
Structural Characterization and Heparanase Inhibitory Activity of Fucosylated Glycosaminoglycan from Holothuria floridana

Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction, Isolation and Purification of HfFG
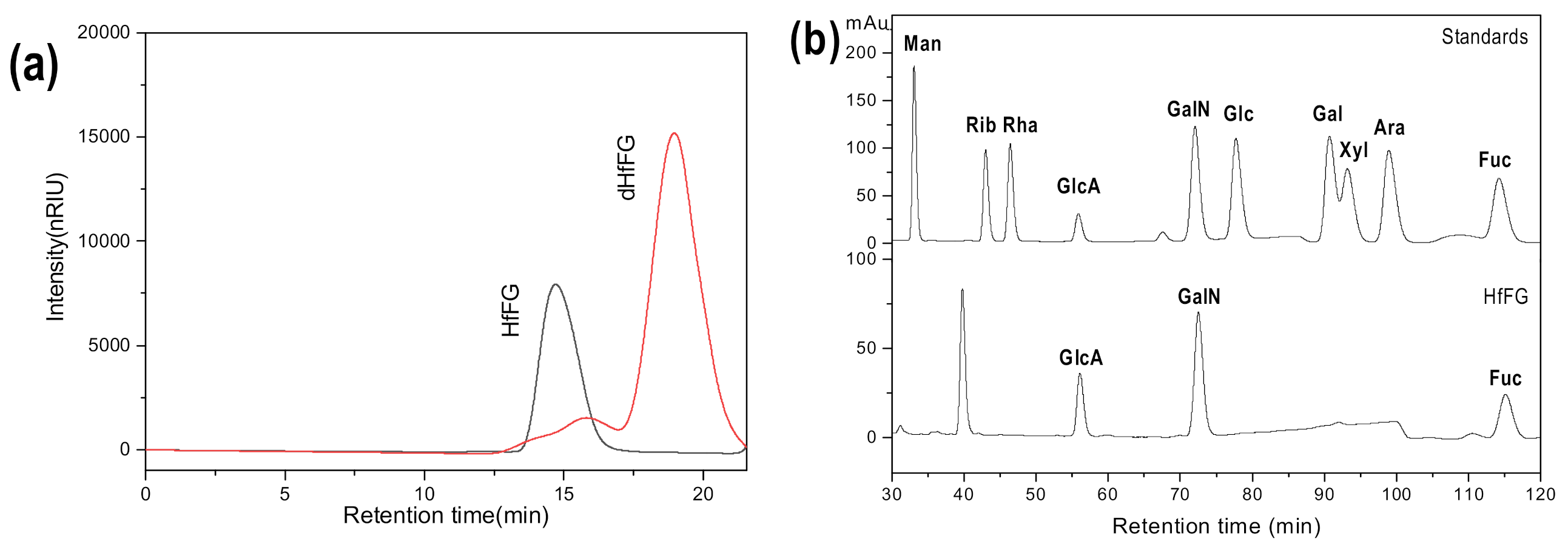
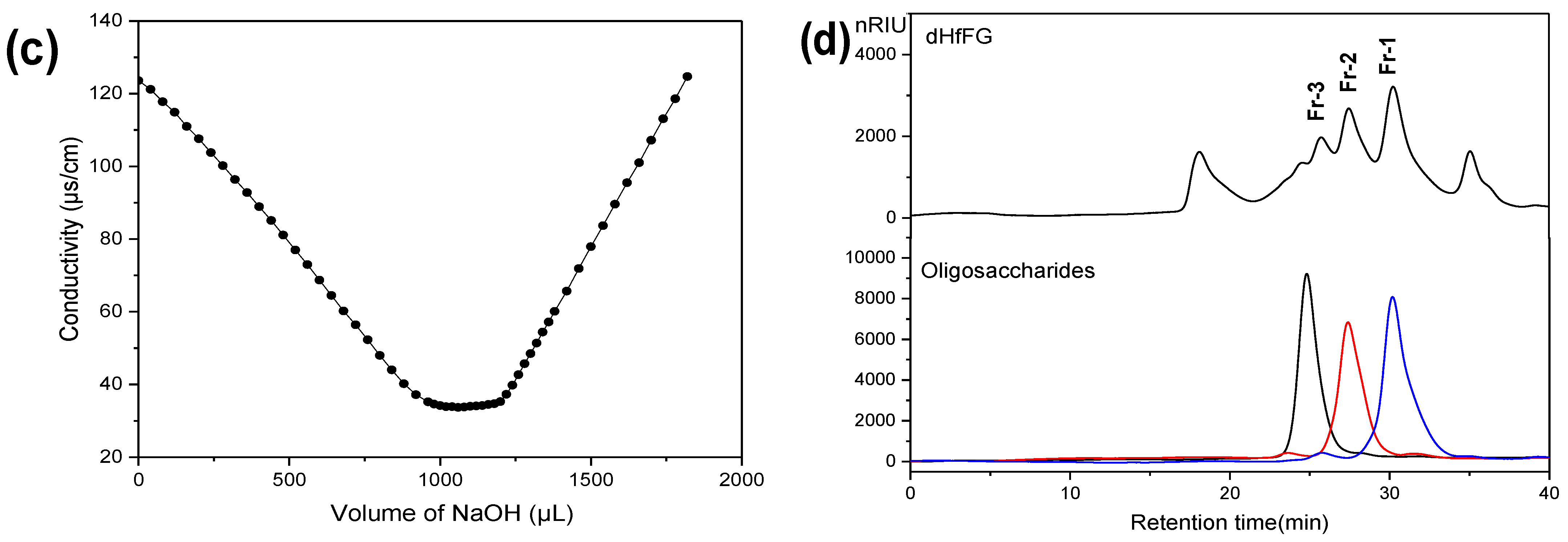
2.2. Chemical Compositions and Physicochemical Properties of HfFG
2.3. Preparation and Structure Characteristics of the Low-Molecular-Weight HfFG (dHfFG)
2.4. Purification of Homogeneous Oligosaccharides Fragments
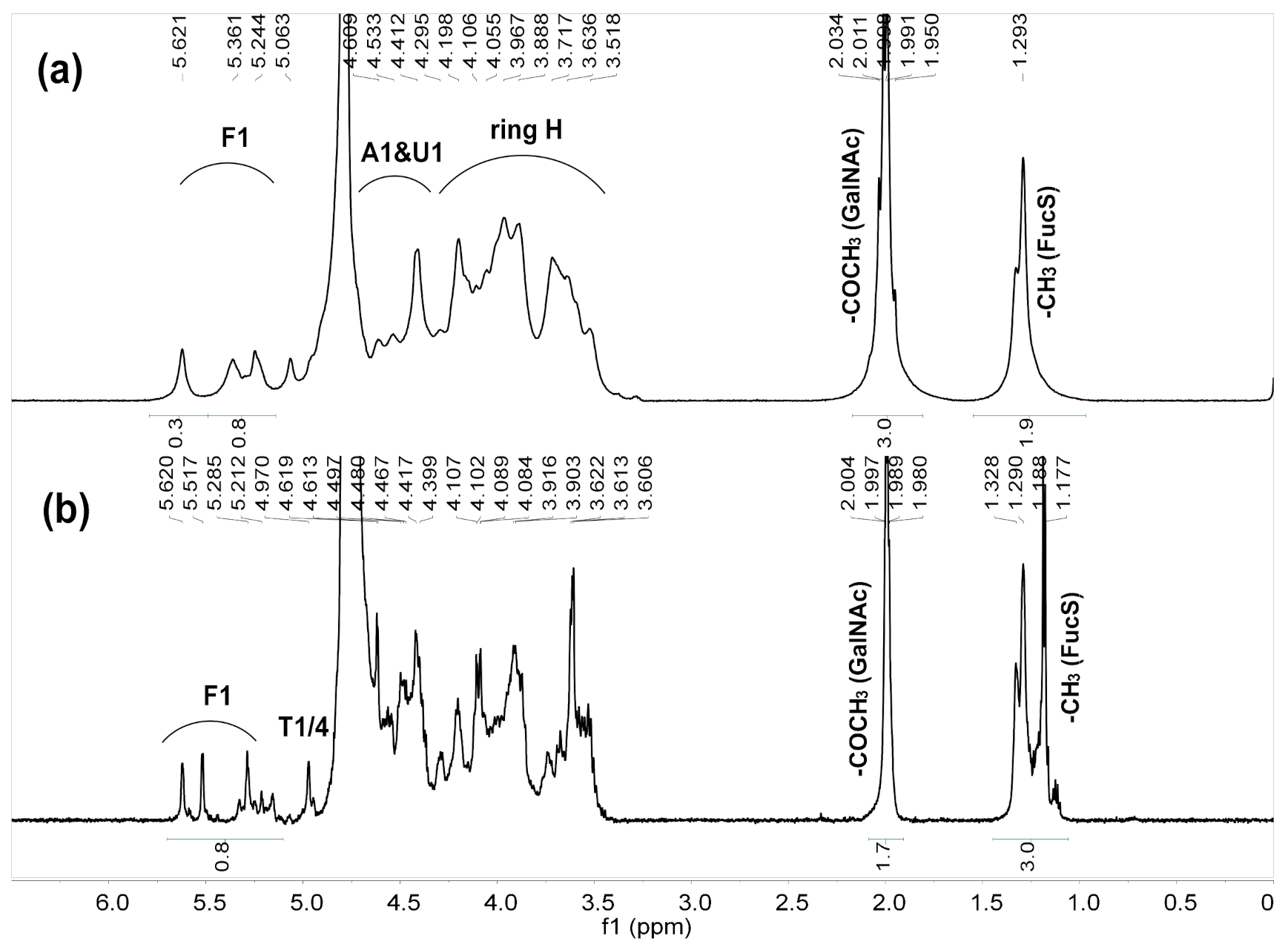
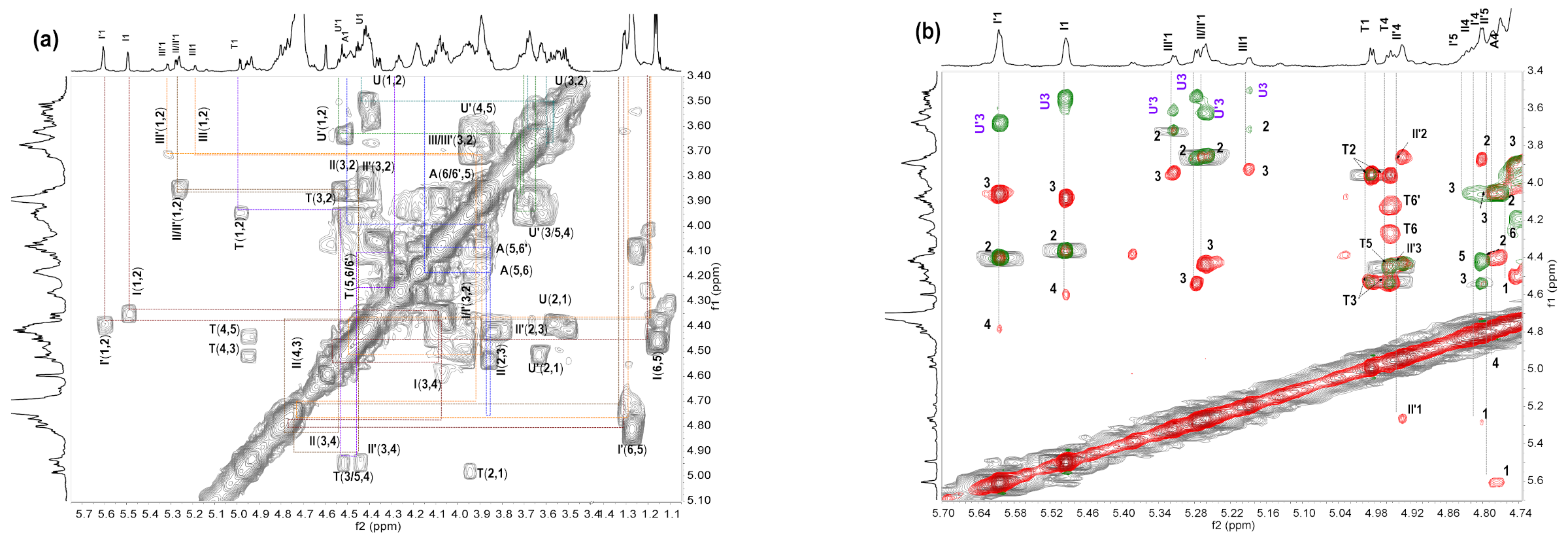
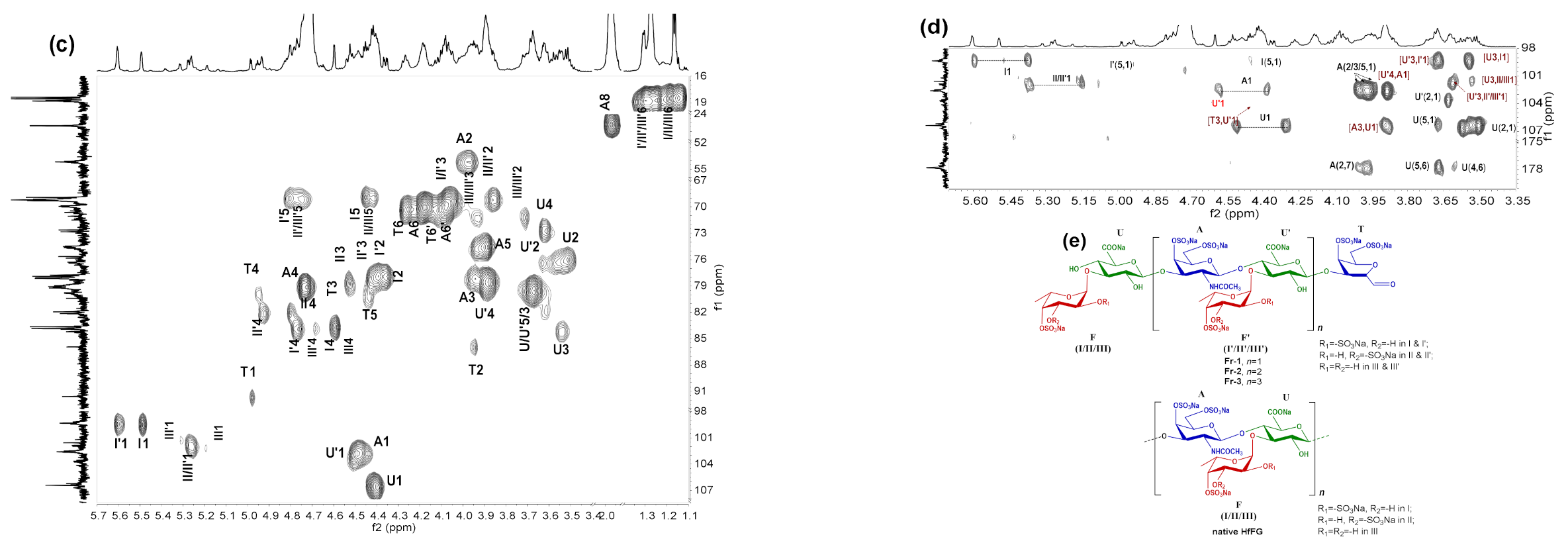
2.5. NMR Analysis of dHfFG, Fr-1, Fr-2 and Fr-3
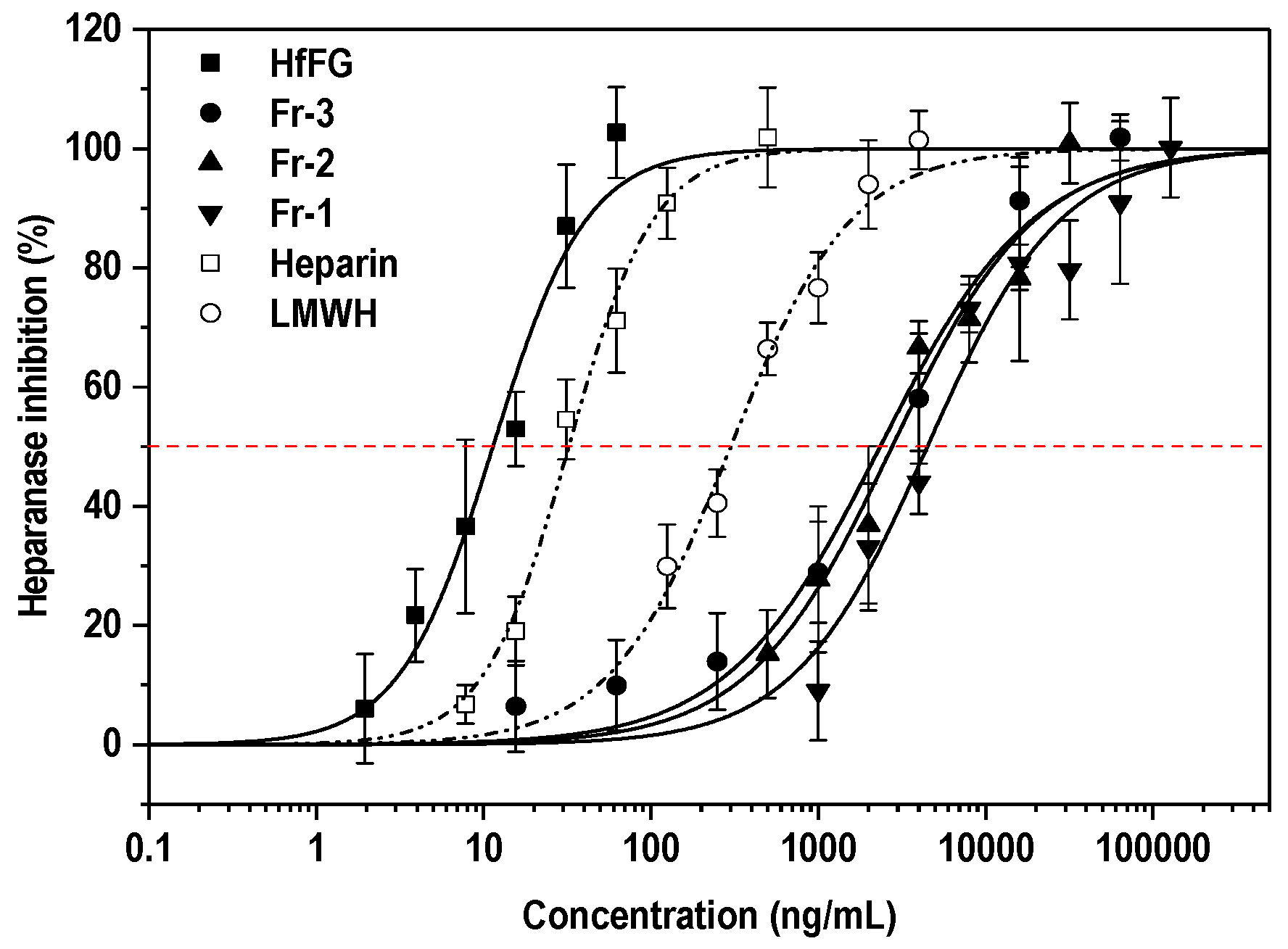
2.6. Heparanase Inhibitory Activity
3. Materials and Methods
3.1. Materials
3.2. Extraction, Isolation and Purification of Polysaccharides
3.3. Preparation of Low-Molecular-Weight HfFG by the Deacetylation–Deaminative Cleavage Method
3.4. Purification of Homogeneous Oligosaccharide Fragments from dHfFG
3.5. General Procedures for Physicochemical Properties Determination
3.6. NMR Analysis
3.7. Heparanase Inhibitory Activities Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purcell, S.W. Value, market preferences and trade of Beche-de-mer from Pacific Island sea cucumbers. PLoS ONE 2014, 9, e95075. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, R.; Wen, Z.Y. Bioactive compounds and biological functions of sea cucumbers as potential functional foods. J. Funct. Foods 2018, 49, 73–84. [Google Scholar] [CrossRef]
- Tian, F.; Zhang, X.W.; Tong, Y.Y.; Yi, Y.H.; Zhang, S.L.; Li, L.; Sun, P.; Lin, L.; Ding, J. PE, a new sulfated saponin from sea cucumber, Exhibits anti-angiogenic and anti-tumor activities in vitro and in vivo. Cancer Biol. Ther. 2005, 4, 874–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsig, L.; Wang, L.; Cavalcante, M.C.; Cardilo-Reis, L.; Ferreira, P.L.; Mourão, P.A.S.; Esko, J.D.; Pavão, M.S. Selectin blocking activity of a fucosylated chondroitin sulfate glycosaminoglycan from sea cucumber. Effect on tumor metastasis and neutrophil recruitment. J. Biol. Chem. 2007, 282, 14984–14991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolmatova, L.S.; Eliseikina, M.G.; Romashina, V.V. Antioxidant enzymatic activity of coelomocytes of the far east sea cucumber Eupentacta fraudatrix. J. Evol. Biochem. Physiol. 2004, 40, 126–135. [Google Scholar] [CrossRef]
- Vieira, R.P.; Mourão, P.A. Occurrence of a unique fucose-branched chondroitin sulfate in the body wall of a sea cucumber. J. Biol. Chem. 1988, 263, 18176–18183. [Google Scholar] [CrossRef]
- Myron, P.; Siddiquee, S.; Azad, S.A. Fucosylated chondroitin sulfate diversity in sea cucumbers: A review. Carbohydr. Polym. 2014, 112, 173–178. [Google Scholar] [CrossRef]
- Pomin, V.H. Holothurian fucosylated chondroitin sulfate. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef] [Green Version]
- Mourão, P.A.; Pereira, M.S.; Pavão, M.S.; Mulloy, B.; Tollefsen, D.M.; Mowinckel, M.C.; Abildgaard, U. Structure and anticoagulant activity of a fucosylated chondroitin sulfate from echinoderm. Sulfated fucose branches on the polysaccharide account for its high anticoagulant action. J. Biol. Chem. 1996, 271, 23973–23984. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Y.; Xu, S.M.; Zhao, J.H.; Kang, H.; Ding, H. Physicochemical characteristics and anticoagulant activities of low molecular weight fractions by free-radical depolymerization of a fucosylated chondroitin sulphate from sea cucumber Thelenata ananas. Food Chem. 2010, 122, 716–723. [Google Scholar] [CrossRef]
- Gao, N.; Wu, M.Y.; Liu, S.; Lian, W.; Li, Z.; Zhao, J.H. Preparation and characterization of O-acylated fucosylated chondroitin sulfate from sea cucumber. Mar. Drugs 2012, 10, 1647–1661. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.G.; Li, G.Y.; Wu, N.; Guo, X.; Liao, N.B.; Ye, X.Q.; Liu, D.H.; Xue, C.H.; Chai, W.G. Sulfation pattern of the fucose branch is important for the anticoagulant and antithrombotic activities of fucosylated chondroitin sulfates. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3054–3066. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. The structure of a fucosylated chondroitin sulfate from the sea cucumber Cucumaria frondosa. Carbohydr. Polym. 2017, 165, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Z.; Chen, J.D.; Lin, K.Z. An acidic mucopolysaccharide isolated from Stichopus japonicus Selenka and some of its physical and chemical properties. Yao Xue Xue Bao 1980, 15, 263–270. [Google Scholar] [PubMed]
- Kariya, Y.; Watabe, S.; Hashimoto, K.; Yoshida, K. Occurrence of chondroitin sulfate E in glycosaminoglycan isolated from the body wall of sea cucumber Stichopus japonicus. J. Biol. Chem. 1990, 265, 5081–5085. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Nifantiev, N.E.; Usov, A.I. New insight on the structural diversity of holothurian fucosylated chondroitin sulfates. Pure Appl. Chem. 2019, 91, 1065–1071. [Google Scholar] [CrossRef]
- Hernández-Flores, A.; Condal, A.; Poot-Salazar, A.; Espinoza-Méndez, J.C. Geostatistical analysis and spatial modeling of population density for the sea cucumbers Isostichopus badionotus and Holothuria floridana on the Yucatan Peninsula, Mexico. Fish. Res. 2015, 172, 114–124. [Google Scholar] [CrossRef]
- Shi, D.; Qi, J.; Zhang, H.; Yang, H.; Yang, Y.; Zhao, X. Comparison of hydrothermal depolymerization and oligosaccharide profile of fucoidan and fucosylated chondroitin sulfate from Holothuria floridana. Int. J. Biol. Macromol. 2019, 132, 738–747. [Google Scholar] [CrossRef]
- Xu, L.; Gao, N.; Xiao, C.; Lin, L.S.; Purcell, S.W.; Wu, M.Y.; Zhao, J.H. Modulating the degree of fucosylation of fucosylated chondroitin sulfate enhances heparin cofactor II-dependent thrombin inhibition. Eur. J. Med. Chem. 2018, 154, 133–143. [Google Scholar] [CrossRef]
- Liu, X.X.; Zhang, Z.X.; Mao, H.; Wang, P.; Zuo, Z.C.; Gao, L.; Shi, X.; Yin, R.H.; Gao, N.; Zhao, J.H. Characterization of the hydrolysis kinetics of fucosylated glycosaminoglycan in mild acid and structures of the resulting oligosaccharides. Mar. Drugs 2020, 18, 286. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Lai, S.S.; Huang, R.; Wu, M.Y.; Gao, N.; Xu, L.; Qin, H.B.; Peng, W.L.; Zhao, J.H. Structure and anticoagulant activity of fucosylated glycosaminoglycan degraded by deaminative cleavage. Carbohydr. Polym. 2013, 98, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.W.; Peng, Y.; Zhou, L.T.; Zheng, W.Q.; Liu, X.X.; Wang, P.; Yuan, Q.X.; Gao, N.; Zhao, L.Y.; Zhao, J.H. Precise structure and anticoagulant activity of fucosylated glycosaminoglycan from Apostichopus japonicus: Analysis of its depolymerized fragments. Mar. Drugs 2019, 17, 195. [Google Scholar] [CrossRef] [Green Version]
- Shang, F.N.; Gao, N.; Yin, R.H.; Lin, L.S.; Xiao, C.; Zhou, L.T.; Li, Z.; Purcell, S.W.; Wu, M.Y.; Zhao, J.H. Precise structures of fucosylated glycosaminoglycan and its oligosaccharides as novel intrinsic factor Xase inhibitors. Eur. J. Med. Chem. 2018, 148, 423–435. [Google Scholar] [CrossRef]
- Yang, W.J.; Chen, D.Y.; He, Z.C.; Zhou, L.T.; Cai, Y.; Mao, H.; Gao, N.; Zuo, Z.L.; Yin, R.H.; Zhao, J.H. NMR characterization and anticoagulant activity of the oligosaccharides from the fucosylated glycosaminoglycan isolated from Holothuria coluber. Carbohydr. Polym. 2020, 233, 115844. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.H.; Zhou, L.T.; Gao, N.; Li, Z.; Zhao, L.Y.; Shang, F.N.; Wu, M.Y.; Zhao, J.H. Oligosaccharides from depolymerized fucosylated glycosaminoglycan: Structures and minimum size for intrinsic factor Xase complex inhibition. J. Biol. Chem. 2018, 293, 14089–14099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naggi, A.; Casu, B.; Perez, M.; Torri, G.; Cassinelli, G.; Penco, S.; Pisano, C.; Giannini, G.; Ishai-Michaeli, R.; Vlodavsky, I. Modulation of the heparanase-inhibiting activity of heparin through selective desulfation, graded N-acetylation, and glycol splitting. J. Biol. Chem. 2005, 280, 12103–12113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi, N.; Maeda, R.; Sesoko, N.; Isono, M.; Ishikawa, S.; Tani, Y.; Takahashi, K.; Oku, T.; Higashi, K.; Onishi, S.; et al. Chondroitin sulfate E blocks enzymatic action of heparanase and heparanase-induced cellular responses. Biochem. Biophys. Res. Commun. 2019, 520, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.X.; Liu, Y.; Hao, J.J.; Zhao, X.L.; Lang, Y.Z.; Fan, F.; Cai, C.; Li, G.Y.; Zhang, L.J.; Yu, G.L. In vivo anti-cancer mechanism of low-molecular-weight fucosylated chondroitin sulfate (LFCS) from sea cucumber Cucumaria frondosa. Molecules 2016, 21, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.T.; Yin, R.H.; Gao, N.; Sun, H.F.; Chen, D.Y.; Cai, Y.; Ren, L.; Yang, L.; Zuo, Z.L.; Zhang, H.B.; et al. Oligosaccharides from fucosylated glycosaminoglycan prevent breast cancer metastasis in mice by inhibiting heparanase activity and angiogenesis. Pharmacol. Res. 2021, 105527. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wu, M.Y.; Xu, L.; Lian, W.; Xiang, J.Y.; Lu, F.; Gao, N.; Xiao, C.; Wang, S.M.; Zhao, J.H. Comparison of physicochemical characteristics and anticoagulant activities of polysaccharides from three sea cucumbers. Mar. Drugs 2013, 11, 399–417. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Yang, W.J.; Yin, R.H.; Zhou, L.T.; Li, Z.K.; Wu, M.Y.; Zhao, J.H. An anticoagulant fucan sulfate with hexasaccharide repeating units from the sea cucumber Holothuria albiventer. Carbohydr. Res. 2018, 464, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.Y.; Wu, M.Y.; Xiao, C.; Yang, L.; Zhou, L.T.; Gao, N.; Li, Z.; Chen, J.; Chen, J.C.; Liu, J.K.; et al. Discovery of an intrinsic tenase complex inhibitor: Pure nonasaccharide from fucosylated glycosaminoglycan. Proc. Natl. Acad. Sci. USA 2015, 112, 8284–8289. [Google Scholar] [CrossRef] [Green Version]
- Chaidedgumjorn, A.; Toyoda, H.; Woo, E.R.; Lee, K.B.; Kim, Y.S.; Toida, T.; Imanari, T. Effect of (1→3)- and (1→4)-linkages of fully sulfated polysaccharides on their anticoagulant activity. Carbohydr. Res. 2002, 337, 925–933. [Google Scholar] [CrossRef]
- Casu, B.; Gennaro, U. A conductimetric method for the determination of sulphate and carboxyl groups in heparin and other mucopolysaccharides. Carbohydr. Res. 1975, 39, 168–176. [Google Scholar] [CrossRef]
- Gao, N.; Lu, F.; Xiao, C.; Yang, L.; Chen, J.; Zhou, K.; Wen, D.D.; Li, Z.; Wu, M.Y.; Jiang, J.M.; et al. β-Eliminative depolymerization of the fucosylated chondroitin sulfate and anticoagulant activities of resulting fragments. Carbohydr. Polym. 2015, 127, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.T.; Gao, N.; Sun, H.F.; Xiao, C.; Yang, L.; Lin, L.S.; Yin, R.H.; Li, Z.; Zhang, H.B.; Ji, X.; et al. Effects of native fucosylated glycosaminoglycan, its depolymerized derivatives on intrinsic factor Xase, coagulation, thrombosis, and hemorrhagic risk. Thromb. Haemostasis. 2020, 120, 607–619. [Google Scholar] [CrossRef] [PubMed]
- He, Z.C.; Zhou, L.T.; Lin, L.S.; Yin, R.H.; Zhao, J.H. Structure and heparanase inhibitory activity of a new glycosaminoglycan from the slug Limacus flavus. Carbohydr. Polym. 2019, 220, 176–184. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue. | Chemical Shift (ppm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| H/C | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| T | →3)-β-d-anTal4S6S-diol | H | 4.983 | 3.949 | 4.523 | 4.952 | 4.433 | 4.263/4.119 | ||
| C | 91.58 | 85.93 | 79.13 | 79.79 | 80.38 | 70.6 | ||||
| U’ | →4)-β-d-GlcA-(1→ | H | 4.484 | 3.623 | 3.673 | 3.893 | 3.673 | |||
| C | 103.64 | 76.38 | 79.79 | 78.13 | 79.32 | 177.34 | ||||
| A | →3)-β-d-GalNAc4S6S-(1→ | H | 4.485 | 3.971 | 3.938 | 4.729 | 3.893 | 4.101/4.184 | ||
| C | 102.58 | 54.05 | 78.9 | 79.05 | 74.53 | 69.88 | 177.76 | 25.3 | ||
| F’ (I’) | α-l-Fuc2S4S-(1→ | H | 5.602 | 4.403 | 4.064 | 4.766 | 4.817 | 1.276 | ||
| C | 99.3 | 77.88 | 67.27 | 83.62 | 69.04 | 18.53 | ||||
| F’ (II’) | α-l-Fuc3S4S-(1→ | H | 5.261 | 3.857 | 4.422 | 4.942 | 4.783 | 1.303 | ||
| C | 101.85 | 69.18 | 77.75 | 81.92 | 68.89 | 18.8 | ||||
| F’(III’) | α-l-Fuc4S-(1→ | H | 5.315 | 3.708 | 3.949 | 4.69 | 4.745 | 1.267 | ||
| C | 102.04 | 71.33 | 71.46 | 83.94 | 68.98 | 18.47 | ||||
| U | d-GlcA-(1→ | H | 4.416 | 3.512 | 3.551 | 3.618 | 3.671 | |||
| C | 106.4 | 76.63 | 84.19 | 72.6 | 79.32 | 177.76 | ||||
| F (I) | α-l-Fuc2S4S-(1→ | H | 5.493 | 4.357 | 4.079 | 4.596 | 4.452 | 1.167 | ||
| C | 99.17 | 77.88 | 69.18 | 83.83 | 68.89 | 18.35 | ||||
| F (II) | α-l-Fuc3S4S-(1→ | H | 5.273 | 3.856 | 4.437 | 4.933 | 4.769 | 1.167 | ||
| C | 101.55 | 69.1 | 78.06 | 82.13 | 69.1 | 18.5 | ||||
| F (III) | α-l-Fuc4S-(1→ | H | 5.187 | 3.711 | 3.93 | 4.511 | 4.408 | 1.144 | ||
| C | 101.19 | 71.33 | 71.46 | 83.55 | 68.98 | 18.35 | ||||
| Sample | Mw (Da) | Specific Rotation [α] | IC50 (ng/mL) | IC50 (nM) |
|---|---|---|---|---|
| HfFG | 47,282 | −50° | 11.4 ± 1.39 | 0.24 ± 2.94 |
| Fr-3 | 3652 a | −45° | 2346 ± 409 | 642.3 ± 112 |
| Fr-2 | 2767 | −32° | 2766 ± 381 | 999.4 ± 138 |
| Fr-1 | 1841 | −44° | 4496 ± 590 | 2440 ± 320 |
| Heparin | ~18,000 | -- | 32.3 ± 2.21 | 1.79 ± 0.12 |
| LMWH | ~4500 | -- | 300 ± 29.4 | 66.67 ± 6.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Guan, R.; Zhou, L.; Zuo, Z.; Tao, X.; Wang, P.; Zhou, Y.; Yin, R.; Zhao, L.; Gao, N.; et al. Structural Characterization and Heparanase Inhibitory Activity of Fucosylated Glycosaminoglycan from Holothuria floridana. Mar. Drugs 2021, 19, 162. https://doi.org/10.3390/md19030162
Shi X, Guan R, Zhou L, Zuo Z, Tao X, Wang P, Zhou Y, Yin R, Zhao L, Gao N, et al. Structural Characterization and Heparanase Inhibitory Activity of Fucosylated Glycosaminoglycan from Holothuria floridana. Marine Drugs. 2021; 19(3):162. https://doi.org/10.3390/md19030162
Chicago/Turabian StyleShi, Xiang, Ruowei Guan, Lutan Zhou, Zhichuang Zuo, Xuelin Tao, Pin Wang, Yanrong Zhou, Ronghua Yin, Longyan Zhao, Na Gao, and et al. 2021. "Structural Characterization and Heparanase Inhibitory Activity of Fucosylated Glycosaminoglycan from Holothuria floridana" Marine Drugs 19, no. 3: 162. https://doi.org/10.3390/md19030162