
Inhibitory Effect of Astaxanthin on Testosterone-Induced Benign Prostatic Hyperplasia in Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
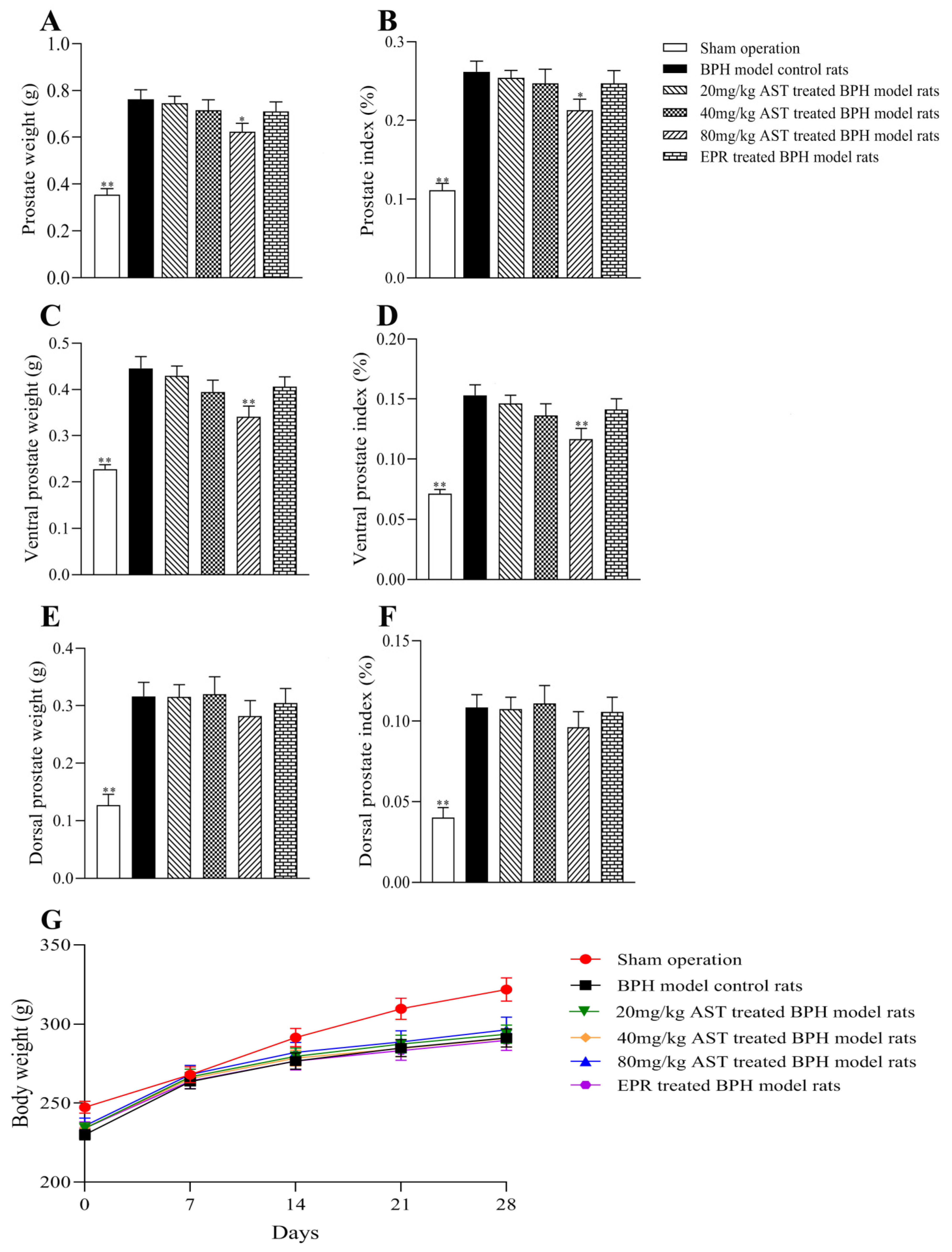
2.1. Effect of AST on Prostate Weight
2.2. Effect of AST on the Body Weight and Prostate Index
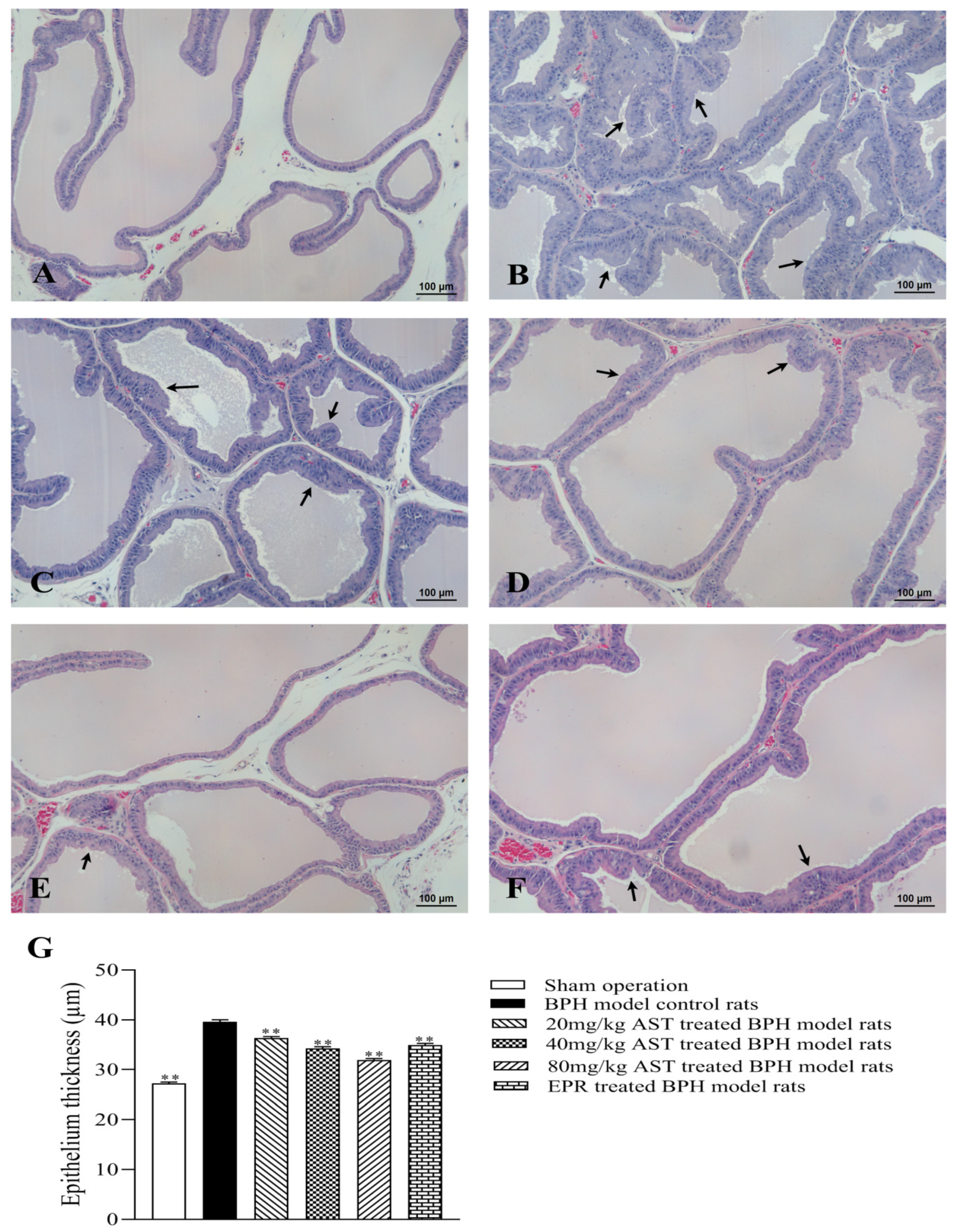
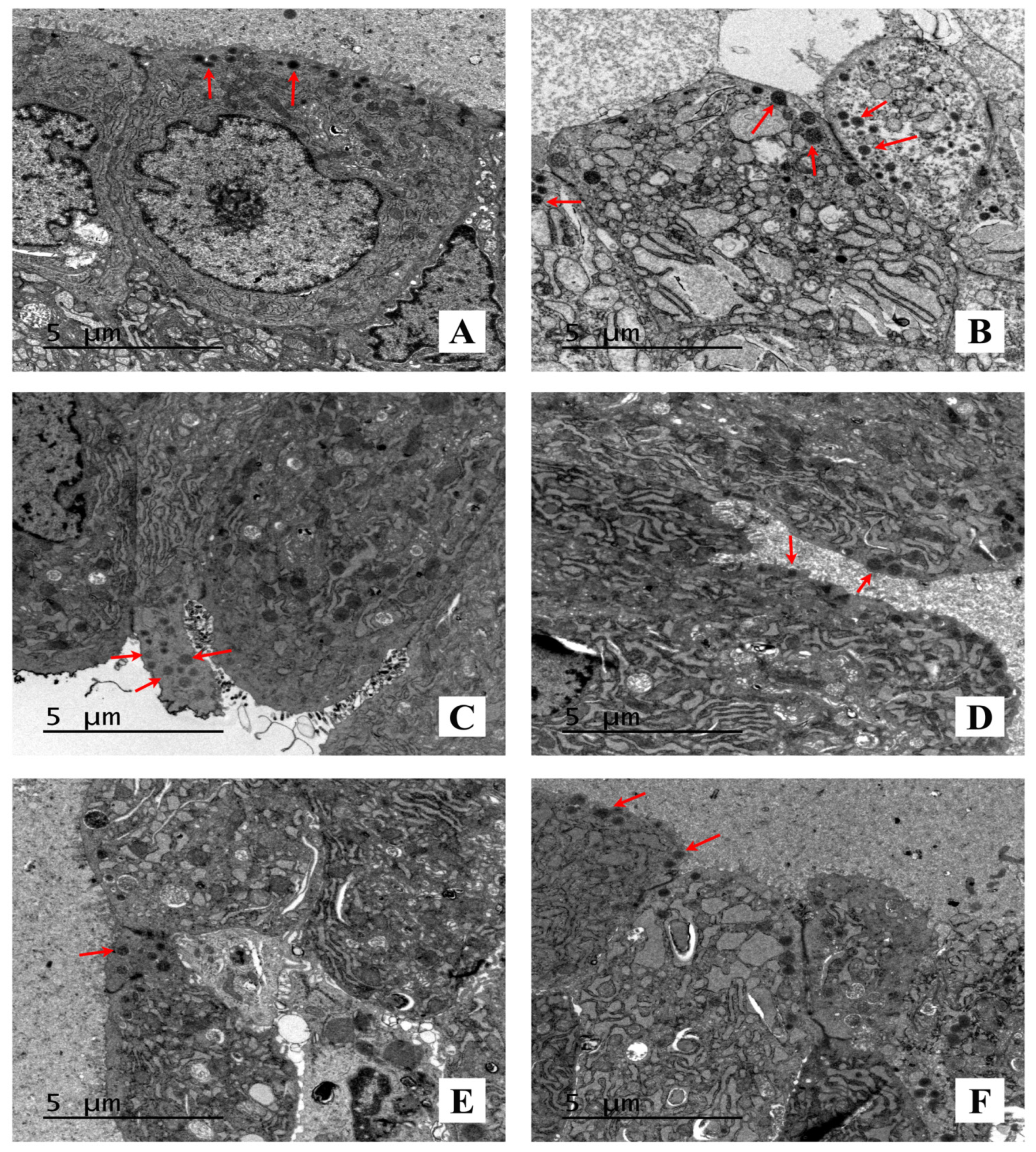
2.3. Effect of AST on the Histopathology and Ultrastructural Pathology of Prostate Tissues
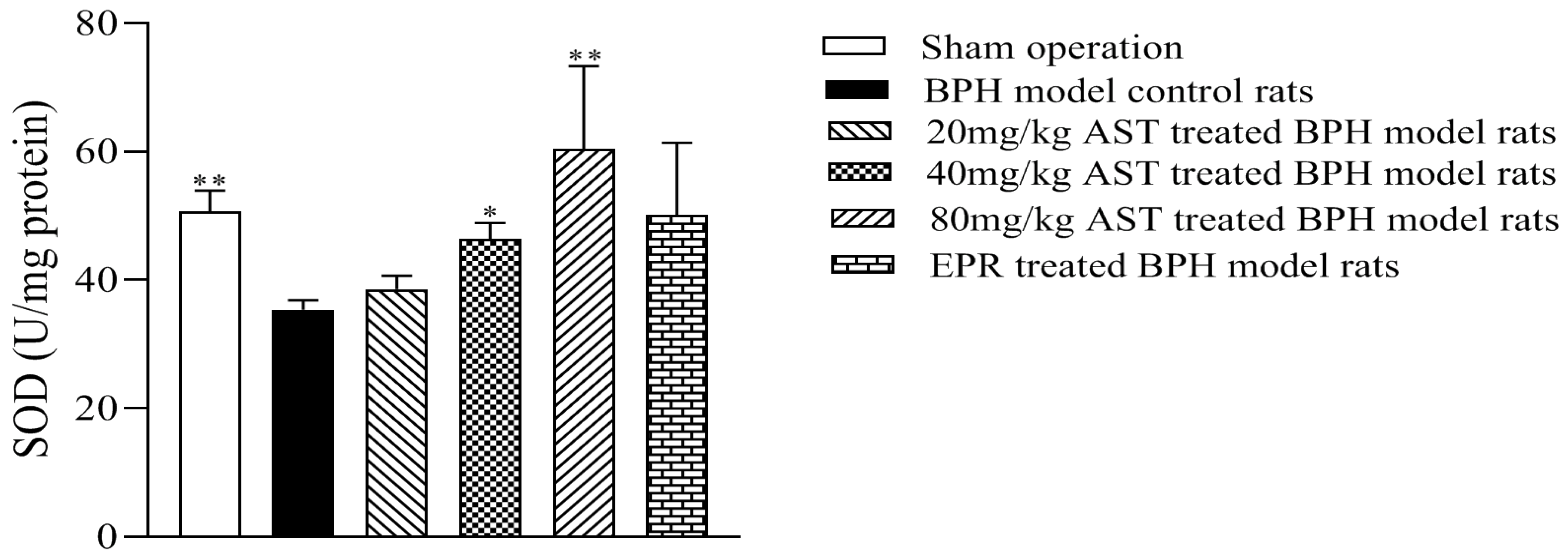
2.4. Effect of AST on the Superoxide Dismutase (SOD) Activity of Prostates
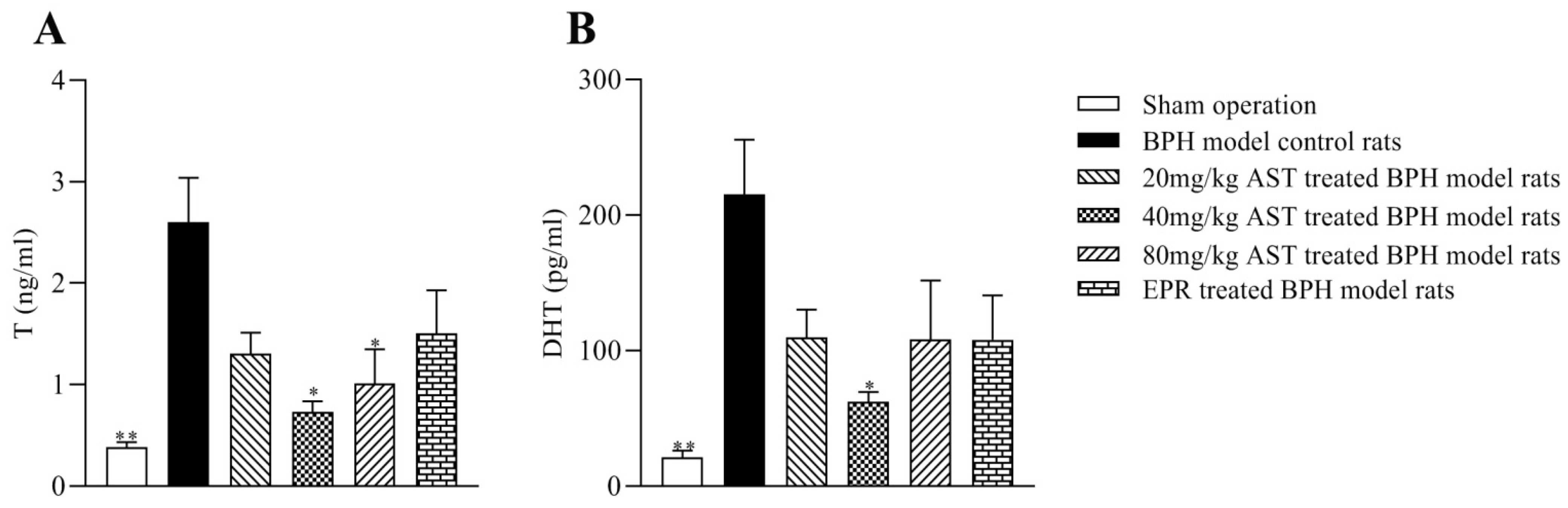
2.5. Effect of AST on the Levels of T and DHT
3. Discussion
4. Materials and Methods
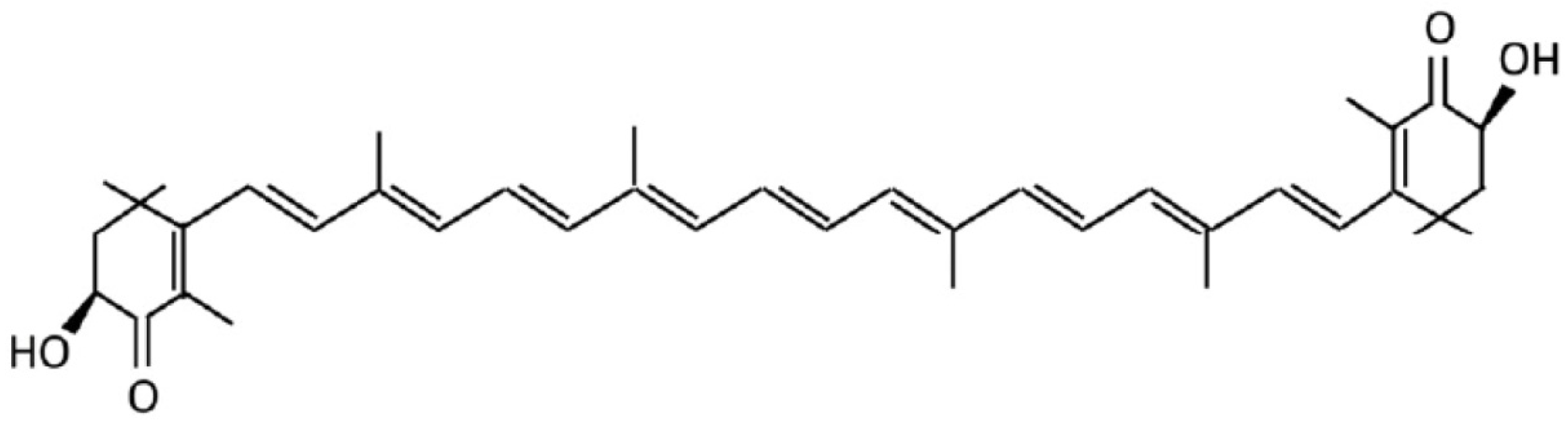
4.1. Materials
4.2. Animals
4.3. Study Design and Treatment
4.4. Prostate Index
4.5. Histopathological Examination
4.6. TEM
4.7. Analysis of SOD Activity
4.8. Analysis of Levels of T and DHT
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chughtai, B.; Forde, J.C.; Thomas, D.D.; Laor, L.; Hossack, T.; Woo, H.H.; Te, A.E.; Kaplan, S.A. Benign prostatic hyperplasia. Nat. Rev. Dis. Primers 2016, 2, 16031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, W.J.; Park, H.J.; Koo, H.C.; Kim, D.R.; Ha, U.S.; Kim, K.S.; Kim, S.J.; Cho, H.J.; Hong, S.H.; Lee, J.Y.; et al. The effect of seoritae extract in men with mild to moderate lower urinary tract symptoms suggestive of benign prostatic hyperplasia. Evid. Based Compl. Alt. 2016, 2016, 1960926. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Li, Y.; Liu, X.; Wu, J.; Zhou, L. Prostate Pharmacology; Shanghai Scientific and Technical Publishers: Shanghai, China, 2013; pp. 191–206. (In Chinese) [Google Scholar]
- Thomas, D.; Chughtai, B.; Kini, M.; Te, A. Emerging drugs for the treatment of benign prostatic hyperplasia. Expert Opin. Emerg. Dr. 2017, 22, 201–212. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef] [PubMed]
- McCall, B.; McPartland, C.K.; Moore, R.; Frank-Kamenetskii, A.; Booth, B.W. Effects of astaxanthin on the proliferation and migration of breast cancer cells in vitro. Antioxidants 2018, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.X.; Xiong, F. Astaxanthin and its Effects in inflammatory responses and inflammation-associated diseases: Recent advances and future directions. Molecules 2020, 25, 5342. [Google Scholar] [CrossRef]
- Kumar, A.; Dhaliwal, N.; Dhaliwal, J.; Dharavath, R.N.; Chopra, K. Astaxanthin attenuates oxidative stress and inflammatory responses in complete Freund-adjuvant-induced arthritis in rats. Pharmacol. Rep. 2020, 72, 104–114. [Google Scholar] [CrossRef]
- Gao, F.; Wu, X.; Mao, X.; Niu, F.; Zhang, B.; Dong, J.; Liu, B. Astaxanthin provides neuroprotection in an experimental model of traumatic brain injury via the Nrf2/HO-1 pathway. Am. J. Transl. Res. 2021, 13, 1483–1493. [Google Scholar]
- Ni, X.; Yu, H.; Wang, S.; Zhang, C.; Shen, S. Astaxanthin inhibits PC-3 xenograft prostate tumor growth in nude mice. Mar. Drugs 2017, 15, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buesen, R.; Schulte, S.; Strauss, V.; Treumann, S.; Becker, M.; Groters, S.; Carvalho, S.; van Ravenzwaay, B. Safety assessment of [3S, 3’S]-astaxanthin--Subchronic toxicity study in rats. Food Chem. Toxicol. 2015, 81, 129–136. [Google Scholar] [CrossRef]
- Creasy, D.; Bube, A.; de Rijk, E.; Kandori, H.; Kuwahara, M.; Masson, R.; Nolte, T.; Reams, R.; Regan, K.; Rehm, S.; et al. Proliferative and nonproliferative lesions of the rat and mouse male reproductive system. Toxicol. Pathol. 2012, 40 (Suppl. S6), 40s–121s. [Google Scholar] [CrossRef] [Green Version]
- Ginja, M.; Pires, M.J.; Gonzalo-Orden, J.M.; Seixas, F.; Correia-Cardoso, M. Anatomy and imaging of rat prostate: Practical monitoring in experimental cancer-induced protocols. Diagnostics 2019, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Madersbacher, S.; Sampson, N.; Culig, Z. Pathophysiology of benign prostatic hyperplasia and benign prostatic enlargement: A mini-review. Gerontology 2019, 65, 458–464. [Google Scholar] [CrossRef]
- Anderson, M.L. A preliminary investigation of the enzymatic inhibition of 5α-reductase and growth of prostatic carcinoma cell line LNCap-FGC by natural astaxanthin and saw palmetto lipid extract in vitro. J. Herb. Pharmacother. 2009, 5, 17–26. [Google Scholar] [CrossRef]
- Sun, S.Q.; Zhao, Y.X.; Li, S.Y.; Qiang, J.W.; Ji, Y.Z. Anti-tumor effects of astaxanthin by inhibition of the expression of STAT3 in prostate cancer. Mar. Drugs 2020, 18, 415. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fang, T.; Li, M.; Song, Y.; Li, J.; Xue, Z.; Li, J.; Bu, D.; Liu, W.; Zeng, Q.; et al. Pao pereira extract attenuates testosterone-induced benign prostatic hyperplasia in rats by inhibiting 5alpha-reductase. Sci. Rep. 2019, 9, 19703. [Google Scholar] [CrossRef]
- Wang, C.T.; Wang, Y.Y.; Liu, W.S.; Cheng, C.M.; Chiu, K.H.; Liu, L.L.; Liu, X.Z. Rhodobacter sphaeroides extract lycogen™ attenuates testosterone-induced benign prostate hyperplasia in Rats. Int. J. Mol. Sci. 2018, 19, 1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W. The ultrastructure of the rat ventral prostate. Acta Acad. Med. Qingdao 1986, 22, 22–28. (In Chinese) [Google Scholar]
- He, D.; Mi, J.; Wang, X.; Zheng, L.; Shi, Z. The histological and ultrastructural observation of experimental prostatic hyperplasia dogs. Chin. J. Vet. Med. 2009, 45, 52–53. (In Chinese) [Google Scholar]
- Wang, Y.R.; Xu, Y.; Jiang, Z.Z.; Zhang, L.Y.; Wang, T. Triptolide reduces prostate size and androgen level on testosterone-induced benign prostatic hyperplasia in Sprague Dawley rats. Chin. J. Nat. Med. 2017, 15, 341–346. [Google Scholar] [CrossRef]
- Minciullo, P.L.; Inferrera, A.; Navarra, M.; Calapai, G.; Magno, C.; Gangemi, S. Oxidative stress in benign prostatic hyperplasia: A systematic review. Urol. Int. 2015, 94, 249–254. [Google Scholar] [CrossRef]
- Zabaiou, N.; Mabed, D.; Lobaccaro, J.M.; Lahouel, M. Oxidative stress in benign prostate hyperplasia. Andrologia 2016, 48, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Roumeguere, T.; Sfeir, J.; El Rassy, E.; Albisinni, S.; Van Antwerpen, P.; Boudjeltia, K.Z.; Fares, N.; Kattan, J.; Aoun, F. Oxidative stress and prostatic diseases. Mol. Clin. Oncol. 2017, 7, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Ismy, J.; Sugandi, S.; Rachmadi, D.; Hardjowijoto, S.; Mustafa, A. The effect of exogenous superoxide dismutase (SOD) on caspase-3 activation and apoptosis induction in Pc-3 prostate cancer cells. Res. Rep. Urol. 2020, 12, 503–508. [Google Scholar] [CrossRef]
- Aydin, A.; Arsova-Sarafinovska, Z.; Sayal, A.; Eken, A.; Erdem, O.; Erten, K.; Ozgok, Y.; Dimovski, A. Oxidative stress and antioxidant status in non-metastatic prostate cancer and benign prostatic hyperplasia. Clin. Biochem. 2006, 39, 176–179. [Google Scholar] [CrossRef] [PubMed]
- El-Sherbiny, M.; El-Shafey, M.; El-Din El-Agawy, M.S.; Mohamed, A.S.; Eisa, N.H.; Elsherbiny, N.M. Diacerein ameliorates testosterone-induced benign prostatic hyperplasia in rats: Effect on oxidative stress, inflammation and apoptosis. Int. Immunopharmacol. 2021, 100, 108082. [Google Scholar] [CrossRef] [PubMed]
- Shoieb, S.M.; Esmat, A.; Khalifa, A.E.; Abdel-Naim, A.B. Chrysin attenuates testosterone-induced benign prostate hyperplasia in rats. Food Chem. Toxicol. 2018, 111, 650–659. [Google Scholar] [CrossRef]
- Abo-Youssef, A.M.; Afify, H.; Azouz, A.A.; Abdel-Rahman, H.M.; Abdel-Naim, A.B.; Allam, S. Febuxostat attenuates testosterone-induced benign prostatic hyperplasia in rats via inhibiting JAK/STAT axis. Life Sci. 2020, 260, 118414. [Google Scholar] [CrossRef] [PubMed]
- Coppenolle, F.V.; Bourhis, X.L.; Carpentier, F.O.; Delaby, G.; Cousse, H.; Raynaud, J.-P.; Dupouy, J.-P.; Prevarskaya, N. Pharmacological effects of the lipidosterolic extract of serenoa repens (permixonT) on ratprostate hyperplasia induced by hyperprolactinemia: Comparison with finasteride. Prostate 2000, 43, 49–58. [Google Scholar] [CrossRef]
- Sun, Z.Y.; Wu, H.Y.; Wang, M.Y.; Tu, Z.H. The mechanism of epristeride against benign prostatic hyperplasia. Eur. J. Pharmacol. 1999, 371, 227–233. [Google Scholar] [CrossRef]
- Mao, G.X.; Zheng, L.D.; Cao, Y.B.; Chen, Z.M.; Lv, Y.D.; Wang, Y.Z.; Hu, X.L.; Wang, G.F.; Yan, J. Antiaging effect of pine pollen in human diploid fibroblasts and in a mouse model induced by D-galactose. Oxid. Med. Cell. Longev. 2012, 2012, 750963. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liu, Y.; Cui, J.; Liu, H.; Liu, Y.B.; Qiao, W.L.; Sun, H.; Yan, C.D. Oxidative stress induces gastric submucosal arteriolar dysfunction in the elderly. World J. Gastroenterol. 2013, 19, 9439–9446. [Google Scholar] [CrossRef]
- Du, J.; Yin, G.; Hu, Y.; Shi, S.; Jiang, J.; Song, X.; Zhang, Z.; Wei, Z.; Tang, C.; Lyu, H. Coicis semen protects against focal cerebral ischemia-reperfusion injury by inhibiting oxidative stress and promoting angiogenesis via the TGFβ/ALK1/Smad1/5 signaling pathway. Aging 2020, 13, 877–893. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A. Changes in nitric oxide, carbon monoxide, hydrogen sulfide and male reproductive hormones in response to chronic restraint stress in rats. Free Radic. Biol. Med. 2021, 162, 353–366. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, W.; Fan, S.; Zhuang, H.; Han, Y.; Zhang, H.; Yuan, Z.; Weng, Q. Seasonal expressions of androgen receptor, estrogen receptors, 5α-reductases and P450arom in the epididymis of the male muskrat (Ondatra zibethicus). J. Steroid Biochem. Mol. Biol. 2019, 194, 105433. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.S.; Kolasa-Wołosiuk, A.; Pilutin, A. Finasteride-induced inhibition of 5α-reductase type 2 could lead to kidney damage-animal, experimental study. Int. J. Environ. Res. Public Health 2019, 16, 1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Hou, Y.; Wang, R.; Pan, Q.; Li, D.; Yan, H.; Sun, Z. Inhibitory Effect of Astaxanthin on Testosterone-Induced Benign Prostatic Hyperplasia in Rats. Mar. Drugs 2021, 19, 652. https://doi.org/10.3390/md19120652
Wang L, Hou Y, Wang R, Pan Q, Li D, Yan H, Sun Z. Inhibitory Effect of Astaxanthin on Testosterone-Induced Benign Prostatic Hyperplasia in Rats. Marine Drugs. 2021; 19(12):652. https://doi.org/10.3390/md19120652
Chicago/Turabian StyleWang, Liping, Yiwen Hou, Rong Wang, Qi Pan, Debao Li, Han Yan, and Zuyue Sun. 2021. "Inhibitory Effect of Astaxanthin on Testosterone-Induced Benign Prostatic Hyperplasia in Rats" Marine Drugs 19, no. 12: 652. https://doi.org/10.3390/md19120652