
Enzyme-Digested Peptides Derived from Lates calcarifer Enhance Wound Healing after Surgical Incision in a Murine Model

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
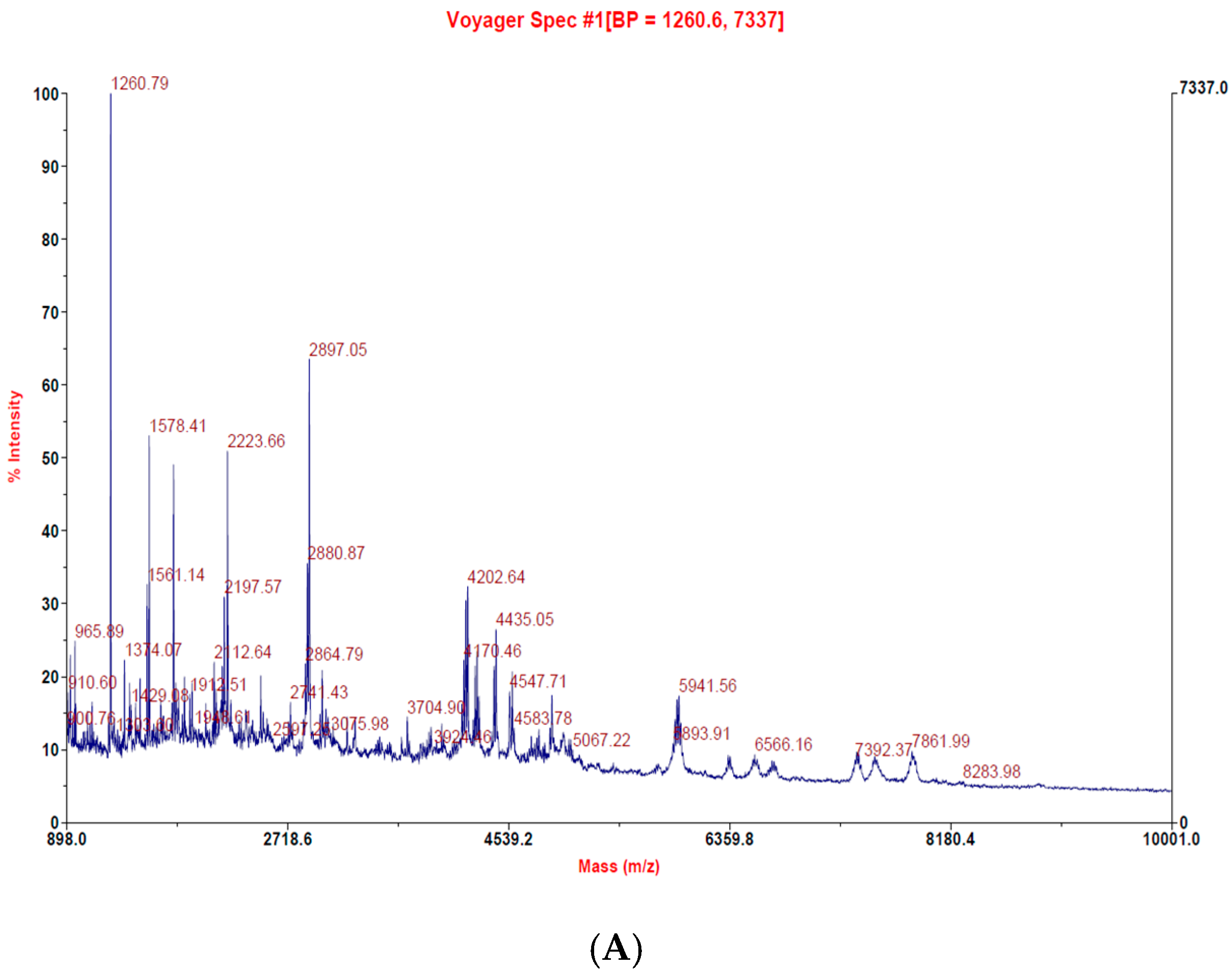
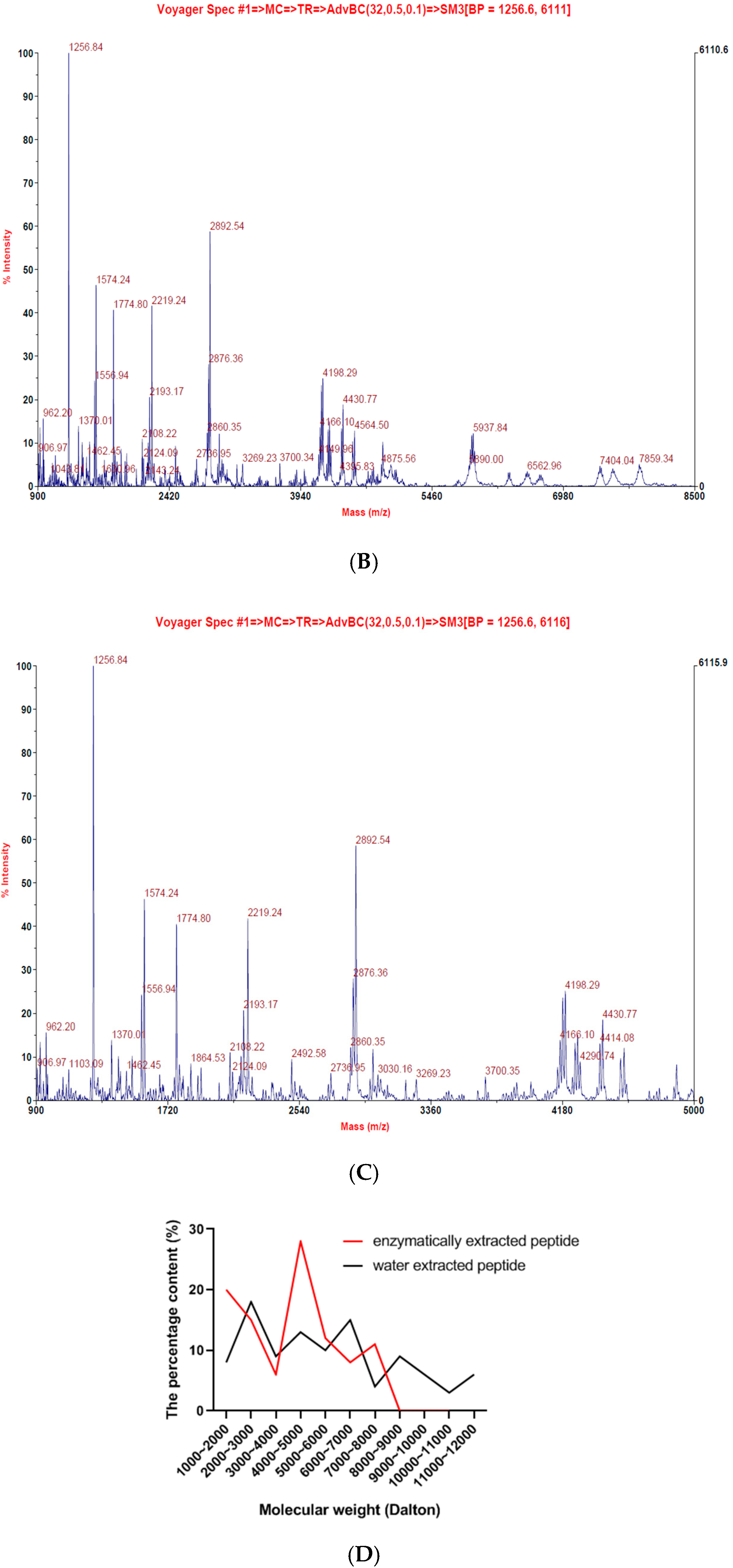
2.1. Lates Calcarifer-Extracted Peptide (LCEP) Molecular Weight Analysis and Comparison
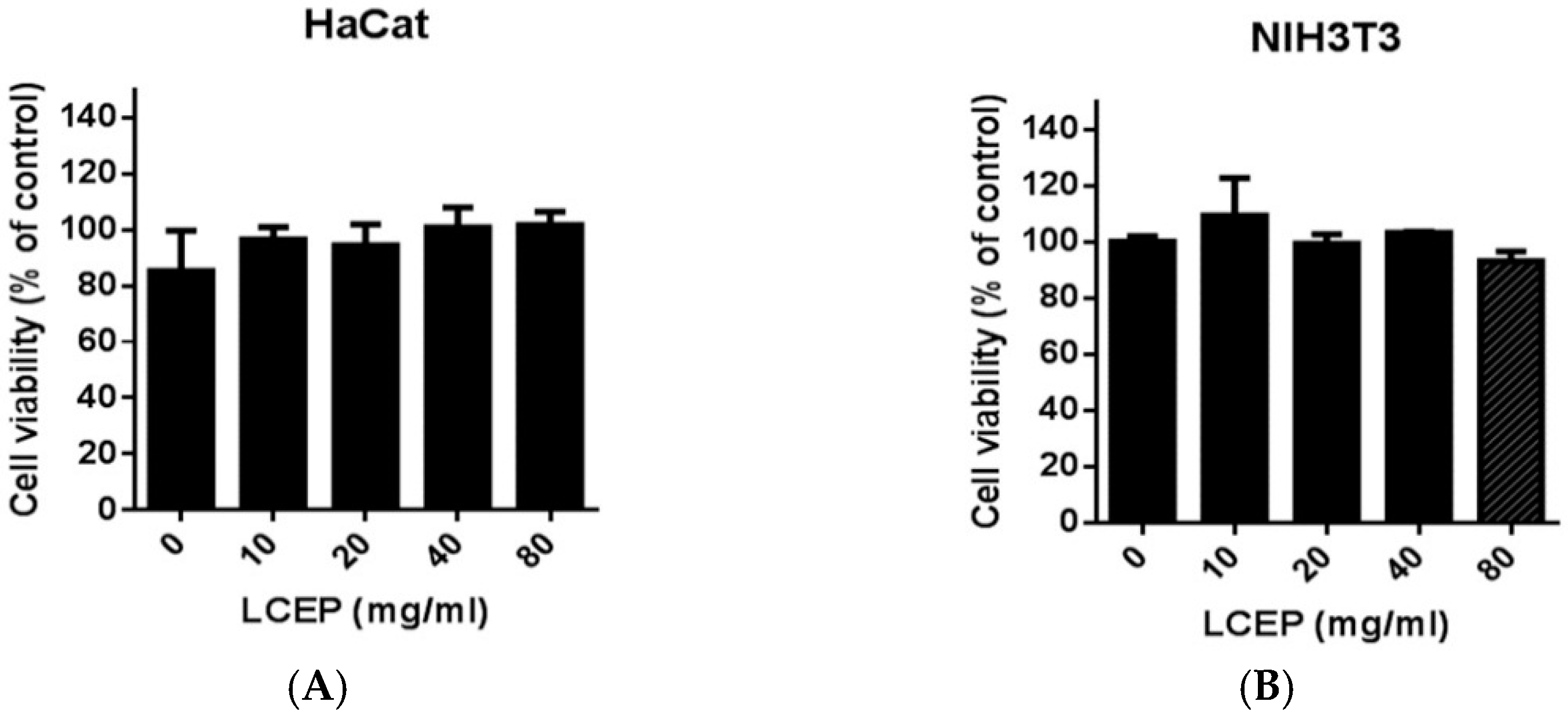
2.2. In Vitro Toxicity Analysis of LCEP
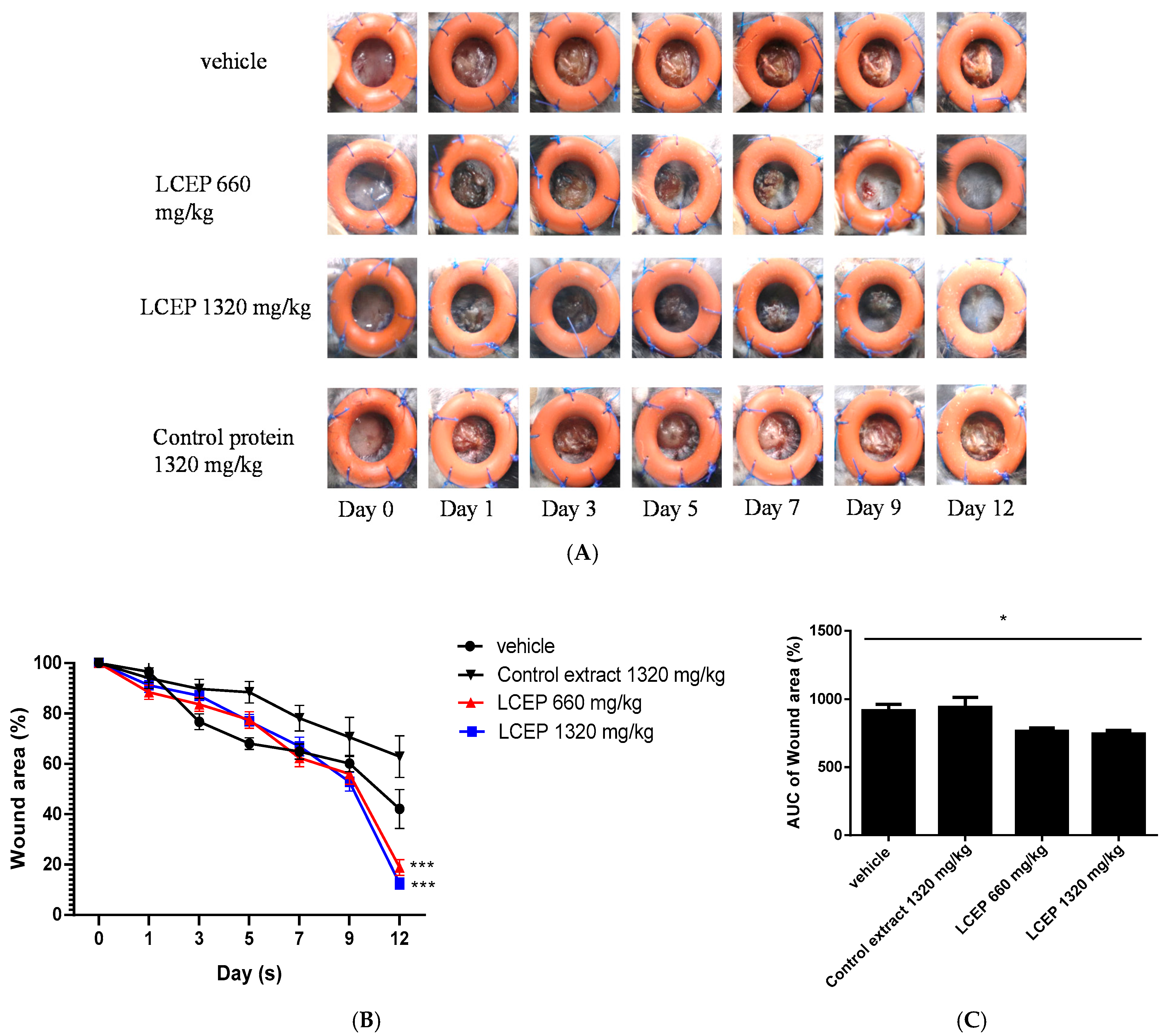
2.3. In Vivo Evaluation of LCEP On Wound Healing Process
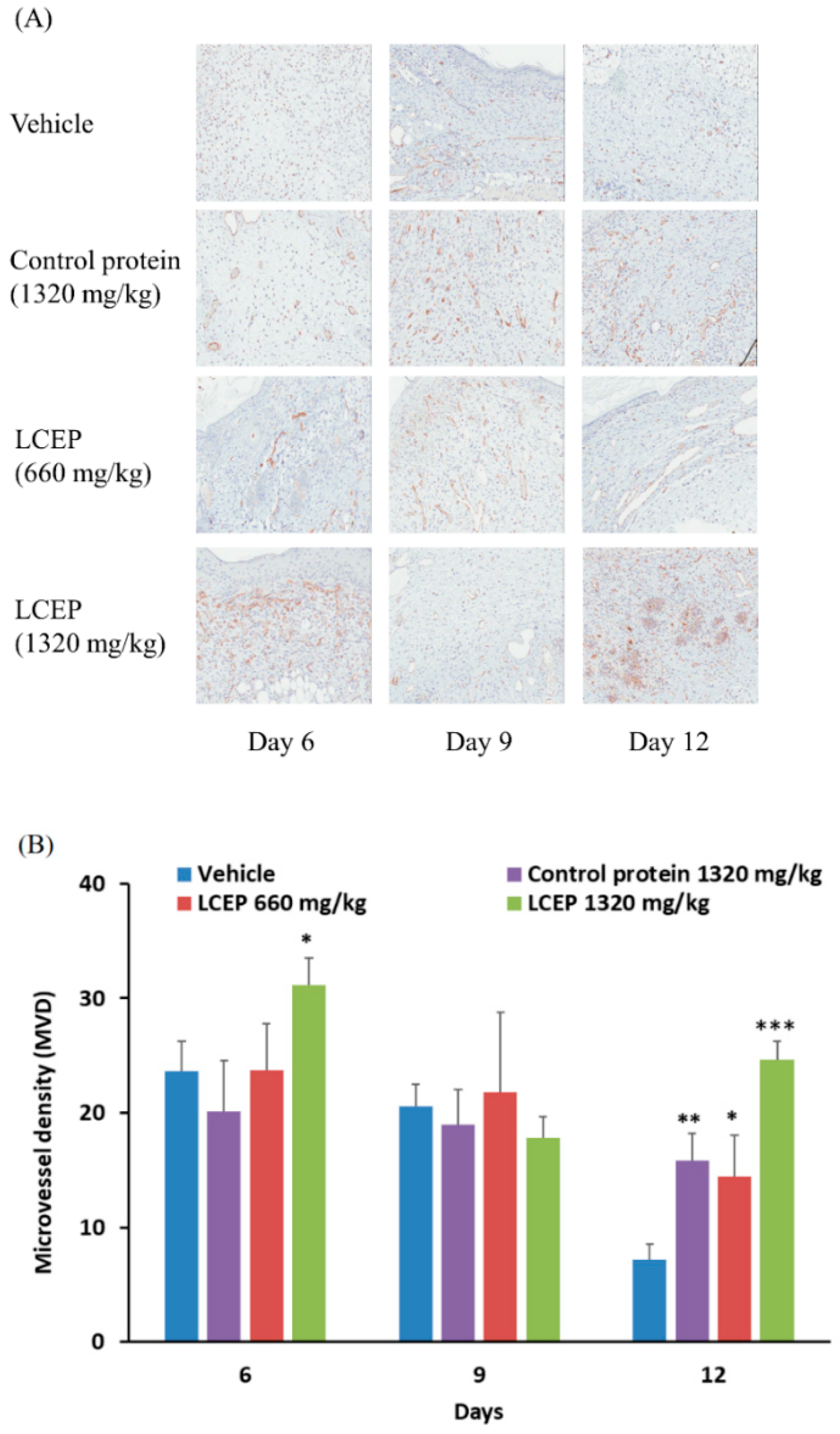
2.4. The Effects of LCEP on the Formation of Microvessel
3. Discussion
4. Materials and Methods
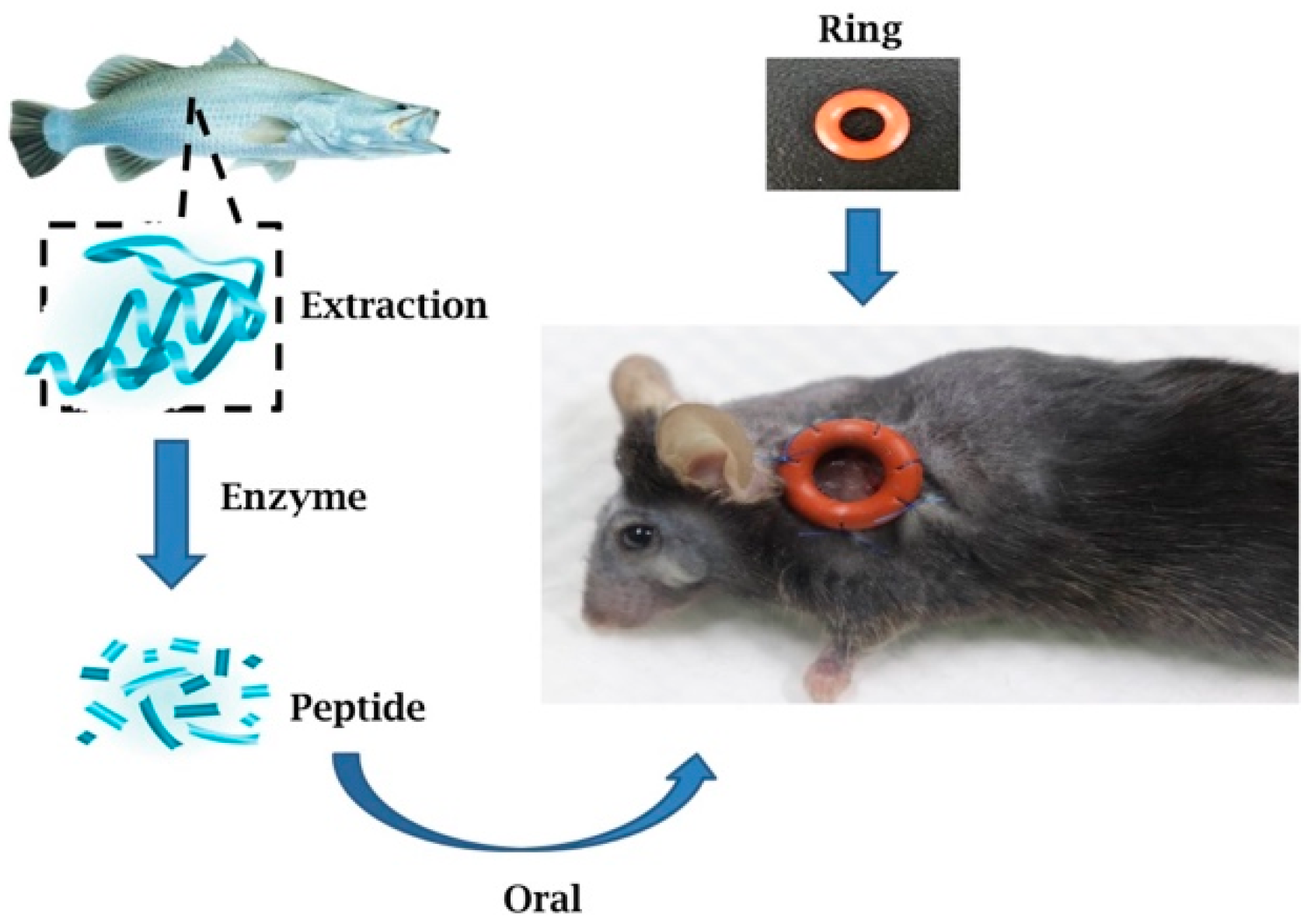
4.1. Lates Calcarifer Sample Preparation
4.2. Lates Calcarifer Molecular Weight Assay
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Animals
4.6. Splints
4.7. Morphological Analysis
4.8. Immunohistochemical Staining
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atef, M.; Mahdi Ojagh, S. Health benefits and food applications of bioactive compounds from fish byproducts: A review. J. Funct. Foods 2017, 35, 673–681. [Google Scholar] [CrossRef]
- Kwan, S.H.; Abdul Aziz, N.H.; Ismail, M.N. Bioactive Proteins in Channa striata Promote Wound Healing through Angiogenesis and Cell Proliferation. Protein Pept. Lett. 2020, 27, 48–59. [Google Scholar] [CrossRef]
- Chen, J.; Gao, K.; Liu, S.; Wang, S.; Elango, J.; Bao, B.; Dong, J.; Liu, N.; Wu, W. Fish collagen surgical compress repairing characteristics on wound healing process in vivo. Mar. Drugs 2019, 17, 33. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Jayachandran, M.; Xu, B.; Yu, Z. Sea bass (Lateolabrax maculatus) accelerates wound healing: A transition from inflammation to proliferation. J. Ethnopharmacol. 2019, 236, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Sadovy, Y.; Cornish, A.S. Reef fishes of Hong Kong; Hong Kong University Press: Hong Kong, China, 2000; 321p. [Google Scholar]
- Wang, Z.T.; Nowacek, D.P.; Akamatsu, T.; Wang, K.X.; Liu, J.C.; Duan, G.-Q.; Cao, H.J.; Wang, D. Diversity of fish sound types in the Pearl River Estuary, China. PeerJ 2017, 5, e3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Liang, Y. Li Shizhen and the Grand Compendium of Materia Medica. J. Tradit. Chin. Med. Sci. 2015, 2, 215–216. [Google Scholar] [CrossRef] [Green Version]
- Munekata, P.E.; Pateiro, M.; Domínguez, R.; Zhou, J.; Barba, F.J.; Lorenzo, J.M. Nutritional Characterization of Sea Bass Processing By-Products. Biomolecules 2020, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, J.S.; Reinhart, P.; Pangalos, M.N. Current concepts in therapeutic strategies targeting cognitive decline and disease modification in Alzheimer’s disease. NeuroRx 2005, 2, 612–626. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.P.; Chen, P.C.; Wang, S.W.; Fong, Y.C.; Tsai, C.H.; Tsai, F.J.; Chung, J.G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; et al. Plumbagin suppresses endothelial progenitor cell-related angiogenesis in vitro and in vivo. J. Funct. Foods 2019, 52, 537–544. [Google Scholar] [CrossRef]
- Song, Y.; Wu, C.; Zhang, X.; Bian, W.; Liu, N.; Yin, S.; Yang, M.; Luo, M.; Tang, J.; Yang, X. A short peptide potentially promotes the healing of skin wound. Biosci. Rep. 2019, 39, BSR20181734. [Google Scholar] [CrossRef]
- Sheets, A.R.; Demidova-Rice, T.N.; Shi, L.; Ronfard, V.; Grover, K.V.; Herman, I.M. Identification and Characterization of Novel Matrix-Derived Bioactive Peptides: A Role for Collagenase from Santyl Ointment in Post-Debridement Wound Healing? PLoS ONE 2016, 11, e0159598. [Google Scholar]
- Demidova-Rice, T.N.; Geevarghese, A.; Herman, I.M. Bioactive peptides derived from vascular endothelial cell extracellular matrices promote microvascular morphogenesis and wound healing in vitro. Wound Repair Regen. 2011, 19, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Yang, P.; Zhou, C.; Li, S.; Hong, P. Marine Collagen Peptides from the Skin of Nile Tilapia (Oreochromis niloticus): Characterization and Wound Healing Evaluation. Mar. Drugs 2017, 15, 102. [Google Scholar] [CrossRef]
- Yang, F.; Qin, X.; Zhang, T.; Zhang, C.; Lin, H. Effect of Oral Administration of Active Peptides of Pinctada Martensii on the Repair of Skin Wounds. Mar. Drugs 2019, 17, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abercrombie, M.; James, D.W.; Newcombe, J.F. Wound contraction in rabbit skin, studied by splinting the wound margins. J. Anat. 1960, 94, 170. [Google Scholar] [PubMed]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Ahn, S.T.; Mustoe, T.A. Effects of ischemia on ulcer wound healing: A new model in the rabbit ear. Ann. Plast. Surg. 1990, 24, 17–23. [Google Scholar] [CrossRef]
- Falanga, V.; Schrayer, D.; Cha, J.; Butmarc, J.; Carson, P.; Roberts, A.B.; Kim, S.J. Full-thickness wounding of the mouse tail as a model for delayed wound healing: Accelerated wound closure in Smad3 knock-out mice. Wound Repair Regen. 2004, 12, 320–326. [Google Scholar] [CrossRef]
- Reid, R.R.; Sull, A.C.; Mogford, J.E.; Roy, N.; Mustoe, T.A. A novel murine model of cyclical cutaneous ischemia-reperfusion injury. J. Surg. Res. 2004, 116, 172–180. [Google Scholar] [CrossRef]
- Galiano, R.D.; Michaels, J.; Dobryansky, M.; Levine, J.P.; Gurtner, G.C. Quantitative and reproducible murine model of excisional wound healing. Wound Repair Regen. 2004, 12, 485–492. [Google Scholar] [CrossRef]
- Lindblad, W.J. Considerations for selecting the correct animal model for dermal wound-healing studies. J. Biomater. Sci. Polym. Ed. 2008, 19, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.W.; Sorkin, M.; Glotzbach, J.P.; Longaker, M.T.; Gurtner, G.C. Surgical approaches to create murine models of human wound healing. J. Biomed. Biotechnol. 2010, 2011, 969618. [Google Scholar] [CrossRef] [PubMed]
- Zomer, H.D.; Trentin, A.G. Skin wound healing in humans and mice: Challenges in translational research. J. Dermatol. Sci. 2018, 90, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimi, S.; De Francesco, F.; Ferraro, G.A.; Riccio, M.; Hara, S. A novel skin splint for accurately mapping dermal remodeling and epithelialization during wound healing. J. Cell. Physiol. 2017, 232, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, J.S.; Papadopoulos, J.; Wook Kim, S.; Maya, M.; Zhang, F.; He, J.; Fan, D.; Langley, R.; Fidler, I.J. Circulating Monocytes Expressing CD31: Implications for Acute and Chronic Angiogenesis. Am. J. Pathol. 2009, 174, 1972–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune regulation of skin wound healing: Mechanisms and novel therapeutic targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Zimmermann, A.; Truss, F.; Zimmermann, A.; Truss, F. The effect of antibiotic drugs on wound-healing. Urol. Res. 1974, 2, 73–77. [Google Scholar] [CrossRef]
- Ewa, K.S.; Besser, M.; Terberger, N.; Koester, V.; Bachmann, H.S.; Severing, A.L. Side effects of frequently used oral antidiabetics on wound healing in vitro. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 371–380. [Google Scholar]
- Kemaloğlu, C.A.; Günay, G.K.; Perçin, D.; Deniz, K.J. An unpredicted side effect of prophylactic antibiotic use. J. Chemother. 2014, 26, 154–158. [Google Scholar] [CrossRef]
- Huang, T.Y.; Peng, S.F.; Huang, Y.P.; Tsai, C.H.; Tsai, F.J.; Huang, C.Y.; Tang, C.H.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Combinational treatment of all-trans retinoic acid (ATRA) and bisdemethoxycurcumin (BDMC)-induced apoptosis in liver cancer Hep3B cells. J. Food Biochem. 2020, 44, e13122. [Google Scholar] [CrossRef]
- Lin, C.C.; Chen, K.B.; Tsai, C.H.; Tsai, F.J.; Huang, C.Y.; Tang, C.H.; Yang, J.S.; Hsu, Y.M.; Peng, S.F.; Chung, J.G. Casticin inhibits human prostate cancer DU 145 cell migration and invasion via Ras/Akt/NF-κB signaling pathways. J. Food Biochem. 2019, 43, e12902. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.P.; Wu, Y.C.; Chen, B.C.; Liu, S.C.; Li, T.M.; Huang, W.C.; Hsu, C.J.; Tang, C.H. Soya-cerebroside reduces interleukin production in human rheumatoid arthritis synovial fibroblasts by inhibiting the ERK, NF-kappa B and AP-1 signalling pathways. J. Biol. Chem. 2020, 31, 740–750. [Google Scholar]
- Liu, S.C.; Tsai, C.H.; Wu, T.Y.; Tsai, C.H.; Tsai, F.J.; Chung, J.G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Soya-cerebroside reduces IL-1β induced MMP-1 production in chondrocytes and inhibits cartilage degradation: Implications for the treatment of osteoarthritis. J. Biol. Chem. 2019, 30, 620–632. [Google Scholar] [CrossRef]
- Lee, H.P.; Wang, S.W.; Wu, Y.C.; Lin, L.W.; Tsai, F.J.; Yang, J.S.; Li, T.M.; Tang, C.H. Soya-cerebroside inhibits VEGF-facilitated angiogenesis in endothelial progenitor cells. J. Biol. Chem. 2020, 31, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.P.; Wang, S.W.; Wu, Y.C.; Tsai, C.H.; Tsai, F.J.; Chung, J.-G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Glucocerebroside reduces endothelial progenitor cell-induced angiogenesis. J. Biol. Chem. 2019, 30, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, Y.; Zou, Z.; Yang, M.; Wu, C.; Su, Y.; Tang, J.; Yang, X. OM-LV20, a novel peptide from odorous frog skin, accelerates wound healing in vitro and in vivo. Chem. Biol. Drug Des. 2018, 91, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Hardwicke, J.; Schmaljohann, D.; Boyce, D.; Thomas, D. Epidermal growth factor therapy and wound healing—Past, present and future perspectives. Surgeon 2008, 6, 172–177. [Google Scholar] [CrossRef]
- Meier, N.T.; Haslam, I.S.; Pattwell, D.M.; Zhang, G.Y.; Emelianov, V.; Paredes, R.; Debus, S.; Augustin, M.; Funk, W.; Amaya, E. Thyrotropin-releasing hormone (TRH) promotes wound re-epithelialisation in frog and human skin. PLoS ONE 2013, 8, e73596. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.R.; Voyles, J.; Cook, D.I.; Dinudom, A. Frog skin epithelium: Electrolyte transport and chytridiomycosis. Int. J. Biochem. Cell Biol. 2012, 44, 431–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, C.D.; Hu, M.S.; Leavitt, T.; Barnes, L.A.; Cheung, A.T.; Malhotra, S.; Lorenz, H.P.; Delp, S.L.; Quake, S.R.; Longaker, M.T. Sanativo Wound Healing Product Does Not Accelerate Reepithelialization in a Mouse Cutaneous Wound Healing Model. Plast. Reconstr. Surg. 2017, 139, 343–352. [Google Scholar] [CrossRef]
- Schlüter, A.; Weller, P.; Kanaan, O.; Nel, I.; Heusgen, L.; Höing, B.; Haßkamp, P.; Zander, S.; Mandapathil, M.; Dominas, N. CD31 and VEGF are prognostic biomarkers in early-stage, but not in late-stage, laryngeal squamous cell carcinoma. BMC Cancer 2018, 18, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felician, F.F.; Yu, R.H.; Li, M.Z.; Li, C.J.; Chen, H.Q.; Jiang, Y.; Tang, T.; Qi, W.Y.; Xu, H.M. The wound healing potential of collagen peptides derived from the jellyfish Rhopilema esculentum. Chin. J. Traumatol. 2019, 22, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.M.; Hsu, Y.M.; Ying, M.C.; Tsai, F.J.; Tsai, C.H.; Chung, J.G.; Yang, J.S.; Tang, C.H.; Cheng, L.Y.; Su, P.H. High-density lipoprotein ameliorates palmitic acid-induced lipotoxicity and oxidative dysfunction in H9c2 cardiomyoblast cells via ROS suppression. Nutr. Metab. 2019, 16, 36. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-A.; Chu, P.-Y.; Ma, W.-L.; Cheng, W.-C.; Chan, S.-T.; Yang, J.-C.; Wu, Y.-C. Enzyme-Digested Peptides Derived from Lates calcarifer Enhance Wound Healing after Surgical Incision in a Murine Model. Mar. Drugs 2021, 19, 154. https://doi.org/10.3390/md19030154
Lin Y-A, Chu P-Y, Ma W-L, Cheng W-C, Chan S-T, Yang J-C, Wu Y-C. Enzyme-Digested Peptides Derived from Lates calcarifer Enhance Wound Healing after Surgical Incision in a Murine Model. Marine Drugs. 2021; 19(3):154. https://doi.org/10.3390/md19030154
Chicago/Turabian StyleLin, Yen-An, Pei-Yi Chu, Wen-Lung Ma, Wei-Chung Cheng, Shu-Ting Chan, Juan-Cheng Yang, and Yang-Chang Wu. 2021. "Enzyme-Digested Peptides Derived from Lates calcarifer Enhance Wound Healing after Surgical Incision in a Murine Model" Marine Drugs 19, no. 3: 154. https://doi.org/10.3390/md19030154