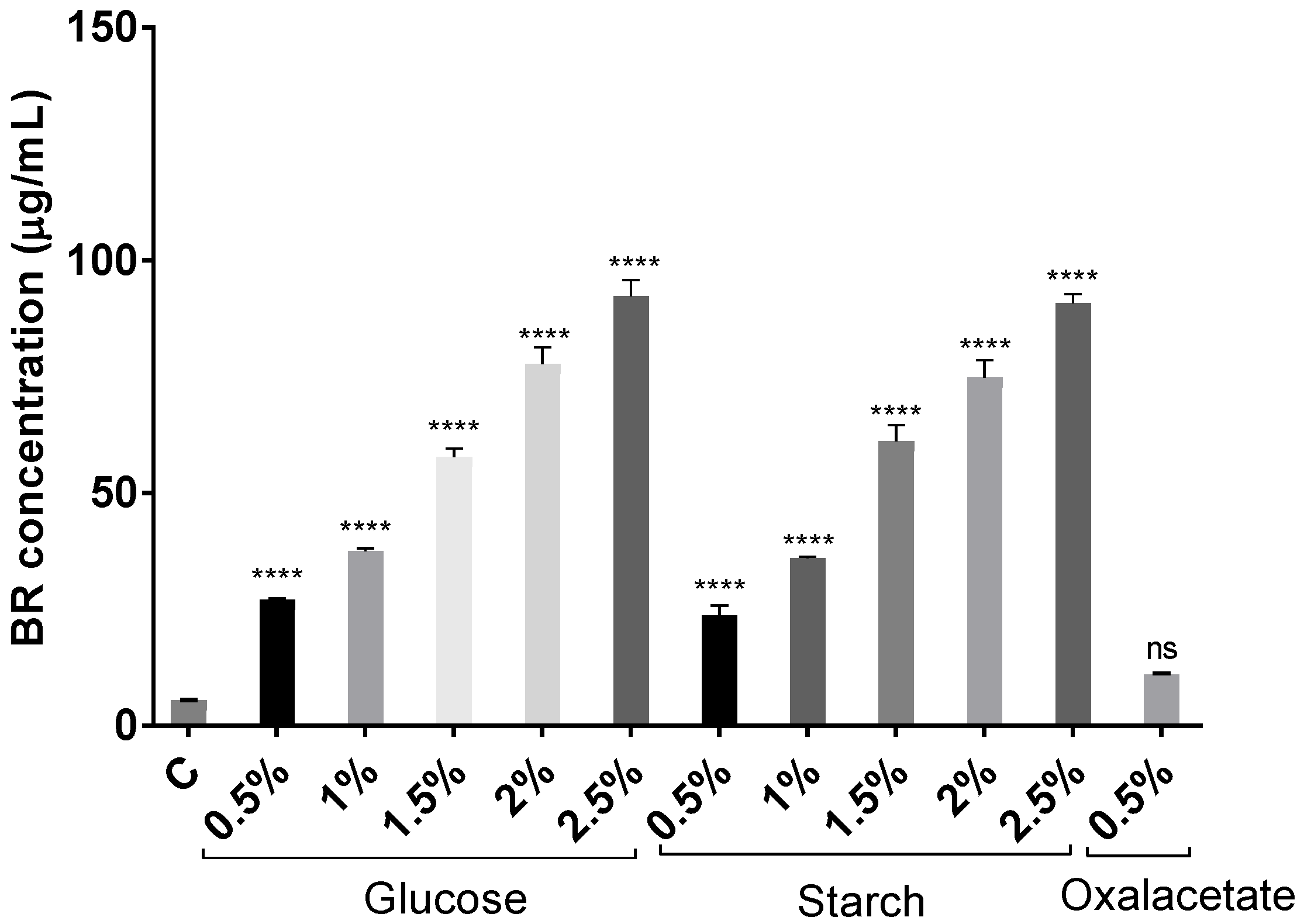
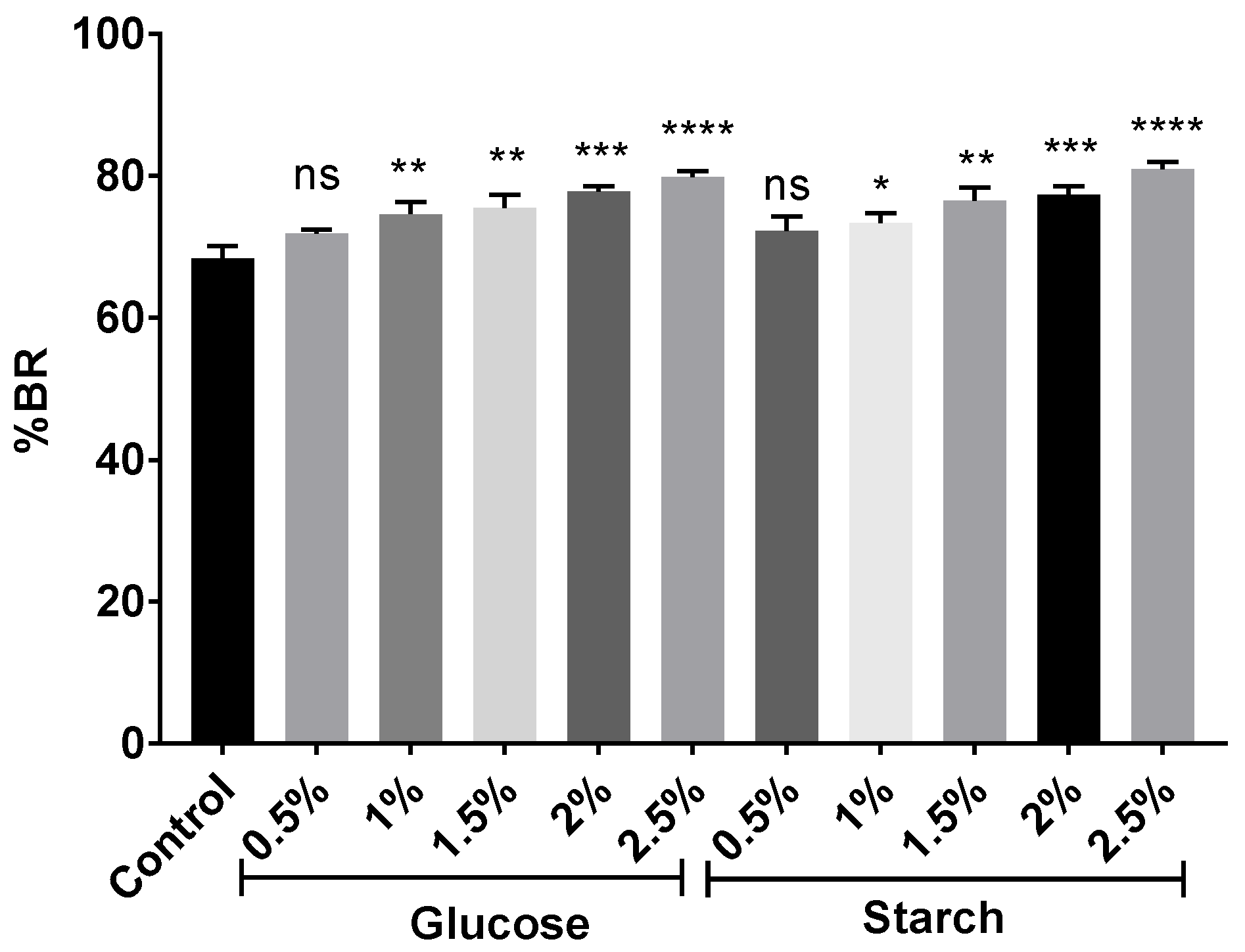
2.2. Effect of Different Carbon Sources on BR Production
2.3. Antioxidant Activity
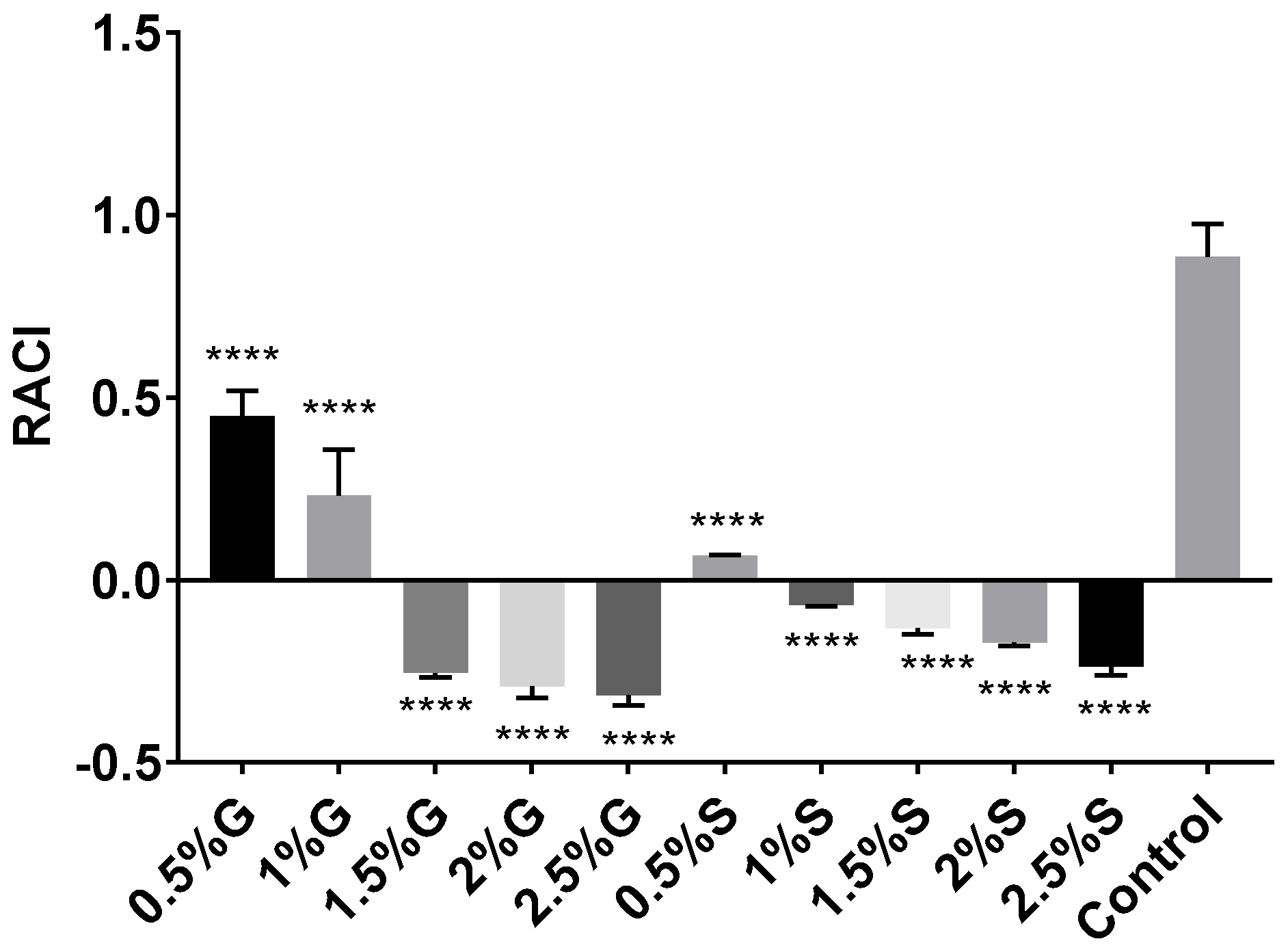
2.4. Antiglycemic and Antilipidemic Activity
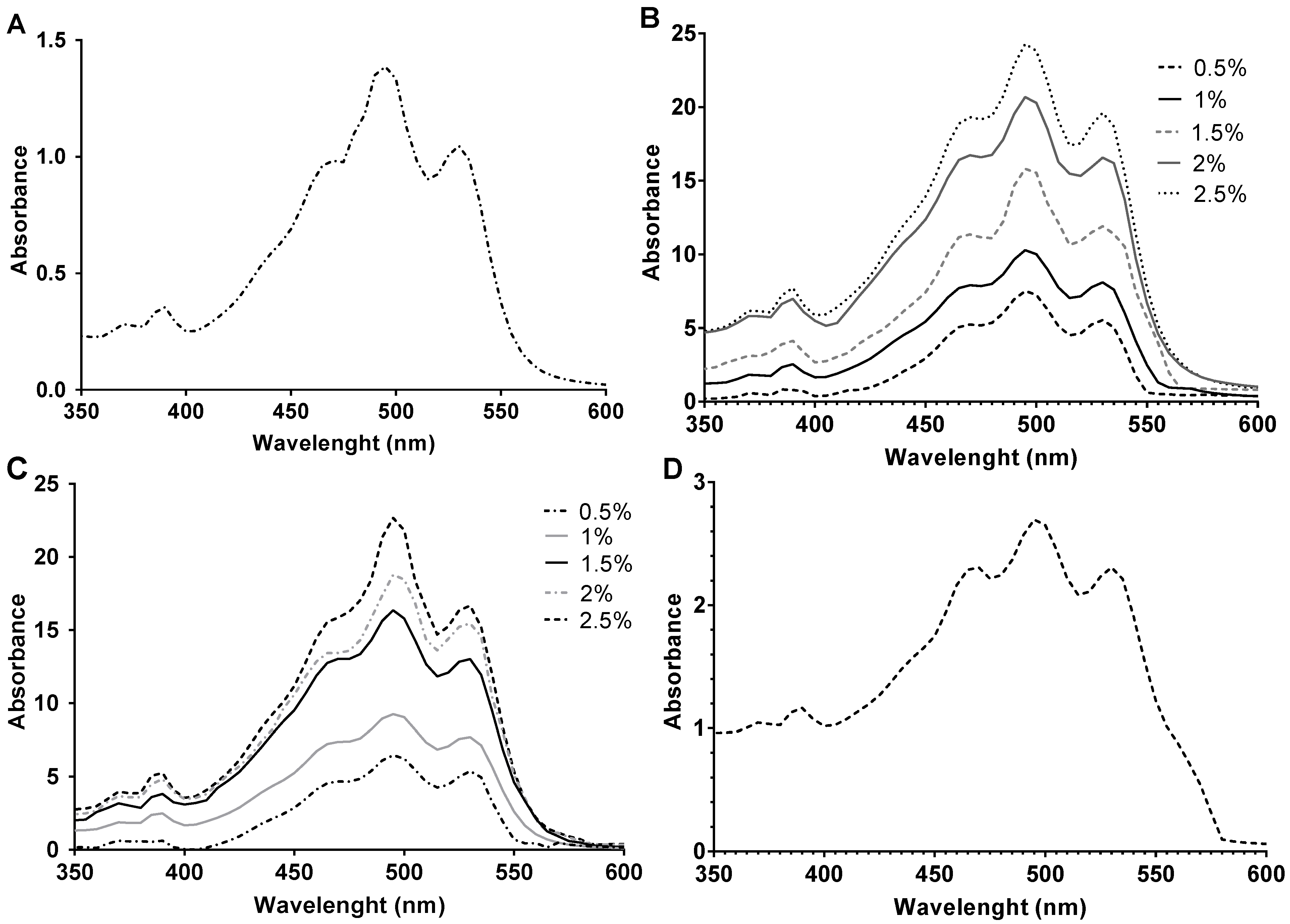
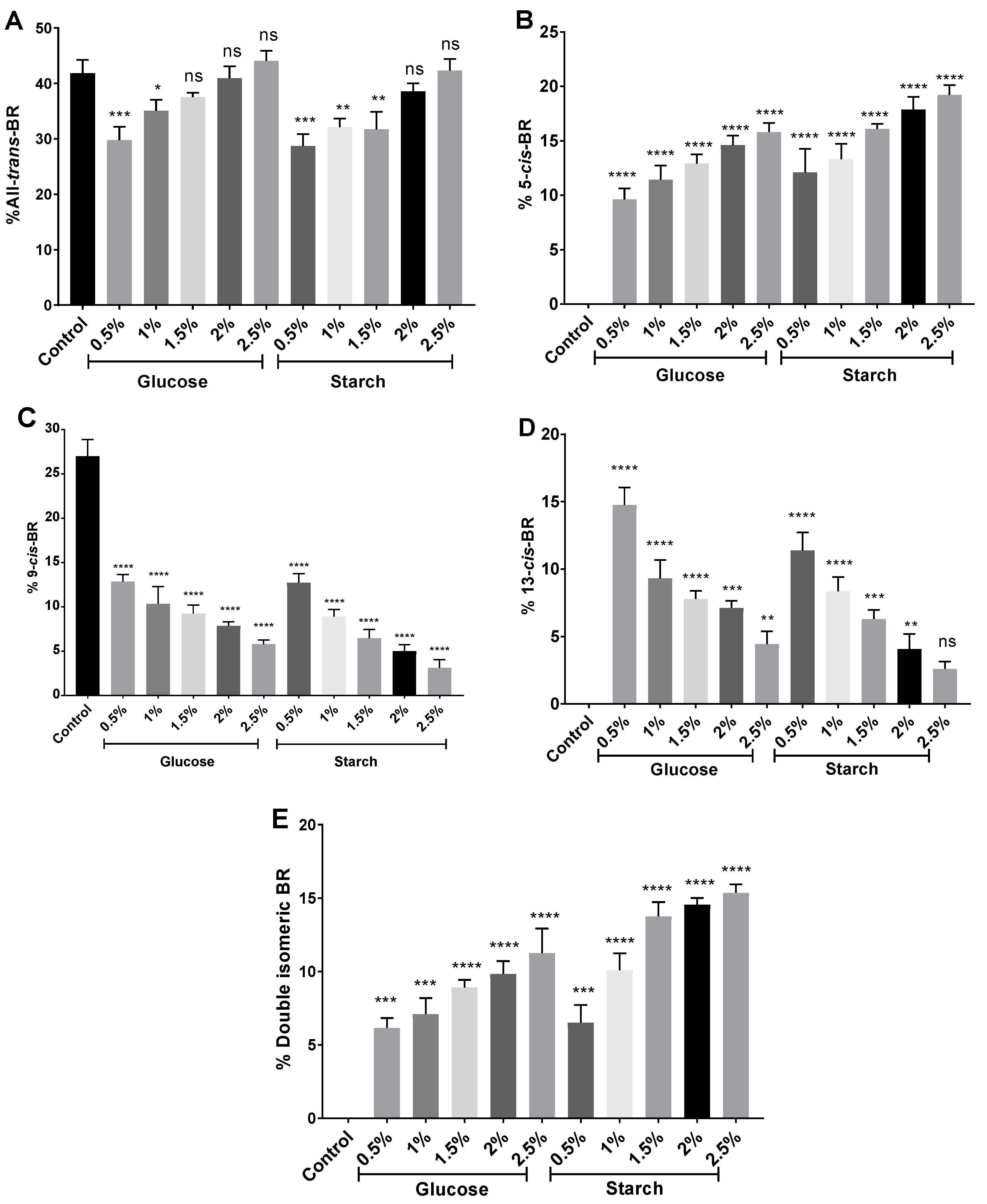
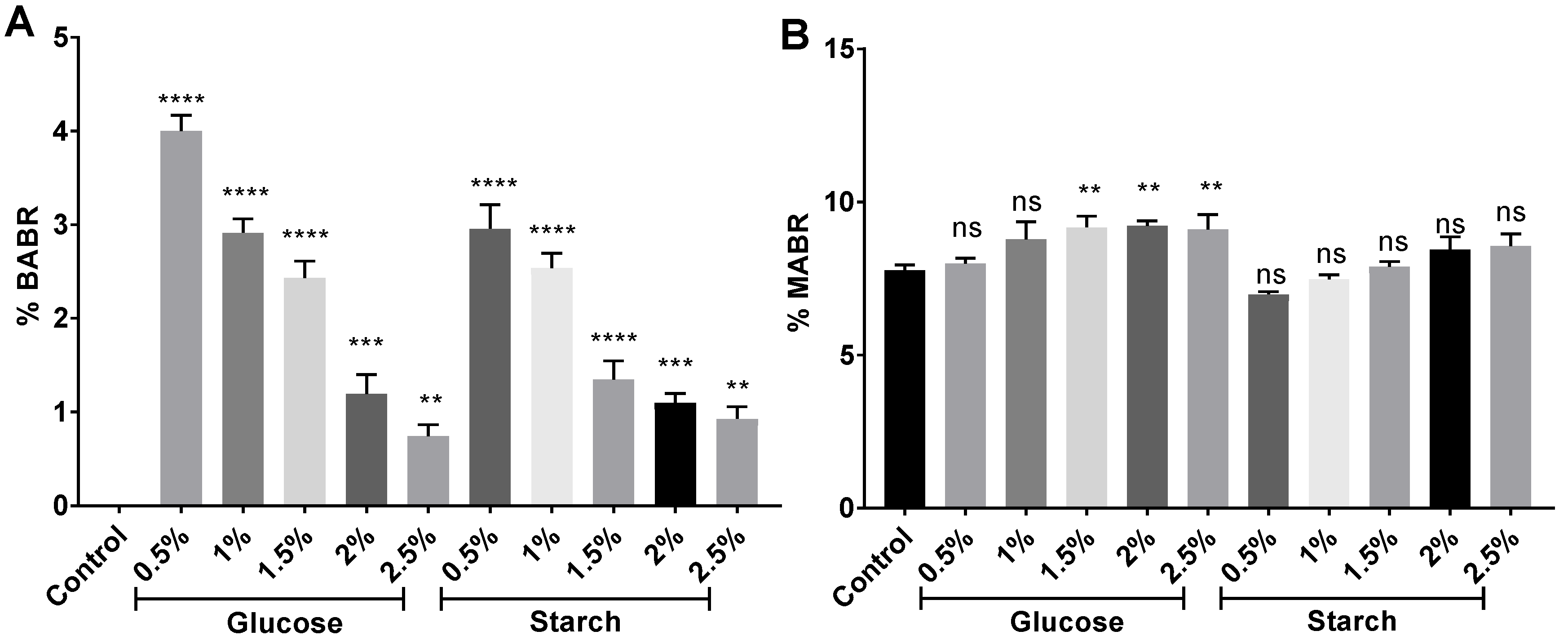
2.5. Carotenoid Composition by HPLC-MS
3. Materials and Methods
3.1. Culture Growth Conditions
3.1.1. Cultivation in the Presence of Different Concentrations of Carbon Sources
3.1.2. Growth Determination
3.2. Carotenoid Extraction and BR Quantification
3.3. In Vitro Antioxidant Activity
3.4. Carbohydrate-Hydrolyzing Enzymes and Lipase Inhibitory Activities
3.5. Determination of the Composition of Carotenoid Extracts by HPLC Analysis
3.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Rao, A.; Rao, L. Carotenoids and Human Health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A Global Perspective on Carotenoids: Metabolism, Biotechnology, and Benefits for Nutrition and Health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, Pharmacology and Treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [Green Version]
- Giani, M.; Montoyo-Pujol, Y.G.; Peiró, G.; Martínez-Espinosa, R.M. Halophilic Carotenoids and Breast Cancer: From Salt Marshes to Biomedicine. Mar. Drugs 2021, 19, 594. [Google Scholar] [CrossRef] [PubMed]
- Yabuzaki, J. Carotenoids Database: Structures, Chemical Fingerprints and Distribution among Organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef] [Green Version]
- Giani, M.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Haloarchaeal Carotenoids: Healthy Novel Compounds from Extreme Environments. Mar. Drugs 2019, 17, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A. The Microbiology of Red Brines. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 113, pp. 57–110. ISBN 978-0-12-820709-3. [Google Scholar]
- Oren, A. Halophilic Microbial Communities and Their Environments. Curr. Opin. Biotechnol. 2015, 33, 119–124. [Google Scholar] [CrossRef]
- Ventosa, A.; Oren, A.; Ma, Y. (Eds.) Halophiles and Hypersaline Environments: Current Research and Future Trends; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 978-3-642-20197-4. [Google Scholar]
- Jones, D.L.; Baxter, B.K. DNA Repair and Photoprotection: Mechanisms of Overcoming Environmental Ultraviolet Radiation Exposure in Halophilic Archaea. Front. Microbiol. 2017, 8, 1882. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.; Jensen, S.L.; Theander, O.; Cyvin, S.J.; Hagen, G. Bacterial Carotenoids. XXVI. C50-Carotenoids. 2. Bacterioruberin. Acta Chem. Scand. 1967, 21, 2578–2580. [Google Scholar] [CrossRef]
- Bidle, K.A.; Hanson, T.E.; Howell, K.; Nannen, J. HMG-CoA Reductase Is Regulated by Salinity at the Level of Transcription in Haloferax volcanii. Extremophiles 2007, 11, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Torregrosa-Crespo, J.; Galiana, C.P.; Martínez-Espinosa, R.M. Biocompounds from Haloarchaea and Their Uses in Biotechnology. In Archaea—New Biocatalysts, Novel Pharmaceuticals and Various Biotechnological Applications; Sghaier, H., Najjari, A., Ghedira, K., Eds.; InTechOpen: Rijeka, Croatia, 2017; ISBN 978-953-51-3569-2. [Google Scholar]
- Saito, T.; Miyabe, Y.; Ide, H.; Yamamoto, O. Hydroxyl Radical Scavenging Ability of Bacterioruberin. Radiat. Phys. Chem. 1997, 50, 267–269. [Google Scholar] [CrossRef]
- Yatsunami, R.; Ando, A.; Yang, Y.; Takaichi, S.; Kohno, M.; Matsumura, Y.; Ikeda, H.; Fukui, T.; Nakasone, K.; Fujita, N.; et al. Identification of Carotenoids from the Extremely Halophilic Archaeon Haloarcula japonica. Front. Microbiol. 2014, 5, 100. [Google Scholar] [CrossRef]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.; Martínez-Espinosa, R. Carotenoids from Haloarchaea and Their Potential in Biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazy, G.E.; Abu-Serie, M.M.; Abo-Elela, G.M.; Ghozlan, H.; Sabry, S.A.; Soliman, N.A.; Abdel-Fattah, Y.R. In Vitro Dual (Anticancer and Antiviral) Activity of the Carotenoids Produced by Haloalkaliphilic Archaeon Natrialba sp. M6. Sci. Rep. 2020, 10, 5986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalazar, L.; Pagola, P.; Miró, M.V.; Churio, M.S.; Cerletti, M.; Martínez, C.; Iniesta-Cuerda, M.; Soler, A.J.; Cesari, A.; de Castro, R. Bacterioruberin Extracts from a Genetically Modified Hyperpigmented Haloferax volcanii Strain: Antioxidant Activity and Bioactive Properties on Sperm Cells. J. Appl. Microbiol. 2019, 126, 796–810. [Google Scholar] [CrossRef]
- Flores, N.; Hoyos, S.; Venegas, M.; Galetović, A.; Zúñiga, L.M.; Fábrega, F.; Paredes, B.; Salazar-Ardiles, C.; Vilo, C.; Ascaso, C.; et al. Haloterrigena Sp. Strain SGH1, a Bacterioruberin-Rich, Perchlorate-Tolerant Halophilic Archaeon Isolated From Halite Microbial Communities, Atacama Desert, Chile. Front. Microbiol. 2020, 11, 324. [Google Scholar] [CrossRef]
- Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Bonete, M.-J.; León, R.; Kharroub, K. Characterization and Biological Activities of Carotenoids Produced by Three Haloarchaeal Strains Isolated from Algerian Salt Lakes. Arch. Microbiol. 2022, 204, 6. [Google Scholar] [CrossRef]
- Henke, N.A.; Frohwitter, J.; Peters-Wendisch, P.; Wendisch, V.F. Carotenoid Production by Recombinant Corynebacterium Glutamicum: Strain Construction, Cultivation, Extraction, and Quantification of Carotenoids and Terpenes. In Microbial Carotenoids; Methods in Molecular Biology; Barreiro, C., Barredo, J.-L., Eds.; Springer: New York, NY, USA, 2018; Volume 1852, pp. 127–141. ISBN 978-1-4939-8741-2. [Google Scholar]
- Mussagy, C.U.; Winterburn, J.; Santos-Ebinuma, V.C.; Pereira, J.F.B. Production and Extraction of Carotenoids Produced by Microorganisms. Appl. Microbiol. Biotechnol. 2019, 103, 1095–1114. [Google Scholar] [CrossRef] [PubMed]
- Mata-Gómez, L.C.; Montañez, J.C.; Méndez-Zavala, A.; Aguilar, C.N. Biotechnological Production of Carotenoids by Yeasts: An Overview. Microb. Cell Fact. 2014, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Montero-Lobato, Z.; Ramos-Merchante, A.; Fuentes, J.; Sayago, A.; Fernández-Recamales, Á.; Martínez-Espinosa, R.; Vega, J.; Vílchez, C.; Garbayo, I. Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology. Mar. Drugs 2018, 16, 372. [Google Scholar] [CrossRef]
- Naziri, D.; Hamidi, M.; Hassanzadeh, S.; Tarhriz, V.; Maleki Zanjani, B.; Nazemyieh, H.; Hejazi, M.A.; Hejazi, M.S. Analysis of Carotenoid Production by Halorubrum sp. TBZ126; an Extremely Halophilic Archeon from Urmia Lake. Adv. Pharm. Bull. 2014, 4, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.-Y.; Duan, K.-J.; Huang, S.-Y.; Chen, C.W. Production of Polyhydroxyalkanoates from Inexpensive Extruded Rice Bran and Starch by Haloferax mediterranei. J. Ind. Microbiol. Biotechnol. 2006, 33, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Quillaguamán, J.; Guzmán, H.; Van-Thuoc, D.; Hatti-Kaul, R. Synthesis and Production of Polyhydroxyalkanoates by Halophiles: Current Potential and Future Prospects. Appl. Microbiol. Biotechnol. 2010, 85, 1687–1696. [Google Scholar] [CrossRef]
- Torregrosa-Crespo, J.; Martínez-Espinosa, R.M.; Esclapez, J.; Bautista, V.; Pire, C.; Camacho, M.; Richardson, D.J.; Bonete, M.J. Anaerobic Metabolism in Haloferax Genus. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 68, pp. 41–85. ISBN 978-0-12-804823-8. [Google Scholar]
- Giani, M.; Montero-Lobato, Z.; Garbayo, I.; Vílchez, C.; Vega, J.M.; Martínez-Espinosa, R.M. Haloferax mediterranei Cells as C50 Carotenoid Factories. Mar. Drugs 2021, 19, 100. [Google Scholar] [CrossRef]
- Koller, M.; Maršálek, L.; de Sousa Dias, M.M.; Braunegg, G. Producing Microbial Polyhydroxyalkanoate (PHA) Biopolyesters in a Sustainable Manner. New Biotechnol. 2017, 37, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, R. Production of Polyhydroxyalkanoates (PHA) by Haloferax mediterranei from Food Waste Derived Nutrients for Biodegradable Plastic Applications. J. Microbiol. Biotechnol. 2021, 31, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Simó-Cabrera, L.; García-Chumillas, S.; Hagagy, N.; Saddiq, A.; Tag, H.; Selim, S.; AbdElgawad, H.; Arribas Agüero, A.; Monzó Sánchez, F.; Cánovas, V.; et al. Haloarchaea as Cell Factories to Produce Bioplastics. Mar. Drugs 2021, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, V.; Garcia-Chumillas, S.; Monzó, F.; Simó-Cabrera, L.; Fernández-Ayuso, C.; Pire, C.; Martínez-Espinosa, R.M. Analysis of Polyhydroxyalkanoates Granules in Haloferax mediterranei by Double-Fluorescence Staining with Nile Red and SYBR Green by Confocal Fluorescence Microscopy. Polymers 2021, 13, 1582. [Google Scholar] [CrossRef]
- Torregrosa-Crespo, J.; Pire, C.; Bergaust, L.; Martínez-Espinosa, R.M. Haloferax mediterranei, an Archaeal Model for Denitrification in Saline Systems, Characterized Through Integrated Physiological and Transcriptional Analyses. Front. Microbiol. 2020, 11, 768. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Cui, H.-L. In Vitro Antioxidant, Antihemolytic, and Anticancer Activity of the Carotenoids from Halophilic Archaea. Curr. Microbiol. 2018, 75, 266–271. [Google Scholar] [CrossRef]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the Extreme Halophilic Archaeon Haloterrigena turkmenica: Identification and Antioxidant Activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.E.; Altekar, W.; D’Souza, S.F. Adaptive Response of Haloferax mediterranei to Low Concentrations of NaCl (<20%) in the Growth Medium. Arch. Microbiol. 1997, 168, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Glovaci, D.; Fan, W.; Wong, N.D. Epidemiology of Diabetes Mellitus and Cardiovascular Disease. Curr. Cardiol. Rep. 2019, 21, 21. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yan, Z.; Chen, Y.; Liu, F. A Social Contagious Model of the Obesity Epidemic. Sci. Rep. 2016, 6, 37961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; McGowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The Obesity Transition: Stages of the Global Epidemic. Lancet Diabetes Endocrinol. 2019, 7, 231–240. [Google Scholar] [CrossRef]
- Ortega, F.B.; Lavie, C.J.; Blair, S.N. Obesity and Cardiovascular Disease. Circ. Res. 2016, 118, 1752–1770. [Google Scholar] [CrossRef] [Green Version]
- Balakumar, P.; Maung-U, K.; Jagadeesh, G. Prevalence and Prevention of Cardiovascular Disease and Diabetes Mellitus. Pharmacol. Res. 2016, 113, 600–609. [Google Scholar] [CrossRef]
- Baron, A.D. Postprandial Hyperglycaemia and α-Glucosidase Inhibitors. Diabetes Res. Clin. Pract. 1998, 40, S51–S55. [Google Scholar] [CrossRef]
- Chiasson, J.-L.; Josse, R.G.; Gomis, R.; Hanefeld, M.; Karasik, A.; Laakso, M. Acarbose for Prevention of Type 2 Diabetes Mellitus: The STOP-NIDDM Randomised Trial. Lancet 2002, 359, 2072–2077. [Google Scholar] [CrossRef]
- Hu, R.; Li, Y.; Lv, Q.; Wu, T.; Tong, N. Acarbose Monotherapy and Type 2 Diabetes Prevention in Eastern and Western Prediabetes: An Ethnicity-Specific Meta-Analysis. Clin. Ther. 2015, 37, 1798–1812. [Google Scholar] [CrossRef]
- Kumar, A.; Chauhan, S. Pancreatic Lipase Inhibitors: The Road Voyaged and Successes. Life Sci. 2021, 271, 119115. [Google Scholar] [CrossRef] [PubMed]
- Bialecka-Florjanczyk, E.; Fabiszewska, A.U.; Krzyczkowska, J.; Kurylowicz, A. Synthetic and Natural Lipase Inhibitors. Mini Rev. Med. Chem. 2018, 18, 672–683. [Google Scholar] [CrossRef]
- Kuprat, T.; Ortjohann, M.; Johnsen, U.; Schönheit, P. Glucose Metabolism and Acetate Switch in Archaea: The Enzymes in Haloferax volcanii. J. Bacteriol. 2021, 203, e00690-20. [Google Scholar] [CrossRef]
- Vázquez-Madrigal, A.S.; Barbachano-Torres, A.; Arellano-Plaza, M.; Kirchmayr, M.R.; Finore, I.; Poli, A.; Nicolaus, B.; de la Torre Zavala, S.; Camacho-Ruiz, R.M. Effect of Carbon Sources in Carotenoid Production from Haloarcula sp. M1, Halolamina sp. M3 and Halorubrum sp. M5, Halophilic Archaea Isolated from Sonora Saltern, Mexico. Microorganisms 2021, 9, 1096. [Google Scholar] [CrossRef]
- Will Chen, C.; Hsu, S.; Lin, M.-T.; Hsu, Y. Mass Production of C50 Carotenoids by Haloferax mediterranei in Using Extruded Rice Bran and Starch under Optimal Conductivity of Brined Medium. Bioprocess Biosyst. Eng. 2015, 38, 2361–2367. [Google Scholar] [CrossRef] [PubMed]
- Gochnauer, M.B.; Kushwaha, S.C.; Kates, M.; Kushner, D.J. Nutritional Control of Pigment and Isoprenoid Compound Formation in Extremely Halophilic Bacteria. Archiv. Mikrobiol. 1972, 84, 339–349. [Google Scholar] [CrossRef]
- Mandelli, F.; Miranda, V.S.; Rodrigues, E.; Mercadante, A.Z. Identification of Carotenoids with High Antioxidant Capacity Produced by Extremophile Microorganisms. World J. Microbiol. Biotechnol. 2012, 28, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Melendez-Martínez, A.; Britton, G.; Vicario, I.; Heredia, F. HPLC Analysis of Geometrical Isomers of Lutein Epoxide Isolated from Dandelion (Taraxacum officinale F. Weber Ex Wiggers). Phytochemistry 2006, 67, 771–777. [Google Scholar] [CrossRef]
- Cerletti, M.; Paggi, R.; Troetschel, C.; Ferrari, M.C.; Guevara, C.R.; Albaum, S.; Poetsch, A.; de Castro, R. LonB Protease Is a Novel Regulator of Carotenogenesis Controlling Degradation of Phytoene Synthase in Haloferax volcanii. J. Proteome Res. 2018, 17, 1158–1171. [Google Scholar] [CrossRef] [Green Version]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological Properties of Carotenoids Extracted from Halobacterium halobium Isolated from a Tunisian Solar Saltern. BMC Complement. Altern. Med. 2013, 13, 255. [Google Scholar] [CrossRef]
- Kumar, P.; Jun, H.-B.; Kim, B.S. Co-Production of Polyhydroxyalkanoates and Carotenoids through Bioconversion of Glycerol by Paracoccus sp. Strain LL1. Int. J. Biol. Macromol. 2018, 107, 2552–2558. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Villegas, P.; Vigara, J.; Vila, M.; Varela, J.; Barreira, L.; Léon, R. Antioxidant, Antimicrobial, and Bioactive Potential of Two New Haloarchaeal Strains Isolated from Odiel Salterns (Southwest Spain). Biology 2020, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Lizama, C.; Romero-Parra, J.; Andrade, D.; Riveros, F.; Bórquez, J.; Ahmed, S.; Venegas-Salas, L.; Cabalín, C.; Simirgiotis, M.J. Analysis of Carotenoids in Haloarchaea Species from Atacama Saline Lakes by High Resolution UHPLC-Q-Orbitrap-Mass Spectrometry: Antioxidant Potential and Biological Effect on Cell Viability. Antioxidants 2021, 10, 1230. [Google Scholar] [CrossRef] [PubMed]
- Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Ghennai, O.; Bonete, M.-J.; León, R.; Kharroub, K. Bioprospecting and Characterization of Pigmented Halophilic Archaeal Strains from Algerian Hypersaline Environments with Analysis of Carotenoids Produced by Halorubrum sp. BS2. J. Basic Microbiol. 2020, 60, 624–638. [Google Scholar] [CrossRef] [PubMed]
- Dose, J.; Matsugo, S.; Yokokawa, H.; Koshida, Y.; Okazaki, S.; Seidel, U.; Eggersdorfer, M.; Rimbach, G.; Esatbeyoglu, T. Free Radical Scavenging and Cellular Antioxidant Properties of Astaxanthin. Int. J. Mol. Sci. 2016, 17, 103. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.-C.; Lin, J.-T.; Lu, F.-J.; Chou, F.-P.; Yang, D.-J. Determination of Carotenoids in Dunaliella salina Cultivated in Taiwan and Antioxidant Capacity of the Algal Carotenoid Extract. Food Chem. 2008, 109, 439–446. [Google Scholar] [CrossRef]
- Singh, P.; Baranwal, M.; Reddy, S.M. Antioxidant and Cytotoxic Activity of Carotenes Produced by Dunaliella salina under Stress. Pharm. Biol. 2016, 54, 2269–2275. [Google Scholar] [CrossRef] [Green Version]
- Bellahcen, T.O.; AAmiri, A.; Touam, I.; Hmimid, F.; Amrani, A.E.; Cherif, A.; Cherki, M. Evaluation of Moroccan Microalgae: Spirulina platensis as a Potential Source of Natural Antioxidants. J. Complement. Integr. Med. 2020, 17, 20190036. [Google Scholar] [CrossRef]
- Sun, T.; Tanumihardjo, S.A. An Integrated Approach to Evaluate Food Antioxidant Capacity. J Food Sci. 2007, 72, R159–R165. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative Stress and Metabolic Disorders: Pathogenesis and Therapeutic Strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Karam, B.S.; Chavez-Moreno, A.; Koh, W.; Akar, J.G.; Akar, F.G. Oxidative Stress and Inflammation as Central Mediators of Atrial Fibrillation in Obesity and Diabetes. Cardiovasc. Diabetol. 2017, 16, 120. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.A.T.; Wang, J.; Tang, J.; Lee, Y.Z.; Ng, K. Evaluation of α-Glucosidase Inhibition Potential of Some Flavonoids from Epimedium Brevicornum. LWT-Food Sci. Technol. 2013, 53, 492–498. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Z.; Yuan, T.; Wang, H.; Xie, X.; Fu, Z. Antioxidants and α-Glucosidase Inhibitors from Ipomoea batatas Leaves Identified by Bioassay-Guided Approach and Structure-Activity Relationships. Food Chem. 2016, 208, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Guo, M. Rapid Screening for α-Glucosidase Inhibitors from Gymnema sylvestre by Affinity Ultrafiltration–HPLC-MS. Front. Pharmacol. 2017, 8, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, H.-C.; Li, S.-L.; Zhang, X.-Q.; Ye, W.-C.; Zhang, Q.-W. Flavonoids with α-Glucosidase Inhibitory Activities and Their Contents in the Leaves of Morus atropurpurea. Chin. Med. 2013, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Li, Y.; Zhang, Q.; Wei, X.; Peng, A.; Chen, L.; Wei, Y. Triterpene Acids Isolated from Lagerstroemia speciosa Leaves as α-Glucosidase Inhibitors: Triterpene acids as alpha-glucosidase inhibitors. Phytother. Res. 2009, 23, 614–618. [Google Scholar] [CrossRef]
- Abbas, G.; Al Harrasi, A.; Hussain, H.; Hamaed, A.; Supuran, C.T. The Management of Diabetes Mellitus-Imperative Role of Natural Products against Dipeptidyl Peptidase-4, α-Glucosidase and Sodium-Dependent Glucose Co-Transporter 2 (SGLT2). Bioorg. Chem. 2019, 86, 305–315. [Google Scholar] [CrossRef]
- Ramírez, G.; Zavala, M.; Pérez, J.; Zamilpa, A. In Vitro Screening of Medicinal Plants Used in Mexico as Antidiabetics with Glucosidase and Lipase Inhibitory Activities. Evid.-Based Complement. Altern. Med. 2012, 2012, 701261. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chi, G.; Wang, L.; Wang, F.; He, S. Isolation, Identification, and Inhibitory Enzyme Activity of Phenolic Substances Present in Spirulina. J. Food Biochem. 2020, 44, e13356. [Google Scholar] [CrossRef]
- Prabakaran, G.; Sampathkumar, P.; Kavisri, M.; Moovendhan, M. Extraction and Characterization of Phycocyanin from Spirulina platensis and Evaluation of Its Anticancer, Antidiabetic and Antiinflammatory Effect. Int. J. Biol. Macromol. 2020, 153, 256–263. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Hung, Y.-L.; Tsai, Y.-K.; Chien, S.-Y.; Kong, Z.-L. The Brown Seaweed Sargassum hemiphyllum Exhibits α-Amylase and α-Glucosidase Inhibitory Activity and Enhances Insulin Release in Vitro. Cytotechnology 2015, 67, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Unnikrishnan, P.; Suthindhiran, K.; Jayasri, M. Alpha-Amylase Inhibition and Antioxidant Activity of Marine Green Algae and Its Possible Role in Diabetes Management. Pharmacogn. Mag. 2015, 11, 511. [Google Scholar] [CrossRef] [Green Version]
- Lowe, M.E. Structure and Function of Pancreatic Lipase and Colipase. Annu. Rev. Nutr. 1997, 17, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xing, D.; Wang, C. Pancreatic Triglyceride Lipase Inhibitors Derived from Natural Products: How to Dig into the Truth. J. Agric. Food Chem. 2021, 69, 6097–6099. [Google Scholar] [CrossRef] [PubMed]
- Samuel Wu, Y.-H.; Chiu, C.-H.; Yang, D.-J.; Lin, Y.-L.; Tseng, J.-K.; Chen, Y.-C. Inhibitory Effects of Litchi (Litchi chinensis Sonn.) Flower-Water Extracts on Lipase Activity and Diet-Induced Obesity. J. Funct. Foods 2013, 5, 923–929. [Google Scholar] [CrossRef]
- Matsuo, Y.; Matsumoto, K.; Inaba, N.; Mimaki, Y. Daisaikoto Inhibits Pancreatic Lipase Activity and Decreases Serum Triglyceride Levels in Mice. Biol. Pharm. Bull. 2018, 41, 1485–1488. [Google Scholar] [CrossRef] [Green Version]
- Loizzo, M.R.; Marrelli, M.; Pugliese, A.; Conforti, F.; Nadjafi, F.; Menichini, F.; Tundis, R. Crocus cancellatus subsp. Damascenus Stigmas: Chemical Profile, and Inhibition of α -Amylase, α-Glucosidase and Lipase, Key Enzymes Related to Type 2 Diabetes and Obesity. J. Enzym. Inhib. Med. Chem. 2016, 31, 212–218. [Google Scholar] [CrossRef]
- Ronnekleiv, M. Bacterial Carotenoids 53∗ C50-Carotenoids 23; Carotenoids of Haloferax volcanii versus Other Halophilic Bacteria. Biochem. Syst. Ecol. 1995, 23, 627–634. [Google Scholar] [CrossRef]
- Fang, C.-J.; Ku, K.-L.; Lee, M.-H.; Su, N.-W. Influence of Nutritive Factors on C50 Carotenoids Production by Haloferax mediterranei ATCC 33500 with Two-Stage Cultivation. Bioresour. Technol. 2010, 101, 6487–6493. [Google Scholar] [CrossRef]
- Asker, D.; Awad, T.; Ohta, Y. Lipids of Haloferax alexandrinus strain TMT: An Extremely Halophilic Canthaxanthin-Producing Archaeon. J. Biosci. Bioeng. 2002, 93, 37–43. [Google Scholar] [CrossRef]
- De la Vega, M.; Sayago, A.; Ariza, J.; Barneto, A.G.; León, R. Characterization of a Bacterioruberin-Producing Haloarchaea Isolated from the Marshlands of the Odiel River in the Southwest of Spain. Biotechnol. Prog. 2016, 32, 592–600. [Google Scholar] [CrossRef]
- Asker, D.; Ohta, Y. Production of Canthaxanthin by Extremely Halophilic Bacteria. J. Biosci. Bioeng. 1999, 88, 617–621. [Google Scholar] [CrossRef]
- Rodriguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Behaviour of Mixed Populations of Halophilic Bacteria in Continuous Cultures. Can. J. Microbiol. 1980, 26, 1259–1263. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Leporini, M.; Sicari, V.; Falco, T.; Pellicanò, T.M.; Tundis, R. Investigating the in Vitro Hypoglycaemic and Antioxidant Properties of Citrus × Clementina Hort. Juice. Eur. Food Res. Technol. 2018, 244, 523–534. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; de Luca, D.; O’Brien, N.; Menichini, F.; Tundis, R. Influence of Drying and Cooking Process on the Phytochemical Content, Antioxidant and Hypoglycaemic Properties of Two Bell Capsicum annum L. Cultivars. Food Chem. Toxicol. 2013, 53, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.; Lucci, P.; Núñez, O.; Tundis, R.; Balzano, M.; Frega, N.; Conte, L.; Moret, S.; Filatova, D.; Moyano, E.; et al. Native Colombian Fruits and Their By-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential. Foods 2019, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Gorjanović, S.Ž.; Alvarez-Suarez, J.M.; Novaković, M.M.; Pastor, F.T.; Pezo, L.; Battino, M.; Sužnjević, D.Ž. Comparative Analysis of Antioxidant Activity of Honey of Different Floral Sources Using Recently Developed Polarographic and Various Spectrophotometric Assays. J. Food Compos. Anal. 2013, 30, 13–18. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | DPPH Test IC50 (µg/mL) | ABTS Test IC50 (µg/mL) | FRAP Test µM Fe (II)/g | β-Carotene Bleaching Test IC50 (µg/mL) | |
|---|---|---|---|---|---|
| 30 min | 60 min | ||||
| Control | 120.0 ± 4.8 | 2.9 ± 0.1 | 18.1 ± 1.4 | 12.1 ± 1.4 | 28.1 ± 2.2 |
| 0.5% Glucose | 73.8 ± 8.2 **** | 0.23 ± 0.058 **** | 20.0 ± 1.6 ns | 14.1 ± 0.7 ** | 21.0 ± 2.1 *** |
| 1% Glucose | 40.3 ± 4.1 **** | 0.12 ± 0.029 **** | 25.3 ± 1.3 **** | 1.5 ± 0.2 **** | 11.9 ± 3.9 **** |
| 1.5% Glucose | 39.4 ± 4.3 **** | 0.05 ± 0.004 **** | 30.7 ± 1.1 **** | 0.51 ± 0.07 **** | 0.75 ± 0.09 **** |
| 2% Glucose | 35.2 ± 2.9 **** | 0.04 ± 0.005 **** | 38.7 ± 1.1 **** | 0.49 ± 0.08 **** | 0.65 ± 0.01 **** |
| 2.5% Glucose | 32.4 ± 3.6 **** | 0.03 ± 0.003 **** | 39.5 ± 0.7 **** | 0.15 ± 0.01 **** | 0.47 ± 0.04 **** |
| 0.5% Starch | 54.4 ± 5.1 **** | 2.0 ± 0.5 **** | 19.6 ± 1.4 ns | 4.1 ± 0.8 **** | 10.2 ± 0.1 **** |
| 1% Starch | 53.0 ± 5.3 **** | 0.10 ± 0.012 **** | 21.0 ± 1.1 * | 2.0 ± 0.3 **** | 4.3 ± 0.4 **** |
| 1.5% Starch | 51.4 ± 6.9 **** | 0.10 ± 0.004 **** | 22.3 ±0.8 *** | 1.4 ± 0.2 **** | 2.0 ± 0.2 **** |
| 2% Starch | 47.6 ± 5.6 **** | 0.05 ± 0.003 **** | 24.2 ± 0.9 **** | 0.28 ± 0.06 **** | 1.6 ± 0.08 **** |
| 2.5% Starch | 40.6 ± 5.0 **** | 0.04 ± 0.006 **** | 30.5 ± 1.4 **** | 0.15 ± 0.008 **** | 1.1 ± 0.11 **** |
| Sample | α-Glucosidase Test IC50 (µg/mL) | α-Amylase Test IC50 (µg/mL) | Lipase Test IC50 (µg/mL) |
|---|---|---|---|
| Control | 117.2 ± 3.2 | 86.9 ± 8.3 | 78.4 ± 5.6 |
| 0.5% Glucose | 73.1 ± 9.7 **** | 23.7 ± 3.5 **** | 80.5 ± 3.5 ns |
| 1% Glucose | 25.4 ± 4.6 **** | 11.5 ± 1.3 **** | 75.2 ± 5.9 ns |
| 1.5% Glucose | 11.9 ± 2.9 **** | 5.1 ± 0.6 **** | 68.3 ± 3.2 ns |
| 2% Glucose | 6.0 ± 1.3 **** | 1.9 ± 0.07 **** | 24.7 ± 0.8 **** |
| 2.5% Glucose | 3.2 ± 0.6 **** | 1.3 ± 0.1 **** | 5.3 ± 0.4 **** |
| 0.5% Starch | 79.8 ± 8.8 **** | 18.8 ± 3.1 **** | 86.0 ± 4.2 ns |
| 1% Starch | 21.0 ± 2.8 **** | 12.7 ± 0.9 **** | 77.8 ± 3.9 ns |
| 1.5% Starch | 13.8 ± 3.3 **** | 3.4 ± 0.4 **** | 70.5 ± 2.6 ns |
| 2% Starch | 7.3 ± 1.3 **** | 1.8 ± 0.2 **** | 46.5 ± 9.7 **** |
| 2.5% Starch | 3.6 ± 0.8 **** | 1.4 ± 0.1 **** | 5.3 ± 0.6 **** |
| BR Isomer | Retention Times (min) | ||
|---|---|---|---|
| Flores et al., 2020 | Lizama et al., 2021 | Hfx. mediterranei (Present Results) | |
| All trans-BR | 10.4 | 4.45 | 3–4 |
| 5-cis BR | 11.2 | 4.92 | 4.2–4.6 |
| 9-cis BR | 11.7 | 5.91 | 4.8–5.2 |
| 13-cis BR | 12.1 | 6.31 | 5.4–5.7 |
| 5-cis-26-cis-BR | - | 7.02 | 7–7.9 |
| 9-cis-26-cis-BR | - | 8.04 | 7–7.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giani, M.; Gervasi, L.; Loizzo, M.R.; Martínez-Espinosa, R.M. Carbon Source Influences Antioxidant, Antiglycemic, and Antilipidemic Activities of Haloferax mediterranei Carotenoid Extracts. Mar. Drugs 2022, 20, 659. https://doi.org/10.3390/md20110659
Giani M, Gervasi L, Loizzo MR, Martínez-Espinosa RM. Carbon Source Influences Antioxidant, Antiglycemic, and Antilipidemic Activities of Haloferax mediterranei Carotenoid Extracts. Marine Drugs. 2022; 20(11):659. https://doi.org/10.3390/md20110659
Chicago/Turabian StyleGiani, Micaela, Luigia Gervasi, Monica Rosa Loizzo, and Rosa María Martínez-Espinosa. 2022. "Carbon Source Influences Antioxidant, Antiglycemic, and Antilipidemic Activities of Haloferax mediterranei Carotenoid Extracts" Marine Drugs 20, no. 11: 659. https://doi.org/10.3390/md20110659