
The Critical Studies of Fucoxanthin Research Trends from 1928 to June 2021: A Bibliometric Review

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Methods
2.1. Data Mining
2.2. Data Refinement
2.3. Data Analysis
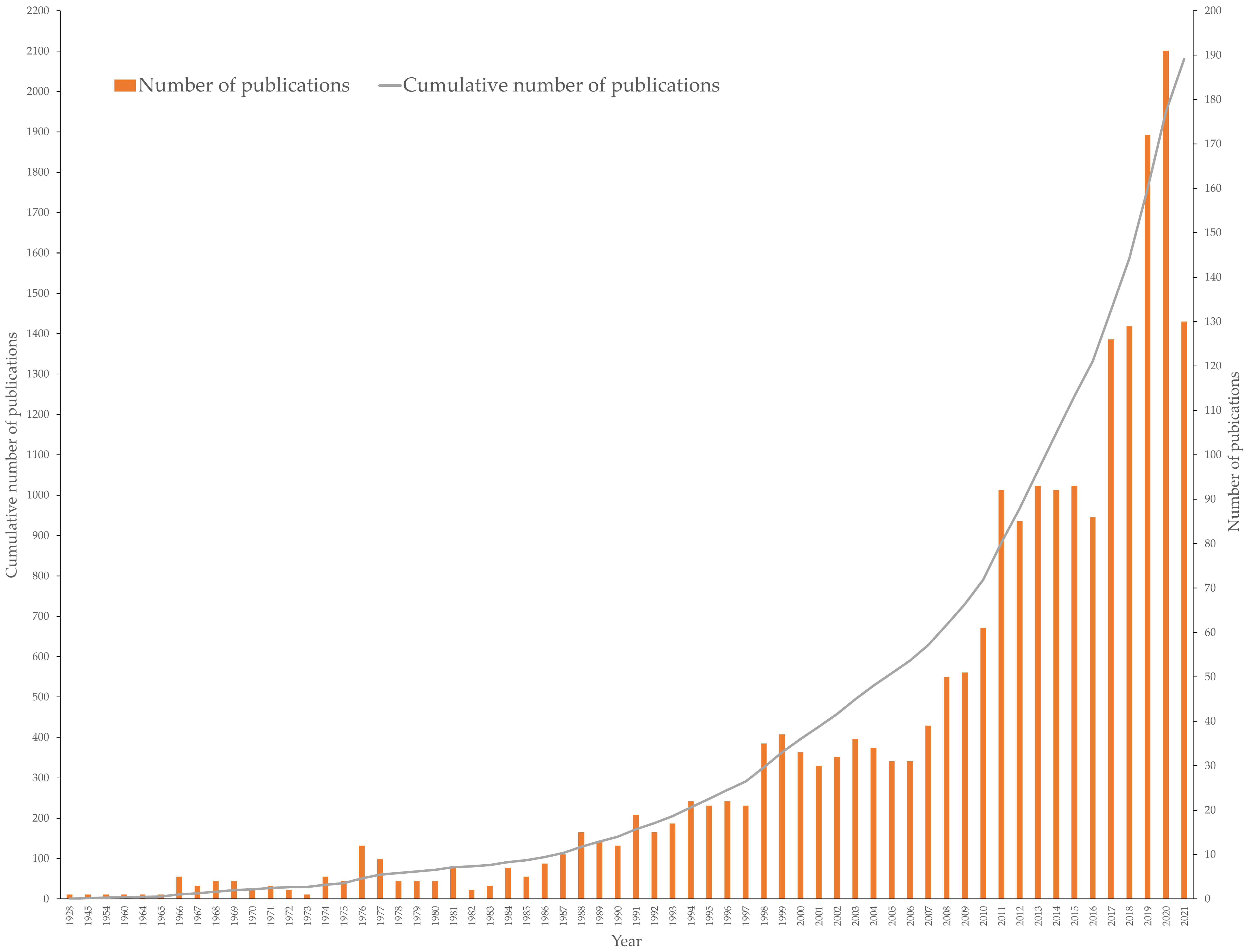
3. Publication Output
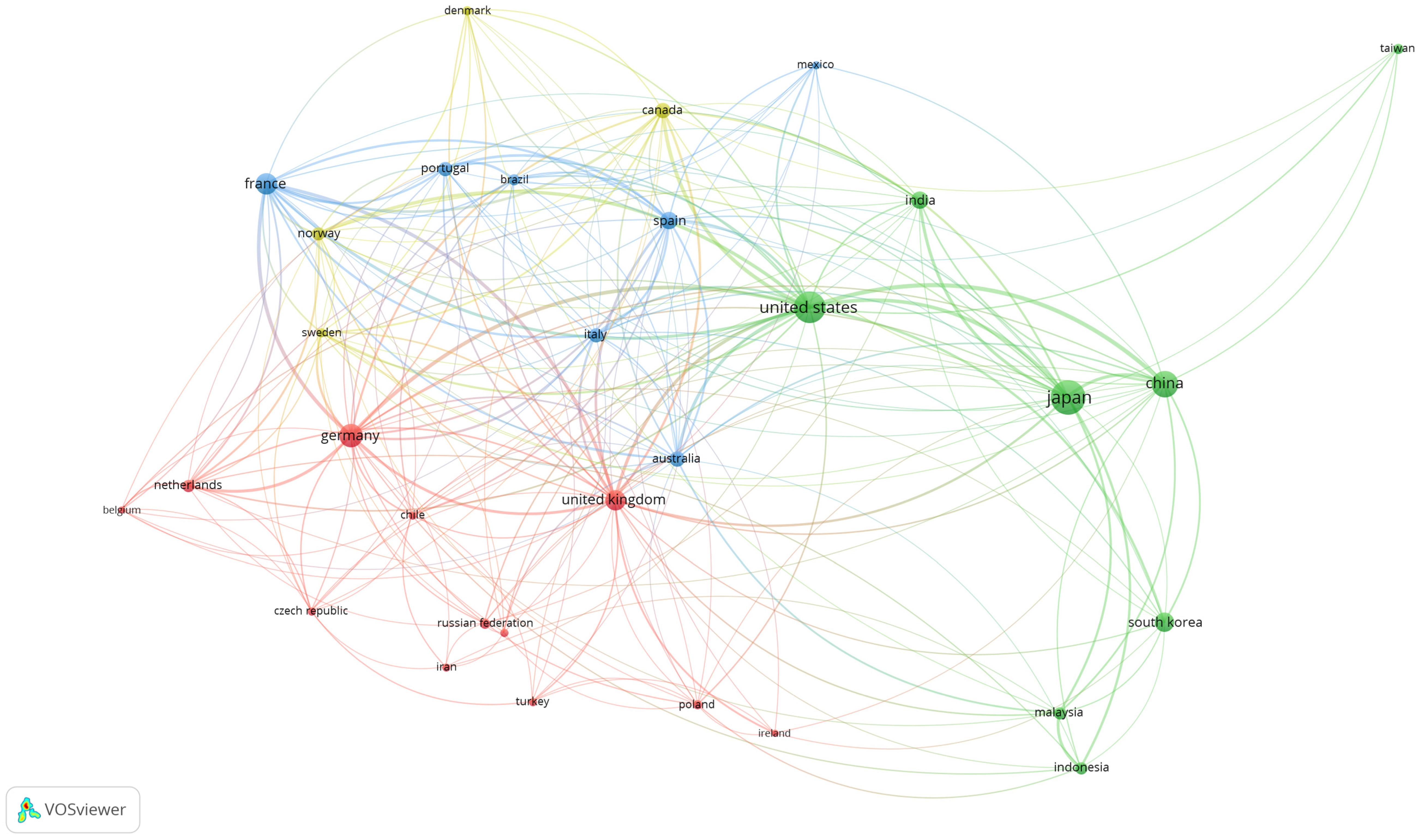
4. Global Production of Fucoxanthin Publications
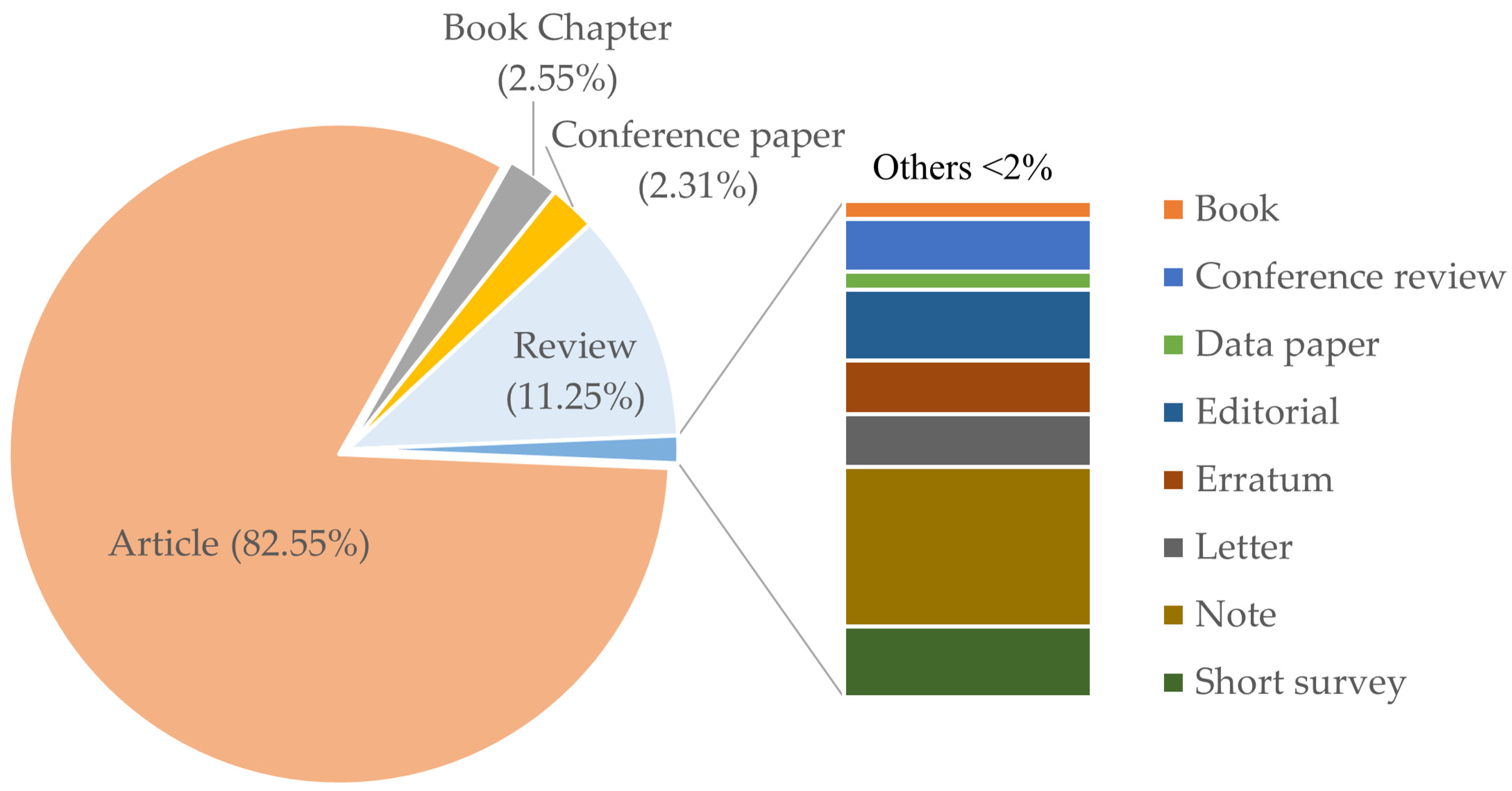
5. Subject Category, Sources and Citations
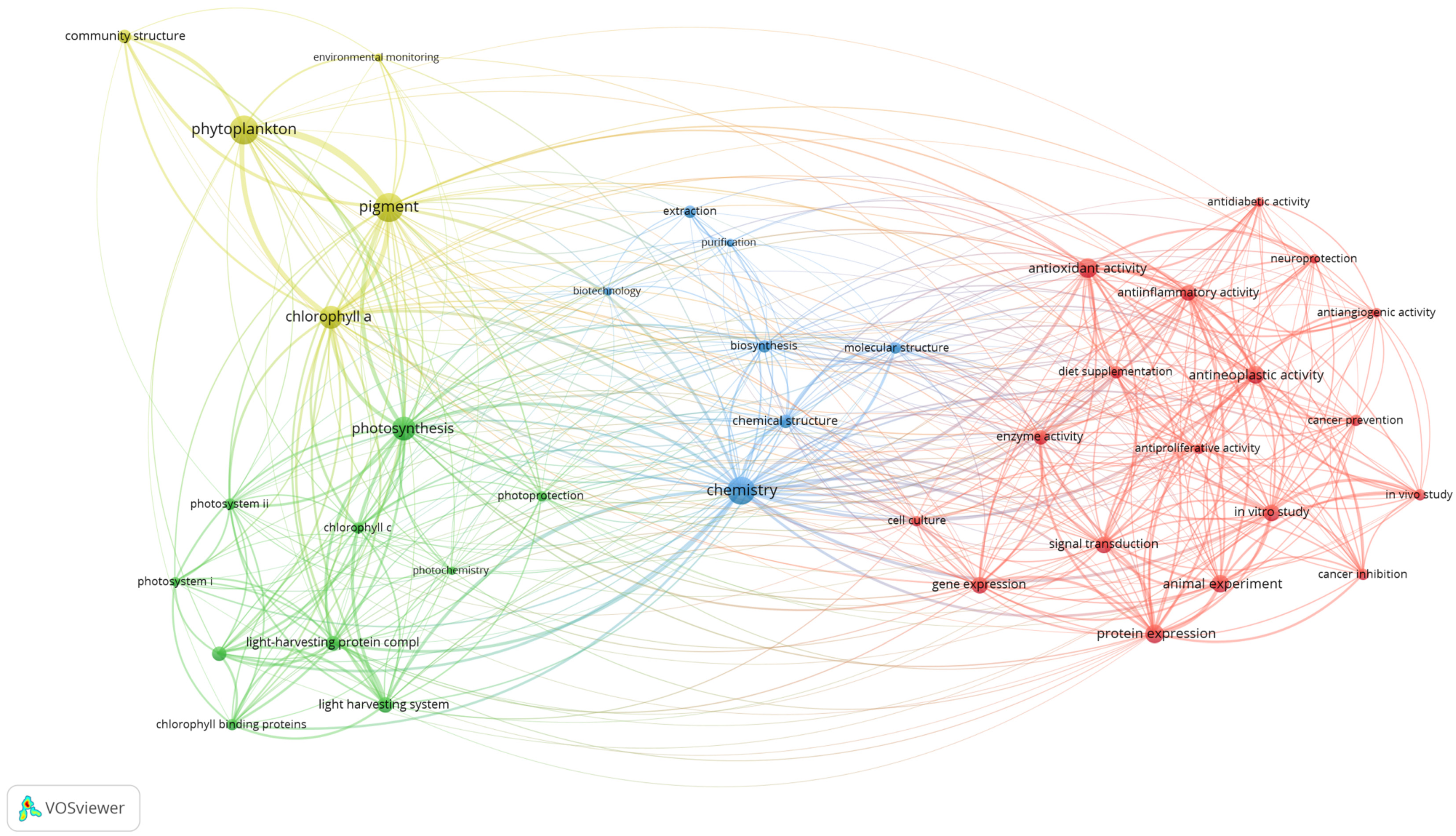
6. Research Concepts
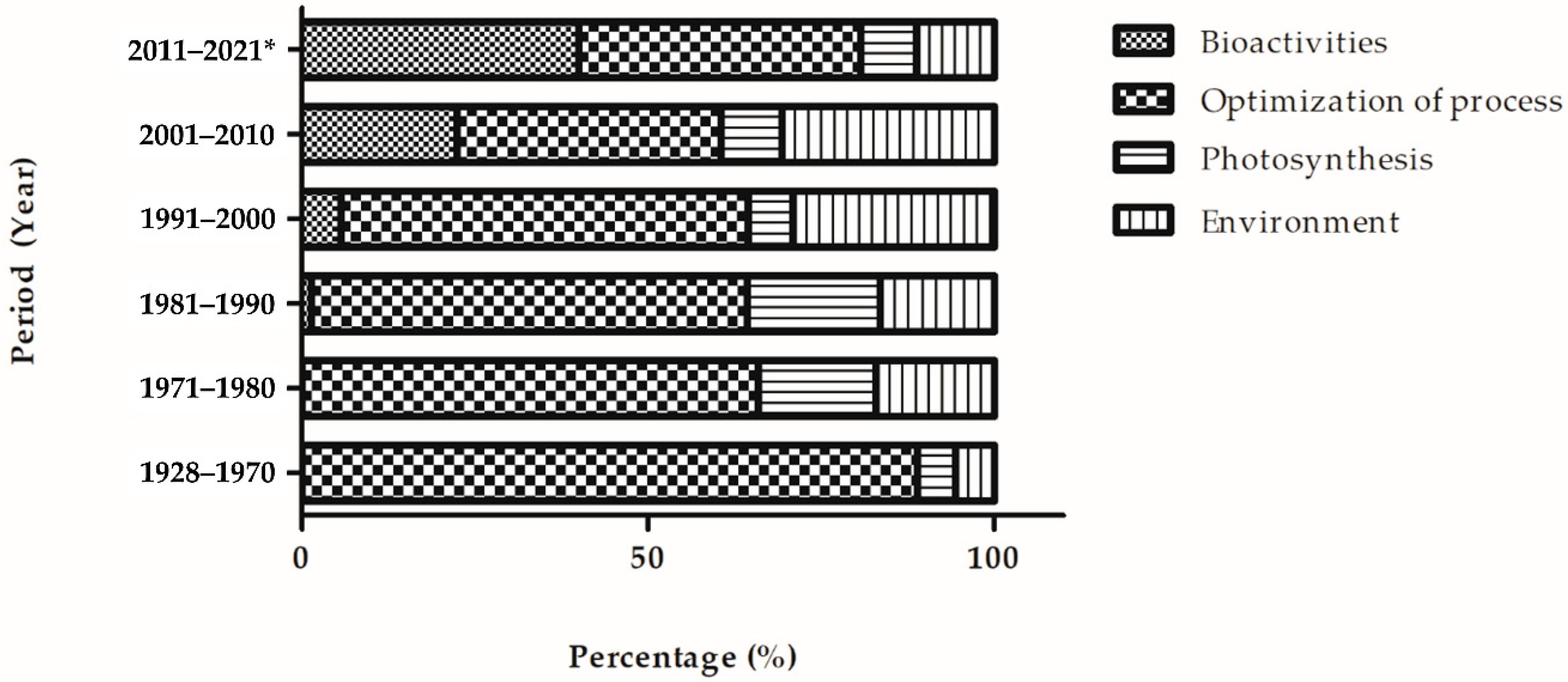
6.1. Research Trends of Fucoxanthin Research from 1928 to June 2021
6.1.1. Bioactivities of Fucoxanthin
6.1.2. Photosynthesis
6.1.3. Optimization of Process
6.1.4. Environment
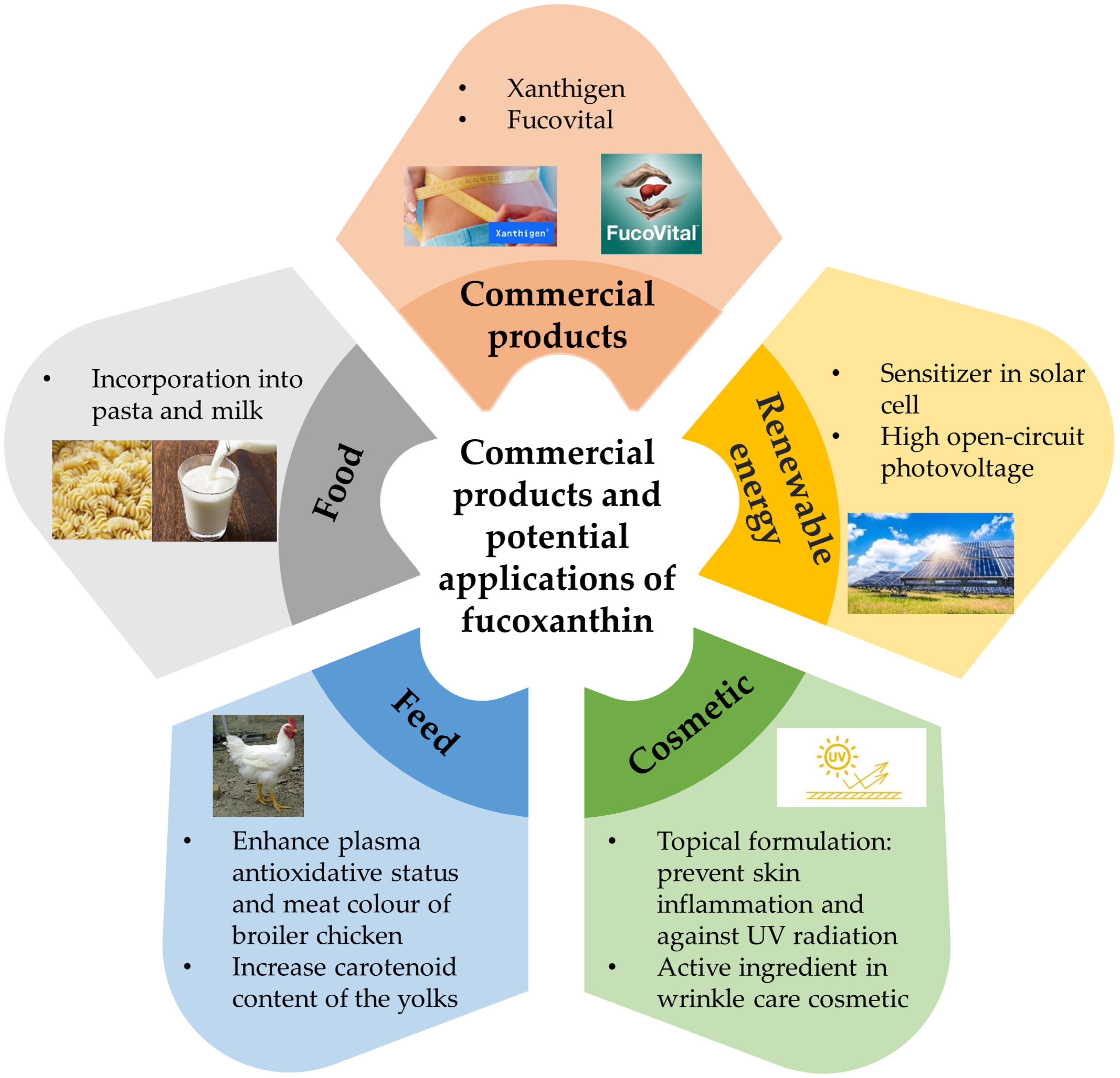
7. Commercial Products and Potential Applications of Fucoxanthin
8. Challenges and Possible Solutions
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liaaen-Jensen, S. Marine Carotenoids. In Chemical and Biochemical Perspective; Academic Press: New York, NY, USA, 1978; pp. 1–73. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef] [PubMed]
- Veith, T.; Brauns, J.; Weisheit, W.; Mittag, M.; Büchel, C. Identification of a specific fucoxanthin-chlorophyll protein in the light harvesting complex of photosystem I in the diatom Cyclotella meneghiniana. Biochim. Biophys. Acta Bioenerg. 2009, 1787, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarasaiyar, K.; Goh, B.-H.; Jeon, Y.-J.; Yow, Y.-Y. Algae Metabolites in Cosmeceutical: An Overview of Current Applications and Challenges. Mar. Drugs 2020, 18, 323. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary Combination of Fucoxanthin and Fish Oil Attenuates the Weight Gain of White Adipose Tissue and Decreases Blood Glucose in Obese/Diabetic KK-AyMice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef] [PubMed]
- Hitoe, S.; Shimoda, H. Seaweed Fucoxanthin Supplementation Improves Obesity Parameters in Mild Obese Japanese Subjects. Funct. Foods Heal. Dis. 2017, 7, 246. [Google Scholar] [CrossRef]
- Muthuirulappan, S.; Francis, S.P. Anti-Cancer Mechanism and Possibility of Nano-Suspension Formulation for a Marine Algae Product Fucoxanthin. Asian Pac. J. Cancer Prev. 2013, 14, 2213–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, M.-N.; Jeon, S.-M.; Kim, H.-J.; Lee, M.-K.; Shin, S.-K.; Shin, Y.C.; Park, Y.-B.; Choi, M.-S. Fucoxanthin supplementation improves plasma and hepatic lipid metabolism and blood glucose concentration in high-fat fed C57BL/6N mice. Chem. Interact. 2010, 186, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.Y.B.; Oliveira, C.D.L.; Müller, M.N.; Santos, E.P.; Dantas, D.M.M.; Gálvez, A.O. A Scientometric Overview of Global Dinoflagellate Research. Publications 2020, 8, 50. [Google Scholar] [CrossRef]
- Garrido-Cardenas, J.A.; Manzano-Agugliaro, F.; Acien-Fernandez, F.G.; Molina-Grima, E. Microalgae research worldwide. Algal Res. 2018, 35, 50–60. [Google Scholar] [CrossRef]
- Silva, S.C.; Ferreira, I.C.F.R.; Dias, M.M.; Barreiro, M.F. Microalgae-Derived Pigments: A 10-Year Bibliometric Review and Industry and Market Trend Analysis. Molecules 2020, 25, 3406. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhu, L. The scientometric analysis of the research on microalgae-based wastewater treatment. Environ. Sci. Pollut. Res. 2021, 28, 25339–25348. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, N.J.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2009, 84, 523–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-tool for comprehensive science mapping analysis. J. Inf. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Tang, Y.; Xin, H.; Yang, F.; Long, X. A historical review and bibliometric analysis of nanoparticles toxicity on algae. J. Nanoparticle Res. 2018, 20, 92. [Google Scholar] [CrossRef]
- Terasaki, M.; Inoue, T.; Murase, W.; Kubota, A.; Kojima, H.; Kojoma, M.; Ohta, T.; Maeda, H.; Miyashita, K.; Mutoh, M.; et al. Fucoxanthinol Induces Apoptosis in a Pancreatic Intraepithelial Neoplasia Cell Line. Cancer Genom. Proteom. 2021, 18, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Hamoya, T.; Kubota, A.; Kojima, H.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M. Fucoxanthin Prevents Colorectal Cancer Development in Dextran Sodium Sulfate-treated ApcMin/+ Mice. Anticancer. Res. 2021, 41, 1299–1305. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Hosokawa, M.; Miyashita, K.; Nishino, H.; Hashimoto, T. Effects of Fucoxanthin on the Inhibition of Dexamethasone-Induced Skeletal Muscle Loss in Mice. Nutrients 2021, 13, 1079. [Google Scholar] [CrossRef] [PubMed]
- Agostini, A.; Büchel, C.; Di Valentin, M.; Carbonera, D. A distinctive pathway for triplet-triplet energy transfer photoprotection in fucoxanthin chlorophyll-binding proteins from Cyclotella meneghiniana. Biochim. Biophys. Acta Bioenerg. 2020, 1862, 148310. [Google Scholar] [CrossRef] [PubMed]
- Gelzinis, A.; Augulis, R.; Büchel, C.; Robert, B.; Valkunas, L. Confronting FCP structure with ultrafast spectroscopy data: Evidence for structural variations. Phys. Chem. Chem. Phys. 2020, 23, 806–821. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Gao, G.; Hand, E.S.; Kispert, L.D.; Strand, A.; Liaaen-Jensen, S. Iodine-Catalyzed R/S Isomerization of Allenic Carotenoids. J. Phys. Chem. A 2002, 106, 2520–2525. [Google Scholar] [CrossRef]
- Refvem, T.; Strand, A.; Kjeldstad, B.; Haugan, J.A.; Liaaen-Jensen, S. Stereoisomerization of Allenic Carotenoids-Kinetic, Thermodynamic and Mechanistic Aspects. Acta Chem. Scand. 1999, 53, 114–123. [Google Scholar] [CrossRef]
- Nagao, R.; Kato, K.; Ifuku, K.; Suzuki, T.; Kumazawa, M.; Uchiyama, I.; Kashino, Y.; Dohmae, N.; Akimoto, S.; Shen, J.-R.; et al. Structural basis for assembly and function of a diatom photosystem I-light-harvesting supercomplex. Nat. Commun. 2020, 11, 2481. [Google Scholar] [CrossRef] [PubMed]
- Oka, K.; Ueno, Y.; Yokono, M.; Shen, J.-R.; Nagao, R.; Akimoto, S. Adaptation of light-harvesting and energy-transfer processes of a diatom Phaeodactylum tricornutum to different light qualities. Photosynth. Res. 2020, 146, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, S.; Ueno, Y.; Yokono, M.; Shen, J.-R.; Nagao, R. Adaptation of light-harvesting and energy-transfer processes of a diatom Chaetoceros gracilis to different light qualities. Photosynth. Res. 2020, 146, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Fagan, J.; Eddens, K.S.; Dolly, J.; Vanderford, N.L.; Weiss, H.; Levens, J.S. Assessing Research Collaboration through Co-authorship Network Analysis. J. Res. Adm. 2018, 49, 76–99. [Google Scholar] [PubMed]
- Shawky, S.M.; Fathalla, S.I.; Orabi, S.H.; El-Mosalhi, H.H.; Abu-Alya, I.S. Effect of Amphora coffeaeformis and Star Anise as Dietary Supplementson the Immunity and Growth Performance of Broiler Chickens. J. World’s Poult. Res. 2020, 10, 631–642. [Google Scholar] [CrossRef]
- Hao, T.-B.; Yang, Y.-F.; Balamurugan, S.; Li, D.-W.; Yang, W.-D.; Li, H.-Y. Enrichment of f/2 medium hyperaccumulates biomass and bioactive compounds in the diatom Phaeodactylum tricornutum. Algal Res. 2020, 47, 101872. [Google Scholar] [CrossRef]
- Conceição, D.; Lopes, R.G.; Derner, R.; Cella, H.; Carmo, A.P.B.D.; D’Oca, M.G.M.; Petersen, R.; Passos, M.F.; Vargas, J.V.C.; Galli-Terasawa, L.V.; et al. The effect of light intensity on the production and accumulation of pigments and fatty acids in Phaeodactylum tricornutum. Environ. Boil. Fishes 2020, 32, 1017–1025. [Google Scholar] [CrossRef]
- Tanabe, M.; Ueno, Y.; Yokono, M.; Shen, J.-R.; Nagao, R.; Akimoto, S. Changes in excitation relaxation of diatoms in response to fluctuating light, probed by fluorescence spectroscopies. Photosynth. Res. 2020, 146, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Ifuku, K.; Yan, D.; Miyahara, M.; Inoue-Kashino, N.; Yamamoto, Y.Y.; Kashino, Y. A stable and efficient nuclear transformation system for the diatom Chaetoceros gracilis. Photosynth. Res. 2014, 123, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Manfellotto, F.; Stella, G.R.; Falciatore, A.; Brunet, C.; Ferrante, M.I. Engineering the Unicellular Alga Phaeodactylum tricornutum for Enhancing Carotenoid Production. Antioxidants 2020, 9, 757. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Leyva, A.B.; Guo, J.; Mudd, E.A.; Wong, J.; Schwartz, J.-M.; Day, A. The chloroplast genome of the marine microalga Tisochrysis lutea. Mitochondrial DNA Part B 2019, 4, 253–255. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Jung, G.; Hwang, Y.-S.; Jin, E. Dynamic response of the transcriptome of a psychrophilic diatom, Chaetoceros neogracile, to high irradiance. Planta 2009, 231, 349–360. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Imam, M.U.; Foo, J.B.; Ismail, N.; Azmi, N.H.; Tor, Y.S.; Khong, N.M.; Ismail, M. Increased fucoxanthin in Chaetoceros calcitrans extract exacerbates apoptosis in liver cancer cells via multiple targeted cellular pathways. Biotechnol. Rep. 2018, 21, e00296. [Google Scholar] [CrossRef]
- Manmuan, S.; Manmuan, P. Fucoxanthin Enhances 5-FU Chemotherapeutic Efficacy in Colorectal Cancer Cells by Affecting MMP-9 Invasive Proteins. J. Appl. Pharm. Sci. 2019, 9, 7–14. [Google Scholar]
- Smith, D.R. Citation Indexing and Highly Cited Articles in the Australian Veterinary Journal. Aust. Vet. J. 2008, 86, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chuda, Y.; Suzuki, M.; Nagata, T. Fucoxanthin as the Major Antioxidant in Hijikia fusiformis, a Common Edible Seaweed. Biosci. Biotechnol. Biochem. 1999, 63, 605–607. [Google Scholar] [CrossRef]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical Scavenging and Singlet Oxygen Quenching Activity of Marine Carotenoid Fucoxanthin and Its Metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Kudo, M.; Maeda, H.; Kohno, H.; Tanaka, T.; Miyashita, K. Fucoxanthin Induces Apoptosis and Enhances the Antiproliferative Effect of the PPARγ Ligand, Troglitazone, on Colon Cancer Cells. Biochim. Biophys. Acta General Subj. 2004, 1675, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Heo, S.J.; Yoon, W.J.; Kang, S.M.; Ahn, G.; Yi, T.H.; Jeon, Y.J. Fucoxanthin Inhibits the Inflammatory Response by Suppressing the Activation of NF-ΚB and MAPKs in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.-J.; Yoon, W.-J.; Kim, K.-N.; Ahn, G.-N.; Kang, S.-M.; Kang, D.-H.; Affan, A.; Oh, C.; Jung, W.-K.; Jeon, Y.-J. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef]
- Jones, K.S.; Jackson, D.M. The use of automatically-obtained keyword classifications for information retrieval. Inf. Storage Retr. 1970, 5, 175–201. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; Reina, G.G.-B.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef]
- Okuzumi, J.; Nishino, H.; Murakoshi, M.; Iwashima, A.; Tanaka, Y.; Yamane, T.; Fujita, Y.; Takahashi, T. Inhibitory effects of fucoxanthin, a natural carotenoid, on N-myc expression and cell cycle progression in human malignant tumor cells. Cancer Lett. 1990, 55, 75–81. [Google Scholar] [CrossRef]
- Okuzumi, J.; Takahashi, T.; Yamane, T.; Kitao, Y.; Inagake, M.; Ohya, K.; Nishino, H.; Tanaka, Y. Inhibitory Effects of Fucoxanthin, a Natural Carotenoid, on N-Ethyl-N′-Nitro-N-Nitrosoguanidine-Induced Mouse Duodenal Carcinogenesis. Cancer Lett. 1993, 68, 159–168. [Google Scholar] [CrossRef]
- Greenwald, P.; Kelloff, G.; Burch-Whitman, C.; Kramer, B.S. Chemoprevention. CA Cancer J. Clin. 1995, 45, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-J.; Pezzuto, J.M. Antioxidant Marine Products in Cancer Chemoprevention. Antioxid. Redox Signal. 2013, 19, 115–138. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Bharathiraja, S.; Moorthy, M.S.; Mondal, S.; Seo, H.; Lee, K.D.; Oh, J. Marine natural pigments as potential sources for therapeutic applications. Crit. Rev. Biotechnol. 2017, 38, 745–761. [Google Scholar] [CrossRef] [PubMed]
- Ojulari, O.V.; Lee, S.G.; Nam, J.-O. Therapeutic Effect of Seaweed Derived Xanthophyl Carotenoid on Obesity Management; Overview of the Last Decade. Int. J. Mol. Sci. 2020, 21, 2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathasivam, R.; Ki, J.-S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Balasubramanian, M.P.; Nishigaki, I. Cancer Preventive Efficacy of Marine Carotenoid Fucoxanthin: Cell Cycle Arrest and Apoptosis. Nutrients 2013, 5, 4978–4989. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J. Fucoxanthin and Its Metabolite Fucoxanthinol in Cancer Prevention and Treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaulmann, A.; Bohn, T. Carotenoids, Inflammation, and Oxidative Stress—Implications of Cellular Signaling Pathways and Relation to Chronic Disease Prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef] [PubMed]
- Muradian, K.; Vaiserman, A.; Min, K.-J.; Fraifeld, V.E. Fucoxanthin and lipid metabolism: A minireview. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 891–897. [Google Scholar] [CrossRef]
- Hu, X.; Li, Y.; Li, C.; Fu, Y.; Cai, F.; Chen, Q.; Li, D. Combination of fucoxanthin and conjugated linoleic acid attenuates body weight gain and improves lipid metabolism in high-fat diet-induced obese rats. Arch. Biochem. Biophys. 2012, 519, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Molina, N.; Morandi, A.C.; Bolin, A.; Otton, R. Comparative effect of fucoxanthin and vitamin C on oxidative and functional parameters of human lymphocytes. Int. Immunopharmacol. 2014, 22, 41–50. [Google Scholar] [CrossRef]
- Lin, H.-T.V.; Tsou, Y.-C.; Chen, Y.-T.; Lu, W.-J.; Hwang, P.-A. Effects of Low-Molecular-Weight Fucoidan and High Stability Fucoxanthin on Glucose Homeostasis, Lipid Metabolism, and Liver Function in a Mouse Model of Type II Diabetes. Mar. Drugs 2017, 15, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.; Li, K.; Lin, Y. Fucoidan–Fucoxanthin Ameliorated Cardiac Function via IRS1GRB2 SOS1, GSK3βCREB Pathways and Metabolic Pathways in Senescent Mice. Mar. Drugs 2019, 17, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, Y.-T.; Chen, H.-Y.; Lin, P.-H.; Hsia, S.-M. Preventive Effects of Fucoidan and Fucoxanthin on Hyperuricemic Rats Induced by Potassium Oxonate. Mar. Drugs 2019, 17, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Luna, A.; Ávila-Román, J.; Oliveira, H.; Motilva, V.; Talero, E. Fucoxanthin and Rosmarinic Acid Combination Has Anti-Inflammatory Effects through Regulation of NLRP3 Inflammasome in UVB-Exposed HaCaT Keratinocytes. Mar. Drugs 2019, 17, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menichini, D.; Alrais, M.; Liu, C.; Xia, Y.; Blackwell, S.C.; Facchinetti, F.; Sibai, B.M.; Longo, M. Maternal Supplementation of Inositols, Fucoxanthin, and Hydroxytyrosol in Pregnant Murine Models of Hypertension. Am. J. Hypertens. 2020, 33, 652–659. [Google Scholar] [CrossRef]
- Kawakamia, T.; Tsushimab, M.; Katabamia, Y.; Mine, M.; Ishida, A.; Matsuno, M. Effect of b, b-Carotene, b-Echinenone, Astaxanthin, Fucoxanthin, Vitamin A and Vitamin E on the Biological Defense of the Sea Urchin Pseudocentrotus depressus. J. Exp. Mar. Biol. Ecol. 1998, 226, 165–174. [Google Scholar] [CrossRef]
- Kitamura, M.; Koyama, T.; Nakano, Y.; Uemura, D. Characterization of a natural inducer of coral larval metamorphosis. J. Exp. Mar. Biol. Ecol. 2007, 340, 96–102. [Google Scholar] [CrossRef]
- Lashmanova, E.; Proshkina, E.; Zhikrivetskaya, S.; Shevchenko, O.; Marusich, E.; Leonov, S.; Melerzanov, A.; Zhavoronkov, A.; Moskalev, A. Fucoxanthin increases lifespan of Drosophila melanogaster and Caenorhabditis elegans. Pharmacol. Res. 2015, 100, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Green, B.R.; Durnford, D.G. The chlorophyll-carotenoid proteins of oxygenic photosynthesis. Annu. Rev. Plant Biol. 1996, 47, 685–714. [Google Scholar] [CrossRef] [PubMed]
- Nagao, R.; Takahashi, S.; Suzuki, T.; Dohmae, N.; Nakazato, K.; Tomo, T. Comparison of oligomeric states and polypeptide compositions of fucoxanthin chlorophyll a/c-binding protein complexes among various diatom species. Photosynth. Res. 2013, 117, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Depauw, F.A.; Rogato, A.; Ribera d’Alcalá, M.; Falciatore, A. Exploring the Molecular Basis of Responses to Light in Marine Diatoms. J. Exp. Bot. 2012, 63, 1575–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.; Zhou, S.; Allen, A.; Apt, K.E.; Bechner, M.; et al. The Genome of the Diatom Thalassiosira pseudonana: Ecology, Evolution, and Metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowler, C.; Allen, A.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lepetit, B.; Volke, D.; Gilbert, M.; Wilhelm, C.; Goss, R. Evidence for the Existence of One Antenna-Associated, Lipid-Dissolved and Two Protein-Bound Pools of Diadinoxanthin Cycle Pigments in Diatoms. Plant Physiol. 2010, 154, 1905–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailleul, B.; Rogato, A.; de Martino, A.; Coesel, S.; Cardol, P.; Bowler, C.; Falciatore, A.; Finazzi, G. An atypical member of the light-harvesting complex stress-related protein family modulates diatom responses to light. Proc. Natl. Acad. Sci. USA 2010, 107, 18214–18219. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.-H.; Green, B.R. Photoprotection in the diatom Thalassiosira pseudonana: Role of LI818-like proteins in response to high light stress. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1449–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepetit, B.; Volke, D.; Szabo, M.; Hoffmann, R.; Garab, G.; Wilhelm, C.; Goss, R. Spectroscopic and Molecular Characterization of the Oligomeric Antenna of the Diatom Phaeodactylum tricornutum. Biochemistry 2007, 46, 9813–9822. [Google Scholar] [CrossRef] [PubMed]
- Gundermann, K.; Schmidt, M.; Weisheit, W.; Mittag, M.; Büchel, C. Identification of several sub-populations in the pool of light harvesting proteins in the pennate diatom Phaeodactylum tricornutum. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundermann, K.; Wagner, V.; Mittag, M.; Büchel, C. Fucoxanthin-Chlorophyll Protein Complexes of the Centric Diatom Cyclotella meneghiniana Differ in Lhcx1 and Lhcx6_1 Content. Plant Physiol. 2019, 179, 1779–1795. [Google Scholar] [CrossRef] [PubMed]
- Büchel, C. Fucoxanthin-Chlorophyll Proteins in Diatoms: 18 and 19 kDa Subunits Assemble into Different Oligomeric States. Biochemistry 2003, 42, 13027–13034. [Google Scholar] [CrossRef]
- Kansy, M.; Volke, D.; Sturm, L.; Wilhelm, C.; Hoffmann, R.; Goss, R. Pre-purification of diatom pigment protein complexes provides insight into the heterogeneity of FCP complexes. BMC Plant Biol. 2020, 20, 456. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, S.; Pi, X.; Kuang, T.; Sui, S.; Shen, J. Structural Features of the Diatom Photosystem II–light-harvesting Antenna Complex. FEBS J. 2020, 287, 2191–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Pi, X.; Huang, Y.; Han, G.; Chen, X.; Qin, X.; Huang, G.; Zhao, S.; Yang, Y.; Kuang, T.; et al. Structural basis for energy transfer in a huge diatom PSI-FCPI supercomplex. Nat. Commun. 2020, 11, 5081. [Google Scholar] [CrossRef]
- Nagao, R.; Kato, K.; Suzuki, T.; Ifuku, K.; Uchiyama, I.; Kashino, Y.; Dohmae, N.; Akimoto, S.; Shen, J.-R.; Miyazaki, N.; et al. Structural basis for energy harvesting and dissipation in a diatom PSII–FCPII supercomplex. Nat. Plants 2019, 5, 890–901. [Google Scholar] [CrossRef]
- Pi, X.; Zhao, S.; Wang, W.; Liu, D.; Xu, C.; Han, G.; Kuang, T.; Sui, S.-F.; Shen, J.-R. The pigment-protein network of a diatom photosystem II–light-harvesting antenna supercomplex. Science 2019, 365, eaax4406. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yu, L.-J.; Xu, C.; Tomizaki, T.; Zhao, S.; Umena, Y.; Chen, X.; Qin, X.; Xin, Y.; Suga, M.; et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science 2019, 363, eaav0365. [Google Scholar] [CrossRef]
- Papagiannakis, E.; Van Stokkum, I.H.M.; Fey, H.; Büchel, C.; Van Grondelle, R. Spectroscopic Characterization of the Excitation Energy Transfer in the Fucoxanthin–Chlorophyll Protein of Diatoms. Photosynth. Res. 2005, 86, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Zigmantas, D.; Hiller, R.G.; Sharples, F.P.; Frank, H.A.; Sundström, V.; Polívka, T. Effect of a Conjugated Carbonyl Group on the Photophysical Properties of Carotenoids. Phys. Chem. Chem. Phys. 2004, 6, 3009–3016. [Google Scholar] [CrossRef]
- Gildenhoff, N.; Herz, J.; Gundermann, K.; Büchel, C.; Wachtveitl, J. The excitation energy transfer in the trimeric fucoxanthin–chlorophyll protein from Cyclotella meneghiniana analyzed by polarized transient absorption spectroscopy. Chem. Phys. 2010, 373, 104–109. [Google Scholar] [CrossRef]
- Ramanan, C.; Berera, R.; Gundermann, K.; van Stokkum, I.; Büchel, C.; van Grondelle, R. Exploring the Mechanism(s) of Energy Dissipation in the Light Harvesting Complex of the Photosynthetic Algae Cyclotella meneghiniana. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 1507–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premvardhan, L.; Sandberg, D.J.; Fey, H.; Birge, R.R.; Büchel, C.; van Grondelle, R. The Charge-Transfer Properties of the S2 State of Fucoxanthin in Solution and in Fucoxanthin Chlorophyll-a/c2 Protein (FCP) Based on Stark Spectroscopy and Molecular-Orbital Theory. J. Phys. Chem. B 2008, 112, 11838–11853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, M.; Premvardhan, L.; Lepetit, B.; Goss, R.; Wilhelm, C.; Garab, G. Functional heterogeneity of the fucoxanthins and fucoxanthin-chlorophyll proteins in diatom cells revealed by their electrochromic response and fluorescence and linear dichroism spectra. Chem. Phys. 2010, 373, 110–114. [Google Scholar] [CrossRef] [Green Version]
- Premvardhan, L.; Bordes, L.; Beer, A.; Büchel, C.; Robert, B. Carotenoid Structures and Environments in Trimeric and Oligomeric Fucoxanthin Chlorophyll a/c2 Proteins from Resonance Raman Spectroscopy. J. Phys. Chem. B 2009, 113, 12565–12574. [Google Scholar] [CrossRef]
- Akimoto, S.; Teshigahara, A.; Yokono, M.; Mimuro, M.; Nagao, R.; Tomo, T. Excitation Relaxation Dynamics and Energy Transfer in Fucoxanthin–chlorophyll a/c-Protein Complexes, Probed by Time-Resolved Fluorescence. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 1514–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gildenhoff, N.; Amarie, S.; Gundermann, K.; Beer, A.; Büchel, C.; Wachtveitl, J. Oligomerization and pigmentation dependent excitation energy transfer in fucoxanthin–chlorophyll proteins. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Kosumi, D.; Kita, M.; Fujii, R.; Sugisaki, M.; Oka, N.; Takaesu, Y.; Taira, T.; Iha, M.; Hashimoto, H. Excitation Energy-Transfer Dynamics of Brown Algal Photosynthetic Antennas. J. Phys. Chem. Lett. 2012, 3, 2659–2664. [Google Scholar] [CrossRef]
- Romero, E.; Augulis, R.; Novoderezhkin, V.; Ferretti, M.; Thieme, J.; Zigmantas, D.; Van Grondelle, R. Quantum coherence in photosynthesis for efficient solar-energy conversion. Nat. Phys. 2014, 10, 676–682. [Google Scholar] [CrossRef]
- Gelzinis, A.; Butkus, V.; Songaila, E.; Augulis, R.; Gall, A.; Büchel, C.; Robert, B.; Abramavicius, D.; Zigmantas, D.; Valkunas, L. Mapping energy transfer channels in fucoxanthin–chlorophyll protein complex. Biochim. Biophys. Acta Bioenerg. 2014, 1847, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Brixner, T.; Stenger, J.; Vaswani, H.M.; Cho, M.; Blankenship, R.E.; Fleming, G.R. Two-dimensional spectroscopy of electronic couplings in photosynthesis. Nature 2005, 434, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Lambrev, P.H.; Akhtar, P.; Tan, H.-S. Insights into the mechanisms and dynamics of energy transfer in plant light-harvesting complexes from two-dimensional electronic spectroscopy. Biochim. Biophys. Acta Bioenerg. 2019, 1861, 148050. [Google Scholar] [CrossRef] [PubMed]
- Lavaud, J.; Rousseau, B.; Etienne, A.-L. Enrichment of the Light-Harvesting Complex in Diadinoxanthin and Implications for the Nonphotochemical Fluorescence Quenching in Diatoms. Biochemistry 2003, 42, 5802–5808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavaud, J.; Rousseau, B.; van Gorkom, H.J.; Etienne, A.-L. Influence of the Diadinoxanthin Pool Size on Photoprotection in the Marine Planktonic Diatom Phaeodactylum tricornutum. Plant Physiol. 2002, 129, 1398–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundermann, K.; Büchel, C. The fluorescence yield of the trimeric fucoxanthin–chlorophyll–protein FCPa in the diatom Cyclotella meneghiniana is dependent on the amount of bound diatoxanthin. Photosynth. Res. 2007, 95, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.; Pinto, E.A.; Wilhelm, C.; Richter, M. The Importance of a Highly Active and ΔpH-Regulated Diatoxanthin Epoxidase for the Regulation of the PS II Antenna Function in Diadinoxanthin Cycle Containing Algae. J. Plant Physiol. 2006, 163, 1008–1021. [Google Scholar] [CrossRef]
- Gundermann, K.; Büchel, C. Factors determining the fluorescence yield of fucoxanthin-chlorophyll complexes (FCP) involved in non-photochemical quenching in diatoms. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Grouneva, I.; Jakob, T.; Wilhelm, C.; Goss, R. A New Multicomponent NPQ Mechanism in the Diatom Cyclotella meneghiniana. Plant Cell Physiol. 2008, 49, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Taddei, L.; Chukhutsina, V.U.; Lepetit, B.; Stella, G.R.; Bassi, R.; Van Amerongen, H.; Bouly, J.-P.; Jaubert, M.; Finazzi, G.; Falciatore, A. Dynamic Changes between Two LHCX-Related Energy Quenching Sites Control Diatom Photoacclimation. Plant Physiol. 2018, 177, 953–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, A.; Gundermann, K.; Beckmann, J.; Büchel, C. Subunit Composition and Pigmentation of Fucoxanthin−Chlorophyll Proteins in Diatoms: Evidence for a Subunit Involved in Diadinoxanthin and Diatoxanthin Binding. Biochemistry 2006, 45, 13046–13053. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.; Bergantino, E.; Giacometti, G.M. Light and oxygenic photosynthesis: Energy dissipation as a protection mechanism against photo-oxidation. EMBO Rep. 2005, 6, 629–634. [Google Scholar] [CrossRef]
- Büchel, C. Light harvesting complexes in chlorophyll c-containing algae. Biochim. Biophys. Acta Bioenerg. 2019, 1861, 148027. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Jung, Y.-J.; Kwon, O.-N.; Cha, K.H.; Um, B.-H.; Chung, D.; Pan, C.-H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.; Chan, K.W.; Ebrahimi, M. Antioxidant capacities of fucoxanthin-producing algae as influenced by their carotenoid and phenolic contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, V.; Chérouvrier, J.-R.; Farhat, F.; Thiéry, V.; Piot, J.M.; Bérard, J.-B.; Kaas, R.; Serive, B.; Patrice, T.; Cadoret, J.-P.; et al. Study on the microalgal pigments extraction process: Performance of microwave assisted extraction. Process. Biochem. 2011, 46, 59–67. [Google Scholar] [CrossRef] [Green Version]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An investigation into the effect of culture conditions on fucoxanthin production using the marine microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Guo, B.; Liu, B.; Yang, B.; Sun, P.; Lu, X.; Liu, J.; Chen, F. Screening of Diatom Strains and Characterization of Cyclotella cryptica as A Potential Fucoxanthin Producer. Mar. Drugs 2016, 14, 125. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Wang, X.; Liu, J. Screening of Isochrysis strains for simultaneous production of docosahexaenoic acid and fucoxanthin. Algal Res. 2019, 41, 101545. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Khaw, Y.S.; Khong, N.M.H.; Shaharuddin, N.A.; Yusoff, F.M. A simple 18S rDNA approach for the identification of cultured eukaryotic microalgae with an emphasis on primers. J. Microbiol. Methods 2020, 172, 105890. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Falciatore, A.; Casotti, R.; Leblanc, C.; Abrescia, C.; Bowler, C. Transformation of Nonselectable Reporter Genes in Marine Diatoms. Mar. Biotechnol. 1999, 1, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Apt, K.E.; Grossman, A.R.; Kroth-Pancic, P.G. Stable nuclear transformation of the diatom Phaeodactylum tricornutum. Mol. Genet. Genom. 1996, 252, 572–579. [Google Scholar] [CrossRef]
- Miyagawa, A.; Okami, T.; Kira, N.; Yamaguchi, H.; Ohnishi, K.; Adachi, M. Research Note: High Efficiency Transformation of the Diatom Phaeodactylum tricornutum with a Promoter from the Diatom Cylindrotheca fusiformis. Phycol. Res. 2009, 57, 142–146. [Google Scholar] [CrossRef]
- Miyagawa-Yamaguchi, A.; Okami, T.; Kira, N.; Yamaguchi, H.; Ohnishi, K.; Adachi, M. Stable nuclear transformation of the diatom Chaetoceros sp. Phycol. Res. 2011, 59, 113–119. [Google Scholar] [CrossRef]
- Erdene-ochir, E.; Shin, B.; Huda, N.; Hye, D.; Eun, K.; Lee, H. Cloning of a Novel Endogenous Promoter for Foreign Gene Expression in Phaeodactylum tricornutum. Appl. Biol. Chem. 2016, 59, 861–867. [Google Scholar] [CrossRef]
- Adler-Agnon, Z.; Leu, S.; Zarka, A.; Boussiba, S.; Khozin-Goldberg, I. Novel promoters for constitutive and inducible expression of transgenes in the diatom Phaeodactylum tricornutum under varied nitrate availability. Environ. Boil. Fishes 2017, 30, 2763–2772. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kadono, T.; Kira, N.; Suzuki, K.; Iwata, O.; Ohnishi, K.; Yamaguchi, H.; Adachi, M. Development of endogenous promoters that drive high-level expression of introduced genes in the model diatom Phaeodactylum tricornutum. Mar. Genom. 2018, 42, 41–48. [Google Scholar] [CrossRef]
- Mikami, K.; Hosokawa, M. Biosynthetic Pathway and Health Benefits of Fucoxanthin, an Algae-Specific Xanthophyll in Brown Seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iravani, N.; Hajiaghaee, R.; Zarekarizi, A. A Review on Biosynthesis, Health Benefits and Extraction Methods of Fucoxanthin, Particular Marine Carotenoids in Algae. J. Med. Plants 2018, 17, 6–30. [Google Scholar]
- Zarekarizi, A.; Hoffmann, L.; Burritt, D. Approaches for the sustainable production of fucoxanthin, a xanthophyll with potential health benefits. Environ. Boil. Fishes 2018, 31, 281–299. [Google Scholar] [CrossRef]
- Eilers, U.; Bikoulis, A.; Breitenbach, J.; Büchel, C.; Sandmann, G. Limitations in the biosynthesis of fucoxanthin as targets for genetic engineering in Phaeodactylum tricornutum. Environ. Boil. Fishes 2015, 28, 123–129. [Google Scholar] [CrossRef]
- Kadono, T.; Kira, N.; Suzuki, K.; Iwata, O.; Ohama, T.; Okada, S.; Nishimura, T.; Akakabe, M.; Tsuda, M.; Adachi, M. Effect of an Introduced Phytoene Synthase Gene Expression on Carotenoid Biosynthesis in the Marine Diatom Phaeodactylum tricornutum. Mar. Drugs 2015, 13, 5334–5357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.M.; Shang, Y.F.; Um, B.-H. A preparative method for isolation of fucoxanthin from Eisenia bicyclis by centrifugal partition chromatography. Phytochem. Anal. 2011, 22, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Tokarek, W.; Listwan, S.; Pagacz, J.; Leśniak, P.; Latowski, D. Column chromatography as a useful step in purification of diatom pigments. Acta Biochim. Pol. 2016, 63, 443–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.F.; Kim, S.M.; Lee, W.J.; Um, B.-H. Pressurized liquid method for fucoxanthin extraction from Eisenia bicyclis (Kjellman) Setchell. J. Biosci. Bioeng. 2011, 111, 237–241. [Google Scholar] [CrossRef]
- Quitain, A.T.; Kai, T.; Sasaki, M.; Goto, M. Supercritical Carbon Dioxide Extraction of Fucoxanthin from Undaria pinnatifida. J. Agric. Food Chem. 2013, 61, 5792–5797. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.J.; Kim, Y.E.; Kim, J.-E.; Park, J.; Kim, Y.H.; Song, K.-M.; Lee, N.H. Production of Undaria pinnatifida sporophyll extract using pilot-scale ultrasound-assisted extraction: Extract characteristics and antioxidant and anti-inflammatory activities. Algal Res. 2020, 51, 102039. [Google Scholar] [CrossRef]
- Jaswir, I.; Noviendri, D.; Salleh, H.M.; Taher, M.; Miyashita, K. Isolation of Fucoxanthin and Fatty Acids Analysis of Padina australis and Cytotoxic Effect of Fucoxanthin on Human Lung Cancer (H1299) Cell Lines. Afr. J. Biotechnol. 2011, 10, 18855–18862. [Google Scholar]
- Noviendri, D.; Salleh, H.M.; Taher, M.; Miyashita, K.; Ramli, N. Fucoxanthin Extraction and Fatty Acid Analysis of Sargassum binderi and Sargassum duplicatum. J. Med. Plants Res. 2011, 5, 2405–2412. [Google Scholar]
- Heffernan, N.; Smyth, T.J.; FitzGerald, R.J.; Vila-Soler, A.; Mendiola, J.A.; Ibáñez, E.; Brunton, N. Comparison of extraction methods for selected carotenoids from macroalgae and the assessment of their seasonal/spatial variation. Innov. Food Sci. Emerg. Technol. 2016, 37, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.T.; Khong, N.M.H.; Khaw, Y.S.; Ahmad, S.A.; Yusoff, F.M. Optimization of the Freezing-Thawing Method for Extracting Phycobiliproteins from Arthrospira sp. Molecules 2020, 25, 3894. [Google Scholar] [CrossRef]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative Alternative Technologies to Extract Carotenoids from Microalgae and Seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef] [PubMed]
- Lourenço-Lopes, C.; Garcia-Oliveira, P.; Carpena, M.; Fraga-Corral, M.; Jimenez-Lopez, C.; Pereira, A.; Prieto, M.; Simal-Gandara, J. Scientific Approaches on Extraction, Purification and Stability for the Commercialization of Fucoxanthin Recovered from Brown Algae. Foods 2020, 9, 1113. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.M. Pigment Diversity in Freshwater Phytoplankton. II. Summer-succession in Three Dutch Lakes with Different Trophic Characteristics. Int. Rev. der gesamten Hydrobiol. Hydrogr. 1977, 62, 19–39. [Google Scholar] [CrossRef]
- Jeffrey, S.W. Profiles of photosynthetic pigments in the ocean using thin-layer chromatography. Mar. Biol. 1974, 26, 101–110. [Google Scholar] [CrossRef]
- Barlow, R.G.; Mantoura, R.F.C.; Peinert, R.D.; Miller, A.E.J.; Fileman, T.W. Distribution, sedimentation and fate of pigment biomarkers following thermal stratification in the western Alboran Sea. Mar. Ecol. Prog. Ser. 1995, 125, 279–291. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, J.; Xiao, L. Response of Phytoneuston Community to Organic Nitrogen and Phosphorus Revealed by HPLC-Pigments Method. J. Ocean Univ. China 2020, 19, 853–864. [Google Scholar] [CrossRef]
- Gieskes, W.W.; Kraay, G.W. Analysis of phytoplankton pigments by HPLC before, during and after mass occurrence of the microflagellate Corymbellus aureus during the spring bloom in the open northern North Sea in 1983. Mar. Biol. 1986, 92, 45–52. [Google Scholar] [CrossRef]
- Terzic, S.; Ahel, M.; Cauwet, G.; Malej, A. Group-specific phytoplankton biomass/dissolved carbohydrate relationships in the Gulf of Trieste (Northern Adriatic). Hydrobiologia 1997, 363, 191–205. [Google Scholar] [CrossRef]
- Van De Vyver, E.; Van Wichelen, J.; Vanormelingen, P.; Van Nieuwenhuyze, W.; Daveloose, I.; De Jong, R.; De Blok, R.; Urrutia, R.; Tytgat, B.; Verleyen, E.; et al. Variation in phytoplankton pigment composition in relation to mixing conditions in temperate South-Central Chilean lakes. Limnologica 2019, 79, 125715. [Google Scholar] [CrossRef]
- Züllig, H. Role of Carotenoids in Lake Sediments for Reconstructing Trophic History during the Late Quaternary; Springer: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Jiménez, L.; Romero-Viana, L.; Conde-Porcuna, J.M.; Pérez-Martínez, C. Sedimentary photosynthetic pigments as indicators of climate and watershed perturbations in an alpine lake in southern Spain. Limnetica 2015, 34, 439–454. [Google Scholar] [CrossRef]
- Paanakker, J.E.; Hallegraeff, G.M. A comparative study on the carotenoid pigmentation of the zooplankton of Lake Maarsseveen (Netherlands) and of Lac Pavin (Auvergne, France)—I. Chromatographic characterization of carotenoid pigments. Comp. Biochem. Physiol. Part B Comp. Biochem. 1978, 60, 51–58. [Google Scholar] [CrossRef]
- Quiblier-Lloberas, C.; Bourdier, G.; Amblard, C.; Pepin, D. Impact of grazing on phytoplankton in Lake Pavin (France): Contribution of different zooplankton groups. J. Plankton Res. 1996, 18, 305–322. [Google Scholar] [CrossRef] [Green Version]
- Abidov, M.; Ramazanov, Z.; Seifulla, R.; Grachev, S. The Effects of XanthigenTM in the Weight Management of Obese Premenopausal Women with Non-alcoholic Fatty Liver Disease and Normal Liver Fat. Diabetes Obes. Metab. 2010, 12, 72–81. [Google Scholar] [CrossRef]
- Lai, C.S.; Tsai, M.L.; Badmaev, V.; Jimenez, M.; Ho, C.T.; Pan, M.H. Xanthigen Suppresses Preadipocyte Differentiation and Adipogenesis through Down-Regulation of PPARγ and C/EBPs and Modulation of SIRT-1, AMPK, and FoxO Pathways. J. Agric. Food Chem. 2012, 60, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- López-Rios, L.; Vega, T.; Chirino, R.; Jung, J.C.; Davis, B.; Pérez-Machín, R.; Wiebe, J.C. Toxicological Assessment of Xanthigen® Nutraceutical Extract Combination: Mutagenicity, Genotoxicity and Oral Toxicity. Toxicol. Rep. 2018, 5, 1021–1031. [Google Scholar] [CrossRef]
- Anand, M.; Suresh, S. Marine seaweed Sargassum wightii extract as a low-cost sensitizer for ZnO photoanode based dye-sensitized solar cell. Adv. Nat. Sci. Nanosci. Nanotechnol. 2015, 6, 035008. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, C.; Dong, W.; Wang, W.; Sun, Y.; Chen, X.; Yang, C.; Dai, N. Photoelectrochemical Complexes of Fucoxanthin-Chlorophyll Protein for Bio-Photovoltaic Conversion with a High Open-Circuit Photovoltage. Chem. Asian J. 2017, 12, 2996–2999. [Google Scholar] [CrossRef]
- Rodriguez-Luna, A.; Ávila-Román, J.; González-Rodríguez, M.L.; Cózar, M.J.; Rabasco, A.M.; Motilva, V.; Talero, E. Fucoxanthin-Containing Cream Prevents Epidermal Hyperplasia and UVB-Induced Skin Erythema in Mice. Mar. Drugs 2018, 16, 378. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-J.; Nam, G.-W. Sunscreen Boosting Effect by Solid Lipid Nanoparticles-Loaded Fucoxanthin Formulation. Cosmetics 2020, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.Y.; Kang, H.; Lee, J.E.; Jo, C.S.; Moon, C.B.; Ha, J.; Hwang, J.S.; Choi, J. Antiaging Potential of Fucoxanthin Concentrate Derived from Phaeodactylum tricornutum. J. Cosmet. Sci. 2020, 71, 53–64. [Google Scholar]
- Sasaki, K.; Ishihara, K.; Yamazaki, M.; Nakashima, K.; Abe, H.; Oyamada, C.; Motoyama, M.; Mitsumoto, M. Oral Administration of Fucoxanthin Increases Plasma Fucoxanthinol Concentration and Antioxidative Ability and Improves Meat Color in Broiler Chicks. J. Poult. Sci. 2010, 47, 316–320. [Google Scholar] [CrossRef] [Green Version]
- Gumus, R.; Gelen, S.U.; Koseoglu, S.; Ozkanlar, S.; Ceylan, Z.; Imik, H. The Effects of Fucoxanthin Dietary Inclusion on the Growth Performance, Antioxidant Metabolism and Meat Quality of Broilers. Brazilian J. Poult. Sci. 2018, 20, 487–496. [Google Scholar] [CrossRef]
- Jensen, A. The Effect of Seaweed Carotenoids on Egg Yolk Coloration. Poult. Sci. 1963, 42, 912–916. [Google Scholar] [CrossRef]
- Prabhasankar, P.; Ganesan, P.; Bhaskar, N.; Hirose, A.; Stephen, N.; Gowda, L.R.; Hosokawa, M.; Miyashita, K. Edible Japanese seaweed, wakame (Undaria pinnatifida) as an ingredient in pasta: Chemical, functional and structural evaluation. Food Chem. 2009, 115, 501–508. [Google Scholar] [CrossRef]
- Mok, I.-K.; Yoon, J.-R.; Pan, C.-H.; Kim, S.M. Development, Quantification, Method Validation, and Stability Study of a Novel Fucoxanthin-Fortified Milk. J. Agric. Food Chem. 2016, 64, 6196–6202. [Google Scholar] [CrossRef]
- Mok, I.-K.; Lee, J.K.; Kim, J.H.; Pan, C.-H.; Kim, S.M. Fucoxanthin bioavailability from fucoxanthin-fortified milk: In vivo and in vitro study. Food Chem. 2018, 258, 79–86. [Google Scholar] [CrossRef]
- Yamano, Y.; Tode, C.; Ito, M. Carotenoids and related polyenes. Part 3. First total synthesis of fucoxanthin and halocynthiaxanthin using oxo-metallic catalyst. J. Chem. Soc. Perkin Trans. 1 1995, 15, 1895–1904. [Google Scholar] [CrossRef]
- Seth, K.; Kumar, A.; Rastogi, R.P.; Meena, M.; Vinayak, V. Harish Bioprospecting of fucoxanthin from diatoms—Challenges and perspectives. Algal Res. 2021, 60, 102475. [Google Scholar] [CrossRef]
- Vella, F.M.; Sardo, A.; Gallo, C.; Landi, S.; Fontana, A.; D’Ippolito, G. Annual outdoor cultivation of the diatom Thalassiosira weissflogii: Productivity, limits and perspectives. Algal Res. 2019, 42, 101553. [Google Scholar] [CrossRef]
- Arora, N.; Philippidis, G.P. Fucoxanthin Production from Diatoms: Current Advances and Challenges. In Algae; Springer: Singapore, 2021; pp. 227–242. [Google Scholar] [CrossRef]
- García, J.L.; De Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Ravi, H.; Kurrey, N.; Manabe, Y.; Sugawara, T.; Baskaran, V. Polymeric chitosan-glycolipid nanocarriers for an effective delivery of marine carotenoid fucoxanthin for induction of apoptosis in human colon cancer cells (Caco-2 cells). Mater. Sci. Eng. C 2018, 91, 785–795. [Google Scholar] [CrossRef]
- Koo, S.Y.; Mok, I.-K.; Pan, C.-H.; Kim, S.M. Preparation of Fucoxanthin-Loaded Nanoparticles Composed of Casein and Chitosan with Improved Fucoxanthin Bioavailability. J. Agric. Food Chem. 2016, 64, 9428–9435. [Google Scholar] [CrossRef]
- Lange, K.W.; Hauser, J.; Nakamura, Y.; Kanaya, S. Dietary seaweeds and obesity. Food Sci. Hum. Wellness 2015, 4, 87–96. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | TP | Number of Publications | |||||
|---|---|---|---|---|---|---|---|
| 1928–1970 | 1971–1980 | 1981–1990 | 1991–2000 | 2001–2010 | 2011–2021 * | ||
| Japan | 379 | 0 | 4 | 13 | 37 | 98 | 227 |
| United States | 319 | 8 | 16 | 31 | 71 | 78 | 115 |
| China | 230 | 0 | 0 | 1 | 1 | 9 | 219 |
| Germany | 181 | 0 | 1 | 3 | 32 | 45 | 100 |
| France | 144 | 1 | 0 | 4 | 32 | 48 | 59 |
| United Kingdom | 132 | 5 | 6 | 5 | 28 | 26 | 62 |
| South Korea | 127 | 0 | 0 | 0 | 1 | 16 | 110 |
| Spain | 99 | 0 | 0 | 1 | 9 | 22 | 67 |
| India | 98 | 0 | 0 | 0 | 3 | 11 | 84 |
| Canada | 75 | 0 | 2 | 5 | 19 | 25 | 24 |
| Institution | Country | TP | Number of Publications | |||||
|---|---|---|---|---|---|---|---|---|
| 1928–1970 | 1971–1980 | 1981–1990 | 1991–2000 | 2001–2010 | 2011–2021 * | |||
| Hokkaido University | Japan | 110 | 0 | 0 | 1 | 8 | 33 | 68 |
| CNRS Centre National de la Recherche Scientifique | France | 61 | 1 | 0 | 1 | 17 | 17 | 25 |
| Goethe-Universität Frankfurt am Main | Germany | 49 | 0 | 0 | 0 | 3 | 13 | 33 |
| Norges teknisk-naturvitenskapelige universitet | Norway | 44 | 0 | 7 | 12 | 12 | 8 | 5 |
| Chinese Academy of Sciences | China | 41 | 0 | 0 | 1 | 1 | 2 | 37 |
| Sorbonne Universite | France | 39 | 0 | 0 | 1 | 10 | 14 | 14 |
| Kyoto University | Japan | 34 | 0 | 0 | 3 | 8 | 9 | 14 |
| Kobe University | Japan | 34 | 0 | 0 | 0 | 1 | 7 | 26 |
| Plymouth Marine Laboratory | United Kingdom | 33 | 0 | 0 | 1 | 15 | 6 | 11 |
| Pukyong National University | Korea | 33 | 0 | 0 | 0 | 1 | 2 | 30 |
| Author | Country | TP | Scopus h-Index | Scopus ID | Number of Publications | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1928–1970 | 1971–1980 | 1981–1990 | 1991–2000 | 2001–2010 | 2011–2021 * | |||||
| Miyashita, Kazuo | Japan | 88 | 50 |
| 0 | 0 | 0 | 0 | 29 | 59 |
| Hosokawa, Masashi | Japan | 68 | 44 |
| 0 | 0 | 0 | 1 | 25 | 42 |
| Buchel, Claudia | Germany | 41 | 34 |
| 0 | 0 | 0 | 0 | 13 | 28 |
| Maeda, Hayato | Japan | 31 | 19 |
| 0 | 0 | 0 | 0 | 11 | 20 |
| Liaaen-Jensen, Synnove | Norway | 29 | 40 |
| 0 | 6 | 11 | 10 | 2 | 0 |
| Nagao, Ryo | Japan | 28 | 21 |
| 0 | 0 | 0 | 0 | 2 | 26 |
| Akimoto, Seiji | Japan | 23 | 27 |
| 0 | 0 | 0 | 0 | 0 | 23 |
| Shen, Jian Ren | Japan | 22 | 50 |
| 0 | 0 | 0 | 0 | 0 | 22 |
| Maoka, Takashi | Japan | 19 | 37 |
| 0 | 0 | 0 | 1 | 5 | 13 |
| Hashimoto, Hideki | Japan | 18 | 44 |
| 0 | 0 | 0 | 0 | 2 | 16 |
| Subject Category | TP (%) |
|---|---|
| Agricultural and Biological Sciences | 27.0 |
| Biochemistry, Genetics and Molecular Biology | 18.1 |
| Pharmacology, Toxicology and Pharmaceutics | 8.3 |
| Chemistry | 7.9 |
| Environmental Science | 7.8 |
| Earth and Planetary Sciences | 7.5 |
| Medicine | 5.8 |
| Chemical Engineering | 3.8 |
| Immunology and Microbiology | 2.5 |
| Engineering | 2.3 |
| Journal | Publisher | IF (2020) | TP | TP (%) | Cummulative TP (%) |
|---|---|---|---|---|---|
| Marine Drugs | MDPI | 5.118 | 123 | 5.91 | 5.91 |
| Journal of Phycology | Wiley Periodical LLC | 2.923 | 67 | 3.22 | 9.13 |
| Journal of Applied Phycology | Springer | 3.215 | 55 | 2.64 | 11.77 |
| Photosynthesis Research | Springer | 3.573 | 39 | 1.88 | 13.65 |
| Biochimica Et Biophysica Acta (BBA)—Bioenergetics | Elsevier B.V. | 3.991 | 38 | 1.83 | 15.48 |
| Algal Research | Elsevier B.V. | 4.401 | 32 | 1.54 | 17.02 |
| Marine Ecology Progress Series | Inter-Research Science | 2.824 | 30 | 1.44 | 18.46 |
| Journal of Agricultural and Food Chemistry | American Chemical Society (ACS) | 5.279 | 30 | 1.44 | 19.90 |
| Journal of Plankton Research | Oxford University Press (OUP) | 2.455 | 24 | 1.15 | 21.05 |
| Marine Biology | Springer | 2.573 | 24 | 1.15 | 22.20 |
| Title | Authors | Year | Journal | Total Citation |
|---|---|---|---|---|
| Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues | Maeda et al. | 2005 | Biochemical and Biophysical Research Communications | 411 |
| Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health | Peng et al. | 2011 | Marine Drugs | 356 |
| Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed | Yan et al. | 1999 | Bioscience, Biotechnology and Biochemistry | 348 |
| Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites | Sachindra et al. | 2007 | Journal of Agricultural and Food Chemistry | 308 |
| Fucoxanthin induces apoptosis and enhances the antiproliferative effect of the PPARγ ligand, troglitazone, on colon cancer cells | Hosokawa et al. | 2004 | Biochimica et Biophysica Acta (BBA)—General Subjects | 261 |
| Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice | Maeda et al. | 2007 | Journal of Agricultural and Food Chemistry | 226 |
| A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum | Kim et al. | 2012 | Applied Biochemistry and Biotechnology | 218 |
| Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages | Heo et al. | 2010 | Food and Chemical Toxicology | 212 |
| Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages | Kim et al. | 2010 | European Journal of Pharmacology | 204 |
| Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita | Xia et al. | 2013 | Marine Drugs | 178 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khaw, Y.S.; Yusoff, F.M.; Tan, H.T.; Noor Mazli, N.A.I.; Nazarudin, M.F.; Shaharuddin, N.A.; Omar, A.R. The Critical Studies of Fucoxanthin Research Trends from 1928 to June 2021: A Bibliometric Review. Mar. Drugs 2021, 19, 606. https://doi.org/10.3390/md19110606
Khaw YS, Yusoff FM, Tan HT, Noor Mazli NAI, Nazarudin MF, Shaharuddin NA, Omar AR. The Critical Studies of Fucoxanthin Research Trends from 1928 to June 2021: A Bibliometric Review. Marine Drugs. 2021; 19(11):606. https://doi.org/10.3390/md19110606
Chicago/Turabian StyleKhaw, Yam Sim, Fatimah Md. Yusoff, Hui Teng Tan, Nur Amirah Izyan Noor Mazli, Muhammad Farhan Nazarudin, Noor Azmi Shaharuddin, and Abdul Rahman Omar. 2021. "The Critical Studies of Fucoxanthin Research Trends from 1928 to June 2021: A Bibliometric Review" Marine Drugs 19, no. 11: 606. https://doi.org/10.3390/md19110606