
Inhibition of A549 Lung Cancer Cell Migration and Invasion by Ent-Caprolactin C via the Suppression of Transforming Growth Factor-β-Induced Epithelial—Mesenchymal Transition

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Results
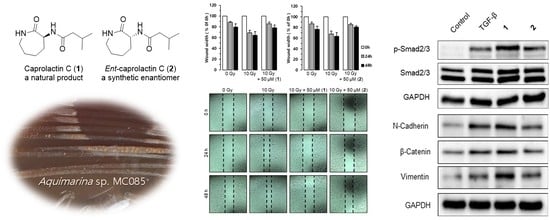
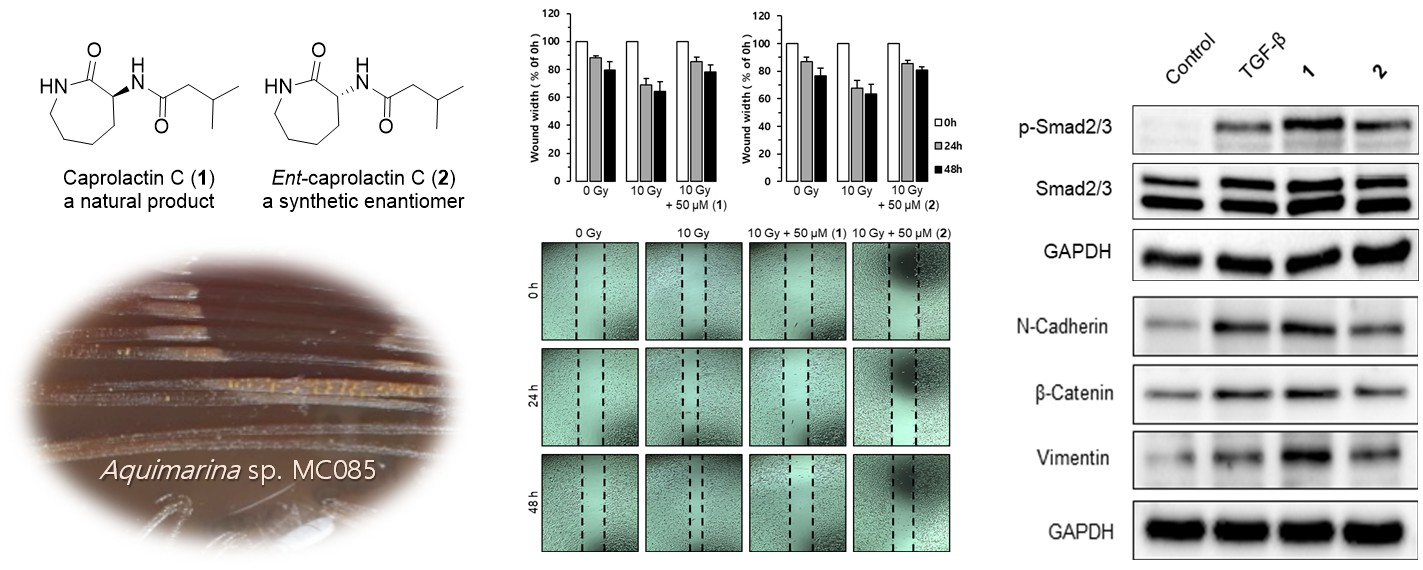
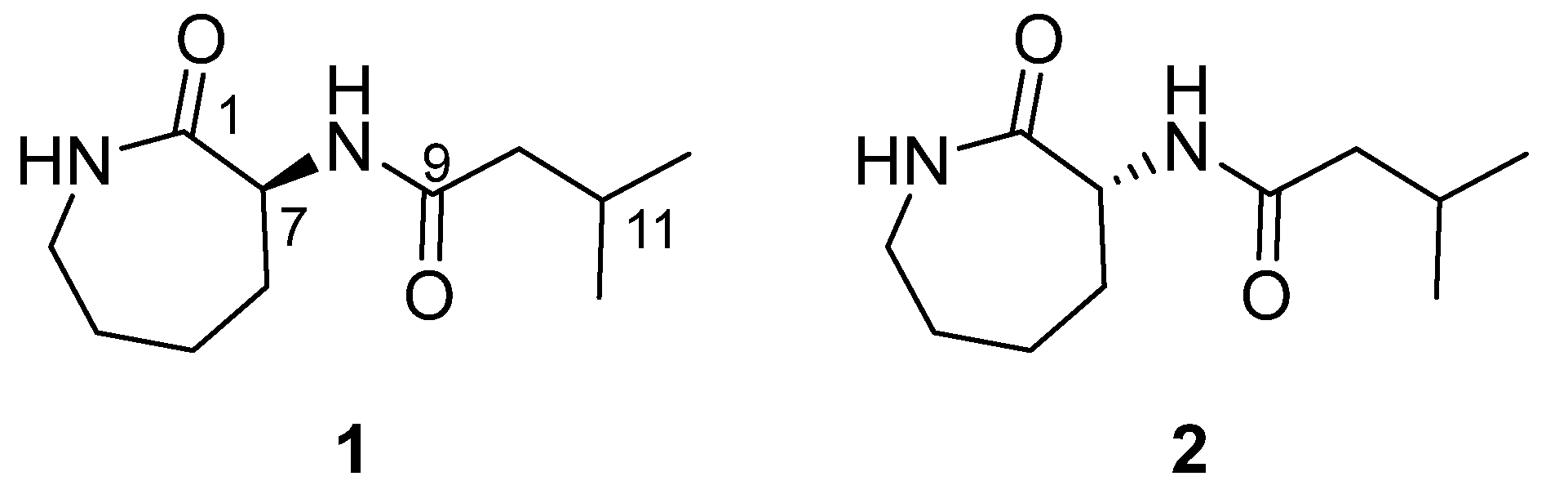
2.1. Structure Elucidation of Compound 1
2.2. Synthesis of Caprolactin C (1) and Ent-Caprolactin C (2)
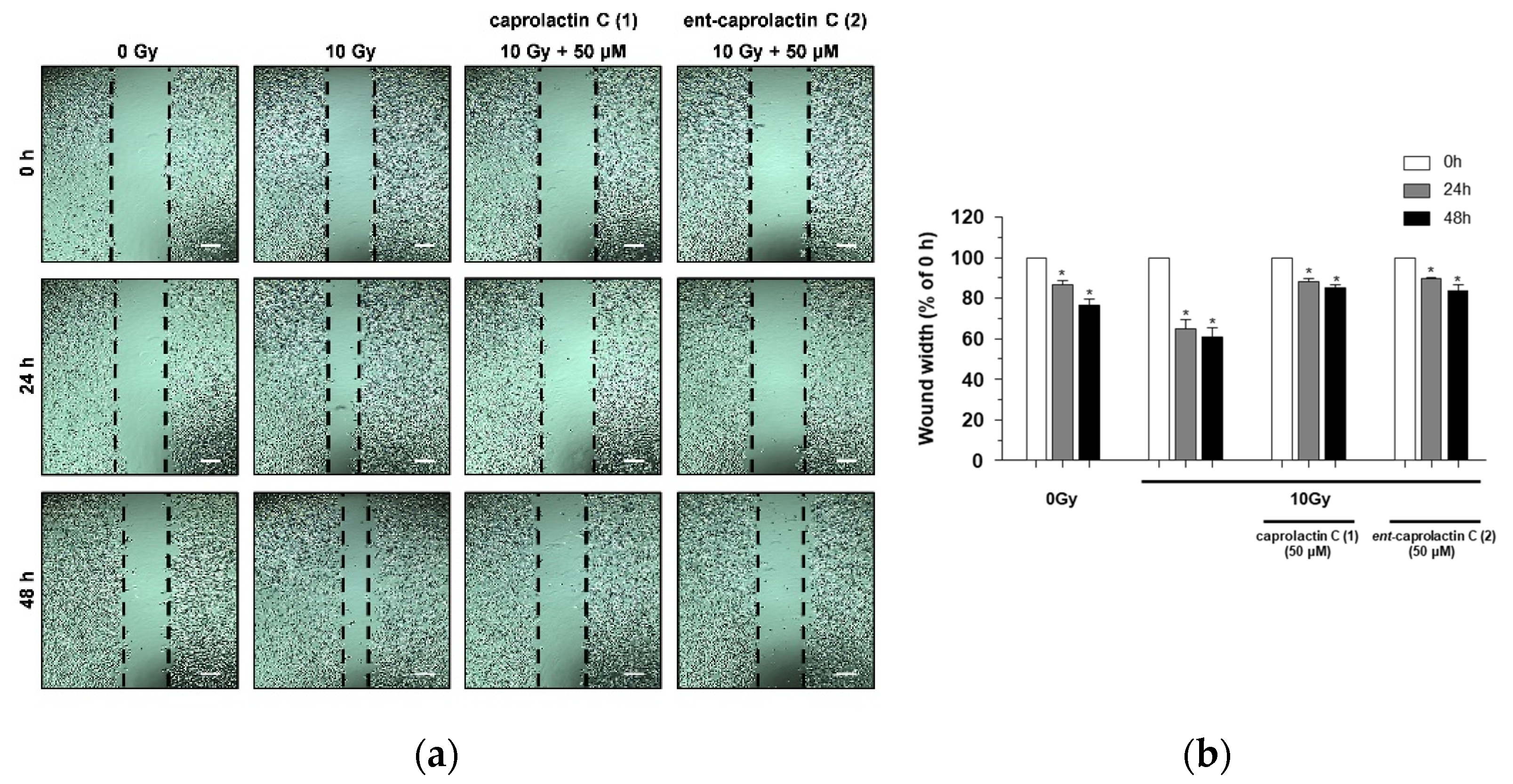
2.3. Effects of Caprolactin C (1) and Ent-Caprolactin C (2) on the Invasion and γ-IR-Induced Migration of A549 Cells
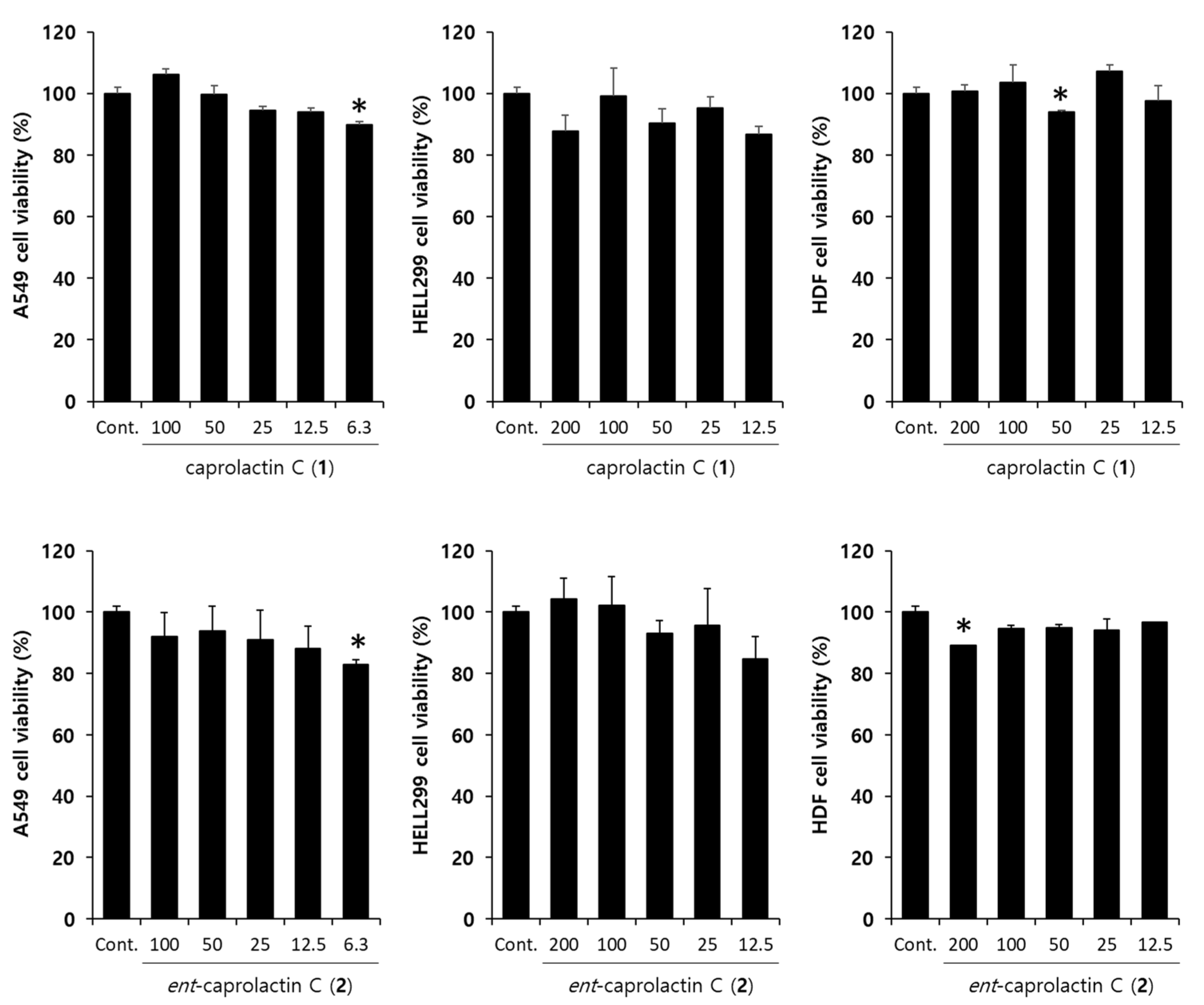
2.4. Effects of Caprolactin C (1) and Ent-Caprolactin C (2) on the Viability of the A549 Cells
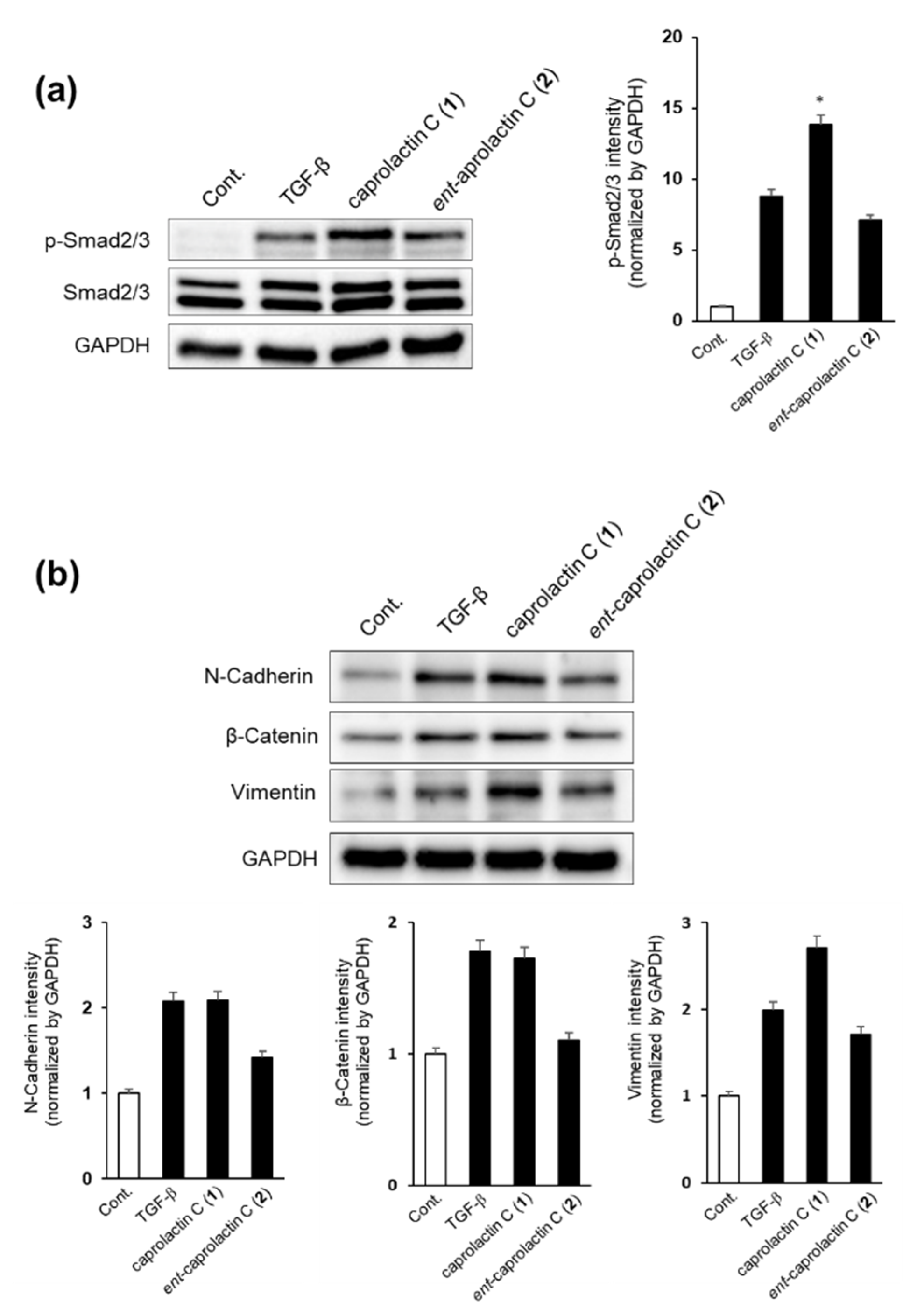
2.5. Effects of Caprolactin C (1) and Ent-Caprolactin C (2) on TGF-β-Induced EMT of A549 Cells
2.6. Effects of Caprolactin C (1) and Ent-Caprolactin C (2) on TGF-β-Induced mRNA Expression Levels of N-Cadherin, MMP-9, Snail, and Vimentin in A549 Cells
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Isolation and Identification of Bacterial Strain
4.3. Extraction and Isolation of Compound 1
Caprolactin C (1)
4.4. Synthesis of Caprolactin C (1) and Ent-Caprolactin C (2)
4.4.1. Synthetic Caprolactin C (1)
4.4.2. Synthetic Ent-Caprolactin C (2)
4.5. Biological Assays
4.5.1. Invasion Assay
4.5.2. γ-IR-Induced Migration Assay (Wound-Healing Assay)
4.5.3. Antibodies and Reagents
4.5.4. Cell Culture
4.5.5. Cell Viability Measurement
4.5.6. Preparation of Cell Lysates for Immunoblotting
4.5.7. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
4.5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Fidler, I.J. The pathogenesis of cancer metastasis: The ’seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.Q.; Walters, E.H.; Shukla, S.D.; Weston, S.; Muller, H.K.; Ward, C.; Sohal, S.S. β-catenin, twist and snail: Transcriptional regulation of EMT in smokers and COPD, and relation to airflow obstruction. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The role of snail in EMT and tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.A. Epithelial–mesenchymal transitions in development and disease: Old views and new perspectives. Int. J. Dev. Biol. 2009, 53, 1541–1547. [Google Scholar] [CrossRef] [Green Version]
- Stone, R.C.; Pastar, I.; Ojeh, N.; Chen, V.; Liu, S.; Garzon, K.I.; Tomic-Canic, M. Epithelial-mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res. 2016, 365, 495–506. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Princep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Nigam, M.; Suleria, H.A.R.; Farzaei, M.H.; Mishra, A.P. Marine anticancer drugs and their relevant targets: A treasure from the ocean. DARU J. Pharm. Sci. 2019, 27, 491–515. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Dybdal-Hargreaves, N.F.; Risinger, A.L.; Mooberry, S.L. Eribulin mesylate: Mechanism of action of a unique microtubule targeting agent. Clin. Cancer Res. 2015, 21, 2445–2452. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.; Kim, G.D.; Jeon, J.-E.; Shin, J.; Lee, S.K. Antimetastatic effect of halichondramide, a trisoxazole macrolide from the marine sponge Chondrosia corticata, on human prostate cancer cells via modulation of epithelial-to-mesenchymal transition. Mar. Drugs 2013, 11, 2472–2485. [Google Scholar] [CrossRef]
- Lin, L.-C.; Kuo, T.-T.; Chang, H.-Y.; Liu, W.-S.; Hsia, S.-M.; Huang, T.-C. Manzamine A exerts anticancer activity against human colorectal cancer cells. Mar. Drugs 2018, 16, 252. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.L.; Kao, Y.C.; Yang, P.H.; Sung, P.J.; Wen, Z.H.; Chen, J.J.; Huang, Y.B.; Chen, P.Y. A small dibromotyrosine derivative purified from Pseudoceratina sp. suppresses TGF-β responsiveness by inhibiting TGF-β type I receptor serine/threonine kinase activity. J. Cell Biochem. 2016, 117, 2800–2814. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-J.; Wei, W.-C.; Dai, G.-F.; Su, J.-H.; Tseng, Y.-H.; Tsai, T.-C. Exploring the mechanism of flaccidoxide-13-acetate in suppressing cell metastasis of hepatocellular carcinoma. Mar. Drugs 2020, 18, 314. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Jeon, Y.J.; Park, S.J. Inhibitory effects of dieckol on hypoxia-induced epithelial-mesenchymal transition of HT29 human colorectal cancer cells. Mol. Med. Rep. 2016, 14, 5148–5154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Braun, D.R.; Ananiev, G.E.; Hoffmann, F.M.; Tsai, I.W.; Rajski, S.R.; Bugni, T.S. Biemamides A-E, inhibitors of the TGF-β pathway that block the epithelial to mesenchymal transition. Org. Lett. 2018, 20, 5529–5532. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V inhibits migration and invasion via suppressing snail/slug-mediated epithelial-mesenchymal transition progression in human breast cancer MDA-MB-231 cells in vitro. Mar. Drugs 2019, 17, 305. [Google Scholar] [CrossRef] [Green Version]
- Shih-Wei, W.; Chih-Ling, C.; Kao, Y.C.; Martin, R.; Knölker, H.J.; Shiao, M.S.; Chen, C.L. Pentabromopseudilin: A myosin V inhibitor suppresses TGF-β activity by recruiting the type II TGF-β receptor to lysosomal degradation. J. Enzyme Inhib. Med. Chem. 2018, 33, 920–935. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Gamage, C.D.B.; Kim, G.J.; Hillman, P.F.; Lee, C.; Lee, E.Y.; Choi, H.; Kim, H.; Nam, S.J.; Fenical, W. Androsamide, a cyclic tetrapeptide from a marine Nocardiopsis sp., suppresses motility of colorectal cancer cells. J. Nat. Prod. 2020, 83, 3166–3172. [Google Scholar] [CrossRef]
- Davidson, B.S.; Schumacher, R.W. Isolation and synthesis of caprolactins A and B, new caprolactams from a marine bacterium. Tetrahedron 1993, 49, 6569–6574. [Google Scholar] [CrossRef]
- Han, A.R.; Lee, S.; Han, S.; Lee, Y.J.; Kim, J.B.; Seo, E.K.; Jung, C.H. Triterpenoids from the leaves of Centella asiatica inhibit ionizing radiation-induced migration and invasion of human lung cancer cells. Evid. Based Complementary Altern. Med. 2020, 2020, 3683460. [Google Scholar] [CrossRef]
- Jung, C.H.; Kim, E.M.; Song, J.Y.; Park, J.K.; Um, H.D. Mitochondrial superoxide dismutase 2 mediates γ-irradiation-induced cancer cell invasion. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derynck, R.; Akhurst, R.J.; Balmain, A. TGF-β signaling in tumor suppression and cancer progression. Nat. Genet. 2001, 29, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Ikushima, H.; Miyazono, K. TGFβ signalling: A complex web in cancer progression. Nat. Rev. Cancer 2020, 10, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, S.; Zeng, J. TGF-β signaling: A complex role in tumorigenesis. Mol. Med. Rep. 2018, 17, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.P.; Rees, S.; Kalindjian, S.B.; Philpott, K.L. Principles of early drug discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.W.; Shin, M.S.; Lee, S.J.; Park, H.R.; Jee, H.S.; Yoon, T.J.; Shin, K.S. Signaling pathways associated with macrophage-activating polysaccharides purified from fermented barley. Int. J. Biol. Macromol. 2019, 131, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Shin, M.S. Inhibitory effects of pectic polysaccharide isolated from Diospyros kaki leaves on tumor cell angiogenesis via VEGF and MMP-9 regulation. Polymers 2021, 13, 64. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Caprolacin C (1) | ||
|---|---|---|---|
| δC1, Mult. | δH2, Mult. (J in Hz) | HMBC (Carbon No.) | |
| 1 | 175.75, C | ||
| 2 | 6.00, br s | ||
| 3a | 42.36, CH2 | 3.30, m | 1, 4, 5 |
| 3b | 3.24, m | 1, 4, 5 | |
| 4a | 29.09, CH2 | 1.84 3, m | 3, 5, 6 |
| 4b | 1.39, m | 3, 6 | |
| 5a | 28.08, CH2 | 1.99, m | 3, 4, 7, 6 |
| 5b | 1.84 3, m | 3, 4, 6, 7 | |
| 6a | 31.90, CH2 | 2.09 3, m | 4, 5 |
| 6b | 1.47, m | 4, 5, 7 | |
| 7 | 52.20, CH | 4.54, ddd (11.34, 5.94, 1.98) | 1, 5, 6, 9 |
| 8 | 6.84, br s | ||
| 9 | 171.80, C | ||
| 10 | 46.09, CH2 | 2.09 3, m | 9, 11, 12, 13 |
| 11 | 26.33, CH | 2.11, m | 9, 10, 12, 13 |
| 12 | 22.67, CH3 | 0.96, d (6.01) | 10, 11, 13 |
| 13 | 22.51, CH3 | 0.95, d (6.20) | 10, 11, 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.Y.; Shin, M.-S.; Kim, G.J.; Kwon, H.; Lee, M.J.; Han, A.-R.; Nam, J.-W.; Jung, C.-H.; Kang, K.S.; Choi, H. Inhibition of A549 Lung Cancer Cell Migration and Invasion by Ent-Caprolactin C via the Suppression of Transforming Growth Factor-β-Induced Epithelial—Mesenchymal Transition. Mar. Drugs 2021, 19, 465. https://doi.org/10.3390/md19080465
Kim SY, Shin M-S, Kim GJ, Kwon H, Lee MJ, Han A-R, Nam J-W, Jung C-H, Kang KS, Choi H. Inhibition of A549 Lung Cancer Cell Migration and Invasion by Ent-Caprolactin C via the Suppression of Transforming Growth Factor-β-Induced Epithelial—Mesenchymal Transition. Marine Drugs. 2021; 19(8):465. https://doi.org/10.3390/md19080465
Chicago/Turabian StyleKim, So Young, Myoung-Sook Shin, Geum Jin Kim, Hyukbean Kwon, Myong Jin Lee, Ah-Reum Han, Joo-Won Nam, Chan-Hun Jung, Ki Sung Kang, and Hyukjae Choi. 2021. "Inhibition of A549 Lung Cancer Cell Migration and Invasion by Ent-Caprolactin C via the Suppression of Transforming Growth Factor-β-Induced Epithelial—Mesenchymal Transition" Marine Drugs 19, no. 8: 465. https://doi.org/10.3390/md19080465