
Expression and Characterization of a Cold-Adapted Alginate Lyase with Exo/Endo-Type Activity from a Novel Marine Bacterium Alteromonas portus HB161718T

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Screening and Identification of Strain HB161718T
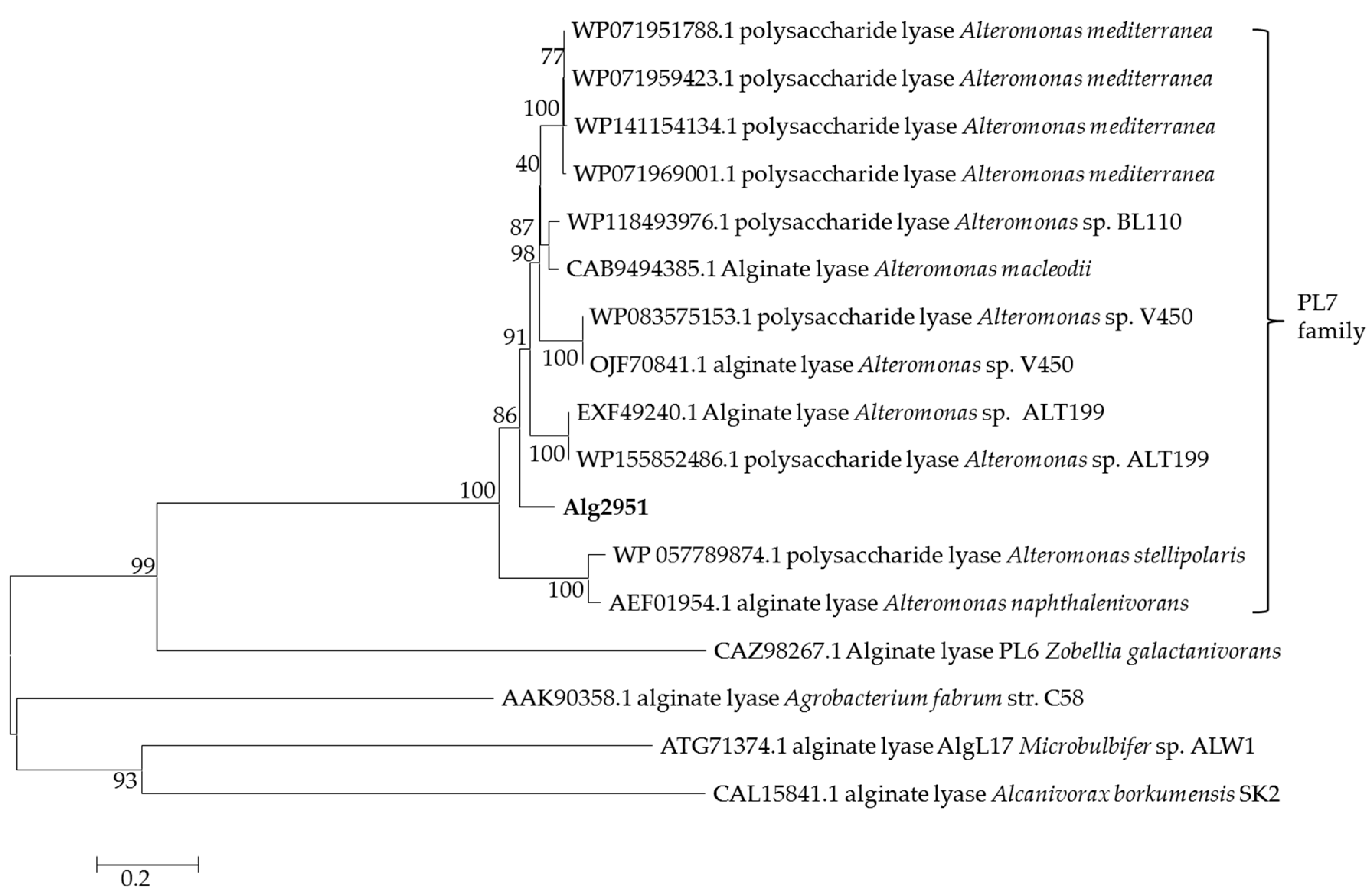
2.2. Genome Sequencing and Bioinformatics Analysis of Alginate Lyase Alg2951
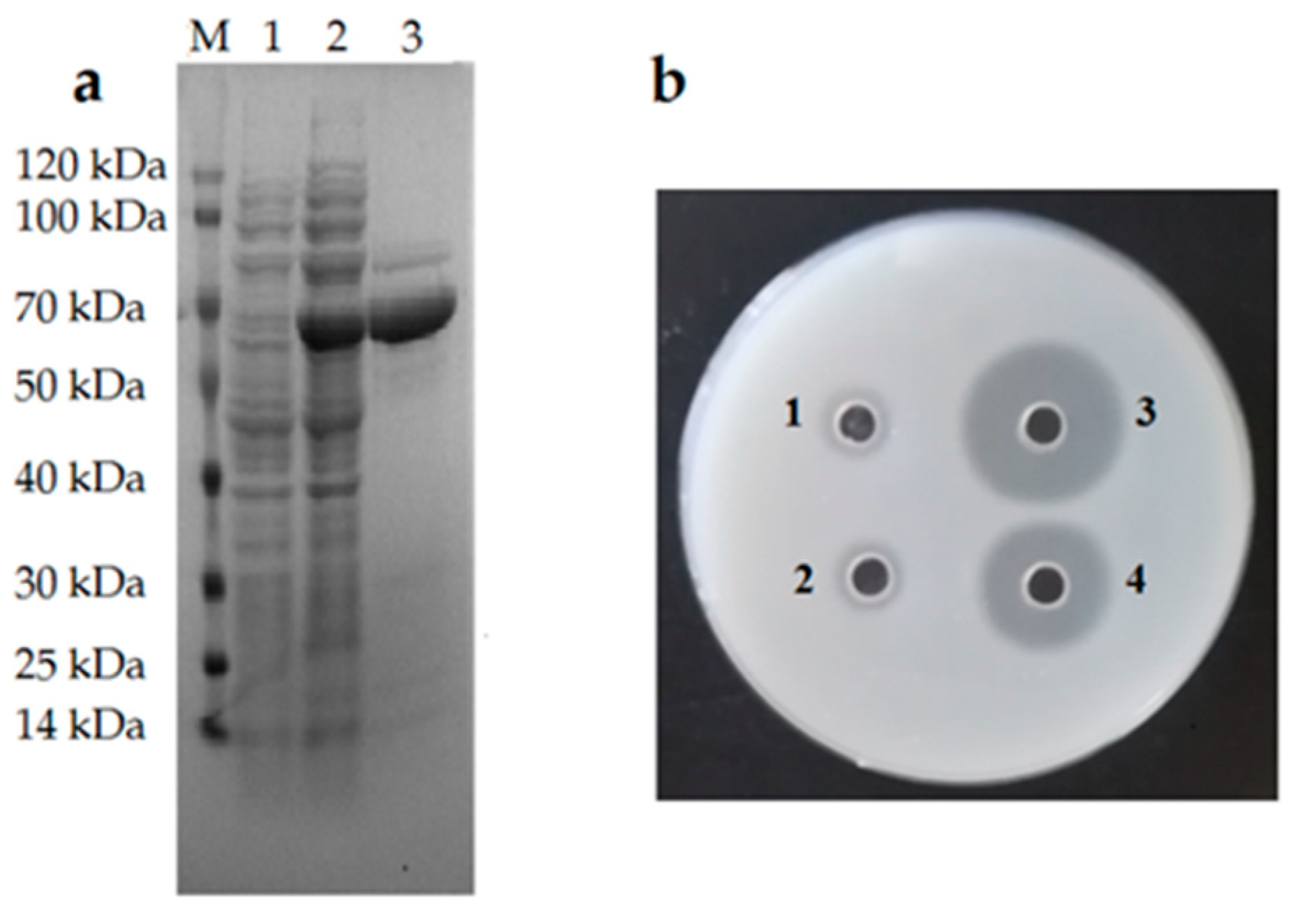
2.3. Expression, Purification, and Activity Detection of the Recombinant Alg2951 (rAlg2951)
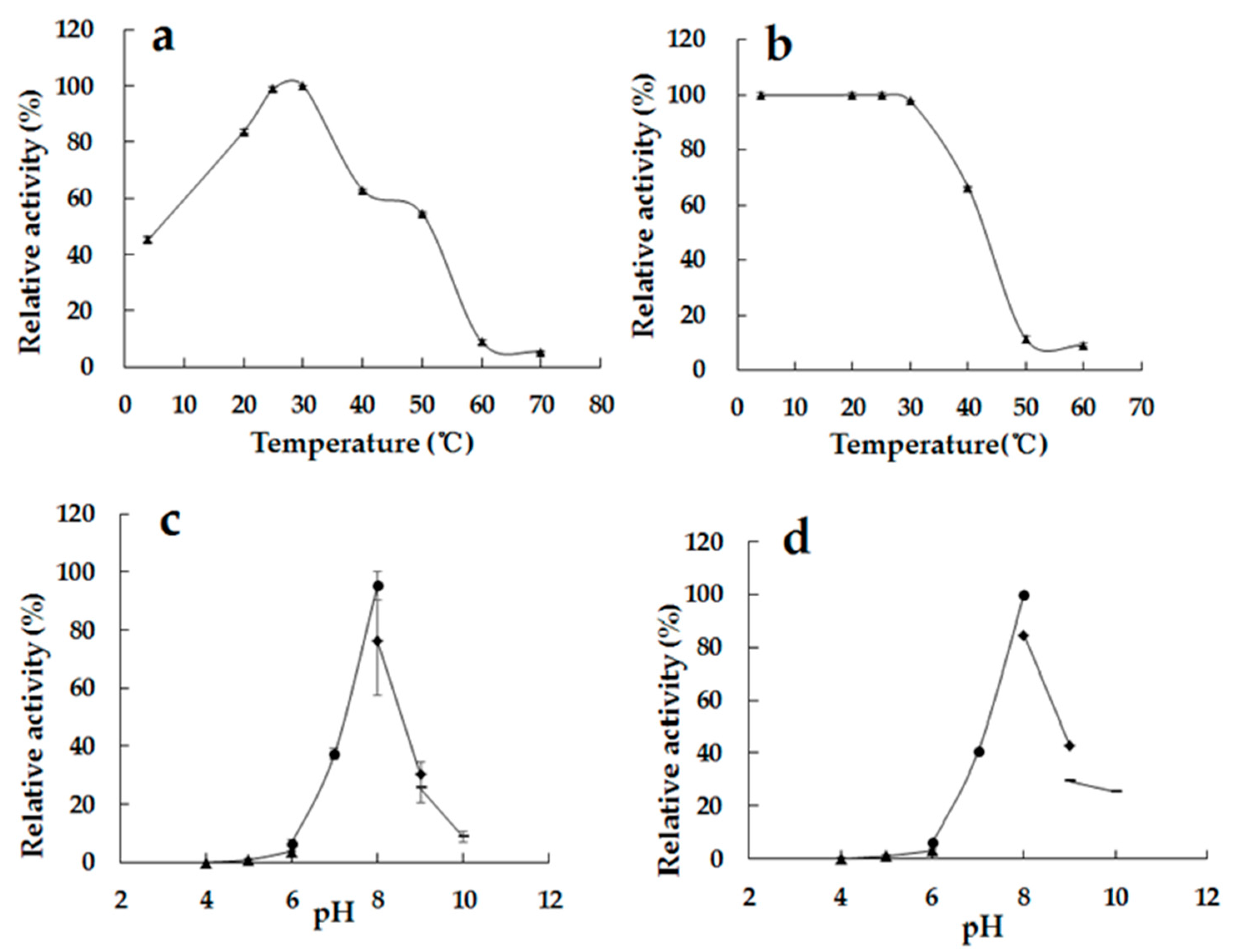
2.4. Temperature and pH Properties of the rAlg2951
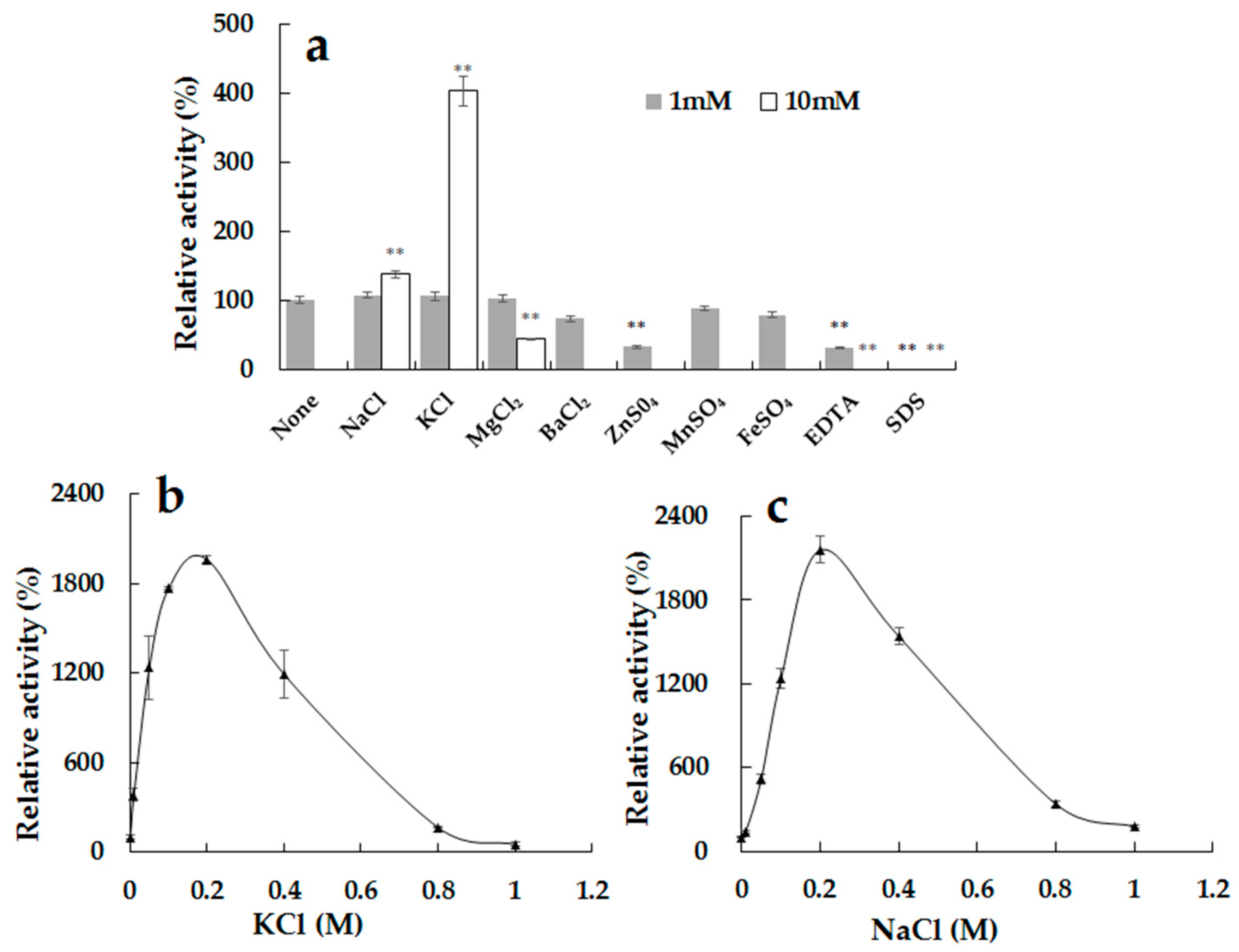
2.5. Effects of Ions and Compounds on the Activity of the Recombinant Alg2951
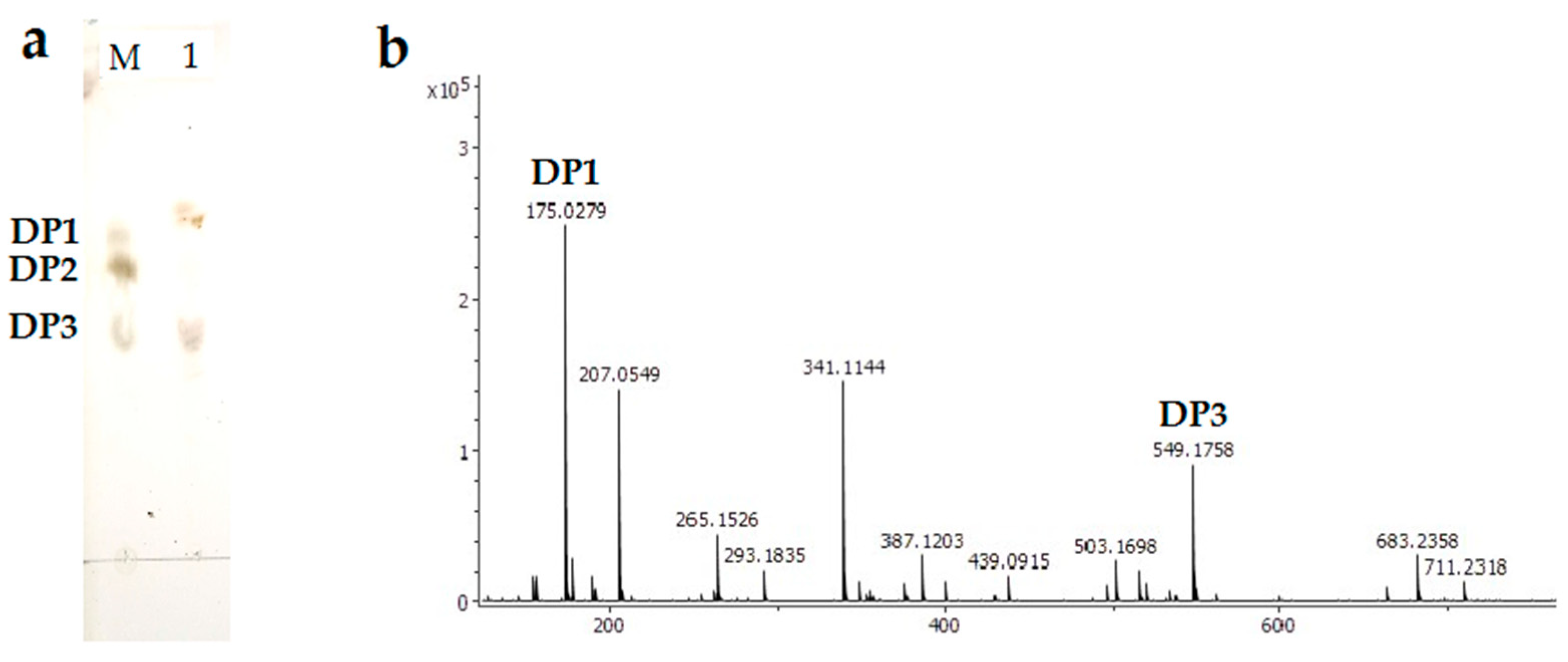
2.6. Analysis of the Degradation Product
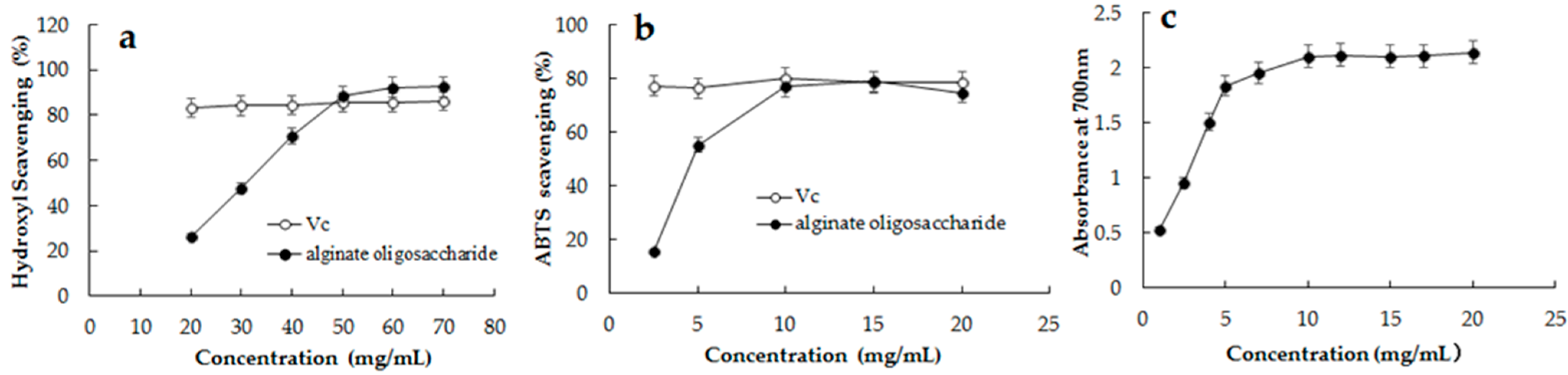
2.7. Antioxidant Activity of the Hydrolysates from rAlg2951-Treated Sodium Alginate
3. Materials and Methods
3.1. Materials, Strains, and Plasmids
3.2. Screening and Identification of Strain HB161718T
3.3. Genome Sequencing and Bioinformatics Analysis of the Alginate Lyase
3.4. Cloning, Expression, Purification, and Activity Detection of the Alginate Lyase Alg2951
3.5. Effects of Temperature and pH Properties Activity and Stability
3.6. Effects of Metal Ions and Compounds on Alg2951 Activity
3.7. Analysis of Alg2951 Reaction Products
3.8. Antioxidant Activity of the Alginate Degradation Products of Alg2951
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, S.; Horn, S.J. Enzymatic saccharification of brown seaweed for production of fermentable sugars. Bioresour. Technol. 2016, 213, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.N.; Edgar, K.J. Alginate derivatization: A review of chemistry, properties and applications. Biomaterials 2012, 33, 3279–3305. [Google Scholar] [CrossRef]
- Zhu, B.W.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusi, S.K.; Khalaj, L.; Ashabi, G.; Kiaei, M.; Khodagholi, F. Alginate oligosaccharide protects against endoplasmic reticulum-and mitochondrialmediated apoptotic cell death and oxidative stress. Biomaterials 2011, 32, 5438–5458. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Yin, H.; Zhao, X.M.; Wang, W.X.; Du, Y.G.; He, A.L.; Sun, K.G. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr. Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef]
- Liu, J.; Yang, S.; Li, X.; Yan, Q.; Reaney, M.J.T.; Jiang, Z. Alginate oligosaccharides: Production, biological activities, and potential applications. Compr. Rev. Food Sci. Food Saf. 2019, 6, 1859–1881. [Google Scholar] [CrossRef] [Green Version]
- Kam, N.; Park, Y.J.; Lee, E.Y.; Kim, H.S. Molecular identification of a polyM-specific alginate lyase from Pseudomonas sp. strain KS-408 for degradation of glycosidic linkages between two mannuronates or mannuronate and guluronate in alginate. Can. J. Microbiol. 2011, 57, 1032–1041. [Google Scholar] [CrossRef]
- Chen, X.L.; Dong, S.; Xu, F.; Dong, F.; Li, P.Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a new cold-adapted and salt-activated polysaccharide lyase family 7 alginate lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120–1128. [Google Scholar] [CrossRef]
- Park, H.H.; Kam, N.; Lee, E.Y.; Kim, H.S. Cloning and characterization of a novel oligoalginate lyase from a newly isolated bacterium Sphingomonas sp. MJ-3. Mar. Biotechnol. 2012, 14, 189–202. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The carbohydrate-aactive enZymes database (CAZy): An expertre source for glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Li, F.L.; Lu, M.; Ji, S.Q.; Wang, B. Biochemical and structural characterization of alginate lyases: An update. Curr. Biotechnol. 2015, 4, 223–239. [Google Scholar]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a novel alginate lyase from Nitratiruptor sp. SB155-2 thriving at deep-sea hydrothermal vents and identification of the residues responsible for its heat stability. J. Bio. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.W.; Ning, L.M.; Jiang, Y.C.; Ge, L. Biochemical characterization and degradation pattern of a novel endo-type bifunctional alginate lyase AlyA from marine bacterium Isoptericola halotolerans. Mar. Drugs 2018, 16, 258. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.H.; Shao, Y.; Jiao, C.; Yang, Q.M.; Weng, H.F.; Xiao, A.F. Characterization and application of an alginate lyase, Aly1281 from marine bacterium Pseudoalteromonas carrageenovora ASY5. Mar. Drugs 2020, 18, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, H.R.; Yoo, J.S.; Choi, Y.L.; Jang, Y.S.; Lee, Y.S. Characterization of an organic solvent-tolerant polysaccharide lyase from Microbulbifer thermotolerans DAU221. Int. J. Biol. Macromol. 2021, 169, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Q.; Mo, K.L.; Li, S.; Sun, D.M.; Zhu, J.; Zou, X.X.; Hu, Y.H.; Bao, S.X. Alteromonas portus sp. nov.; an alginate lyase–excreting marine bacterium. Int. J. Syst. Evol. Microb. 2020, 70, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Q.; Mo, K.L.; Hu, Y.H.; Liu, M.; Zhu, J.; Zou, X.X.; Bao, S.X. Microbulbifer harenosus sp. nov.; an alginate-degrading bacterium isolated from coastal sand. Int. J. Syst. Evol. Microb. 2020, 70, 1639–1643. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Lundqvist, L.; Jam, M.; Jeudy, A.; Barbeyron, T.; Sandström, C.; Michel, G.; Czjzek, M. Comparative characterization of two marine alginate lyases from Zobellia galactanivorans reveals distinct modes of action and exquisite adaptation to their natural substrate. J. Biol. Chem. 2013, 288, 23021–23037. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.W.; Sun, Y.; Ni, F.; Ning, L.M.; Yao, Z. Characterization of a new endo-type alginate lyase from Vibrio sp. NJU-03. Int. J. Biol. Macromol. 2018, 108, 1140–1147. [Google Scholar] [CrossRef]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Ogura, K.; Hashimoto, W.; Mikami, B.; Murata, K. A structural basis for depolymerization of alginate by polysaccharide lyase family-7. J. Mol. Biol. 2005, 352, 11–21. [Google Scholar] [CrossRef]
- Huang, L.S.X.; Zhou, J.G.; Li, X.; Peng, Q.; Lu, H.; Du, Y.G. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Ochi, Y.; Takeuchi, T.; Murata, K.; Kawabata, Y.; Kusakabe, I. A simple method for preparation of poly-mannuronate using poly-guluronate lyase. Biosci. Biotech. Biochem. 1995, 59, 1560–1561. [Google Scholar] [CrossRef]
- Cao, L.; Xie, L.; Xue, X.; Tan, H.; Liu, Y.; Zhou, S. Purification and characterization of alginate lyase from Streptomyces species strain A5 isolated from banana rhizosphere. J. Agr. Food Chem. 2007, 55, 5113–5117. [Google Scholar] [CrossRef] [PubMed]
- Swift, S.M.; Hudgens, J.W.; Heselpoth, R.D.; Bales, P.M.; Nelson, D.C. Characterization of AlgMsp, an alginate lyase from Microbulbifer sp. 6532A. PLoS ONE 2014, 9, e112939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.Y.; Yang, X.M.; Bao, M.M.; Wu, F.; Yu, W.G.; Han, F. Family 13 carbohydrate-binding module of alginate lyase from Agarivorans sp. L11 enhances its catalytic efficiency and thermostability, and alters its substrate preference and product distribution. FEMS Microbiol. Lett. 2015, 362, fnv054. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, S.; Nishiyama, R.; Inoue, A.; Ojima, T. A novel aldo-keto reductase, HdRed, from the pacific abalone Haliotis discus hannai, which reduces alginate-derived 4-deoxy-l-erythro-5-hexoseulose uronic acid to 2-keto-3-deoxy-d-gluconate. J. Biol. Chem. 2015, 290, 30962–30974. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef]
- Zhu, B.W.; Ni, F.; Ning, L.M.; Sun, Y.; Yao, Z. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04. Int. J. Biol. Macromol. 2018, 115, 1063–1070. [Google Scholar] [CrossRef]
- Li, S.Y.; Yang, X.M.; Zhang, L.; Yu, W.G.; Han, F. Cloning, expression, and characterization of a cold-adapted and surfactant-stable alginate lyase from marine bacterium Agarivorans sp. L11. J. Microbiol. Biotechnol. 2015, 25, 681–686. [Google Scholar] [CrossRef]
- Xiao, L.; Feng, H.; Yang, Z.; Lu, X.Z.; Yu, W.G. A novel alginate lyase with high activity on acetylated alginate of Pseudomonas aeruginosa FRD1 from Pseudomonas sp. QD03. World J. Microbiol. Biotechnol. 2006, 22, 81–88. [Google Scholar] [CrossRef]
- Wang, Z.P.; Cao, M.; Li, B.; Ji, X.F.; Zhang, X.Y.; Zhang, Y.Q.; Wang, H.Y. Cloning, secretory expression and characterization of a unique pH-stable and cold-adapted alginate lyase. Mar. Drugs 2020, 18, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, W.F.; Wei, D.; Li, H.; Li, H.; Rahman, M.M.; Shi, J.S.; Xu, Z.H.; Ma, Y.H. Purification and characterisation of a bifunctional alginate lyase from novel Isoptericola halotolerans CGMCC 5336. Carbohydr. Polym. 2013, 98, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhu, Y.M.; Men, Y.; Zeng, Y.; Sun, Y.X. Purification and characterization of a novel alginate lyase from the marine bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.Y.; Wang, L.N.; Chen, X.H.; Zhao, W.W.; Sun, M.; Han, Y.T. Cloning, expression, and biochemical characterization of two new oligoalginate lyases with synergistic degradation capability. Mar. Biotechnol. 2018, 20, 75–86. [Google Scholar] [CrossRef]
- Huang, G.; Wen, S.; Liao, S.; Wang, Q.; Pan, S.; Zhang, R.; Lei, F.; Liao, W.; Feng, J.; Huang, S. Characterization of a bifunctional alginate lyase as a new member of the polysaccharide lyase family 17 from a marine strain BP-2. Biotechnol. Lett. 2019, 41, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Cui, D.; Ma, S.; Chen, W.; Chen, D.; Shen, H. Characterization of a novel PL 17 family alginate lyase with exolytic and endolytic cleavage activity from marine bacterium Microbulbifer sp. SH-1. Int. J. Bio. Macro. 2021, 169, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, A.; Yamasaki, M.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of exotype alginate lyase Atu3025 from Agrobacterium tumefaciens. J. Biol. Chem. 2010, 285, 24519–24528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersters, K.; De Ley, J. The occurrence of the Entner–Doudoroff pathway in bacteria. Antonie Van Leeuwenhoek 1968, 34, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.D.; Guo, Y.X.; Wang, X.X.; Lie, H.B.; Ni, H.; Li, L.J.; Xiao, A.F.; Zhu, Y.B. Molecular cloning and characterization of AlgL17, a new exo-oligoalginate lyase from Microbulbifer sp. ALW1. Protein Expr. Purif. 2019, 161, 17–27. [Google Scholar] [CrossRef]
- Kim, H.T.; Chung, J.H.; Wang, D.; Lee, J.; Woo, H.C.; Choi, I.; Kim, K.H. Depolymerization of alginate into a monomeric sugar acid using Alg17C, an exo-oligoalginate lyase cloned from Saccharophagus degradans 2–40. Appl. Microbiol. Biotechnol. 2012, 93, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Takahashi, M.; Tanaka, R.; Miyake, H.; Shibata, T.; Chow, S.; Kuroda, K.; Ueda, M.; Takeyama, H. Falsirhodobacter sp. alg1 harbors single homologs of endo and exo-type alginate lyases efficient for alginate depolymerization. PLoS ONE 2016, 11, e155537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakata, S.; Murata, K.; Hashimoto1, W.; Kawai, S. Uncovering the reactive nature of 4-deoxy-l-erythro-5-hexoseulose uronate for the utilization of alginate, a promising marine biopolymer. Sci. Rep. 2019, 9, 17147–17157. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, J.; Zhang, X.; Zhou, F.; Lyu, P.; Zhao, Y. Ding, Preparation, composition analysis and antioxidant activities of konjac oligo–glucomannan. Carbohyd. Polym. 2015, 130, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Hisano, T.; Nishimura, M.; Yamashita, T.; Imanaka, T.; Muramatsu, T.; Kimura, A.; Murata, K. A simple method for determination of substrate specificity of alginate lyases. J. Fermen. Bioeng. 1994, 78, 182–184. [Google Scholar] [CrossRef]
- Budi, S.W.; van Tuinen, D.; Arnould, C.; Dumas-Gaudot, E.; Gianinazzi-Pearson, V.; Gianinazzi, S. Hydrolytic enzyme activity of Paenibacillus sp. strain B2 and effects of the antagonistic bacterium on cell integrity of two soil-borne pathogenic fungi. Appl. Soil. Ecol. 2000, 15, 191–199. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2013, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, H. Predicting secretory proteins with SignalP. Methods Mol. Biol. 2017, 1611, 59–73. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Zhu, B.; Hu, F.; Yuan, H.; Sun, Y.; Yao, Z. Biochemical characterization and degradation pattern of a unique pH-stable polyM-specific alginate lyase from newly isolated Serratia marcescens NJ-07. Mar. Drugs 2018, 16, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gülçin, İ.; Bursal, E.; Şehitoğlu, M.H.; Bilsel, M.; Gören, A.C. Polyphenol contents and antioxidant activity of lyophilized aqueous extract of propolis from Erzurum, Turkey. Food Chem. Toxicol. 2010, 48, 2227–2238. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Li, S.; Bao, S.; Mo, K.; Sun, D.; Hu, Y. Expression and Characterization of a Cold-Adapted Alginate Lyase with Exo/Endo-Type Activity from a Novel Marine Bacterium Alteromonas portus HB161718T. Mar. Drugs 2021, 19, 155. https://doi.org/10.3390/md19030155
Huang H, Li S, Bao S, Mo K, Sun D, Hu Y. Expression and Characterization of a Cold-Adapted Alginate Lyase with Exo/Endo-Type Activity from a Novel Marine Bacterium Alteromonas portus HB161718T. Marine Drugs. 2021; 19(3):155. https://doi.org/10.3390/md19030155
Chicago/Turabian StyleHuang, Huiqin, Shuang Li, Shixiang Bao, Kunlian Mo, Dongmei Sun, and Yonghua Hu. 2021. "Expression and Characterization of a Cold-Adapted Alginate Lyase with Exo/Endo-Type Activity from a Novel Marine Bacterium Alteromonas portus HB161718T" Marine Drugs 19, no. 3: 155. https://doi.org/10.3390/md19030155