Identification of a Key Enzyme for the Hydrolysis of β-(1→3)-Xylosyl Linkage in Red Alga Dulse Xylooligosaccharide from Bifidobacterium Adolescentis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
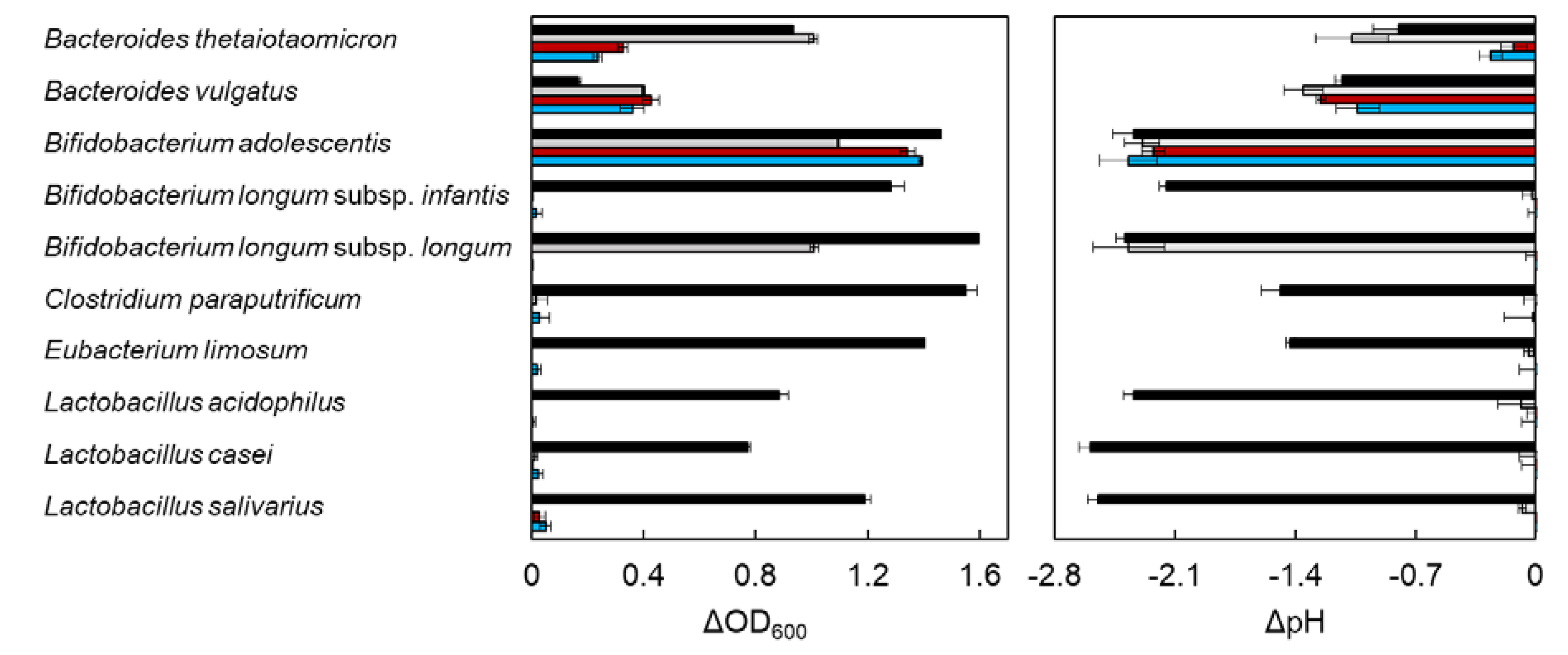
2.1. In Vitro Utilization of XOS by Entric Bacteria
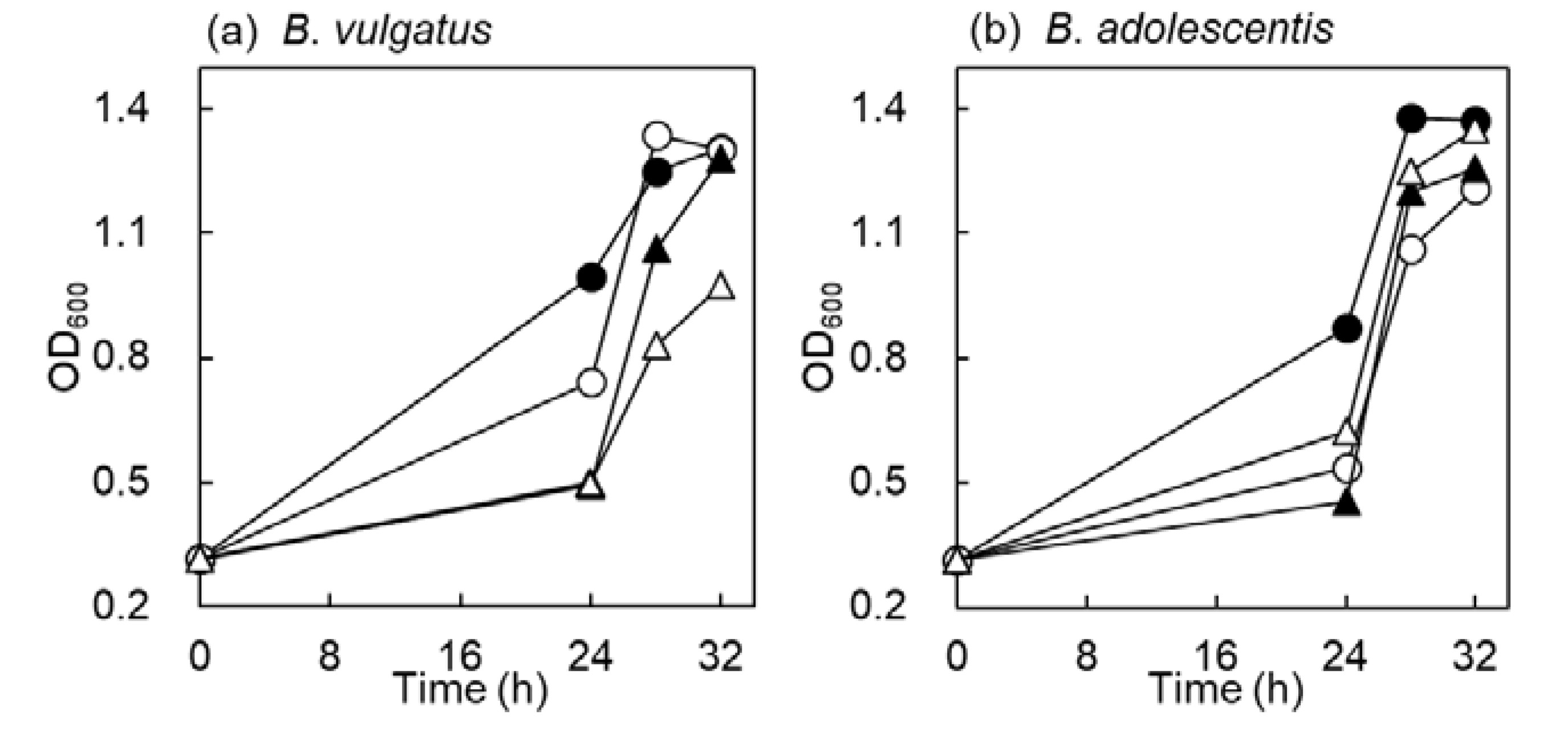
2.2. Time Course Growth in B. vulgatus and B. adolescentis
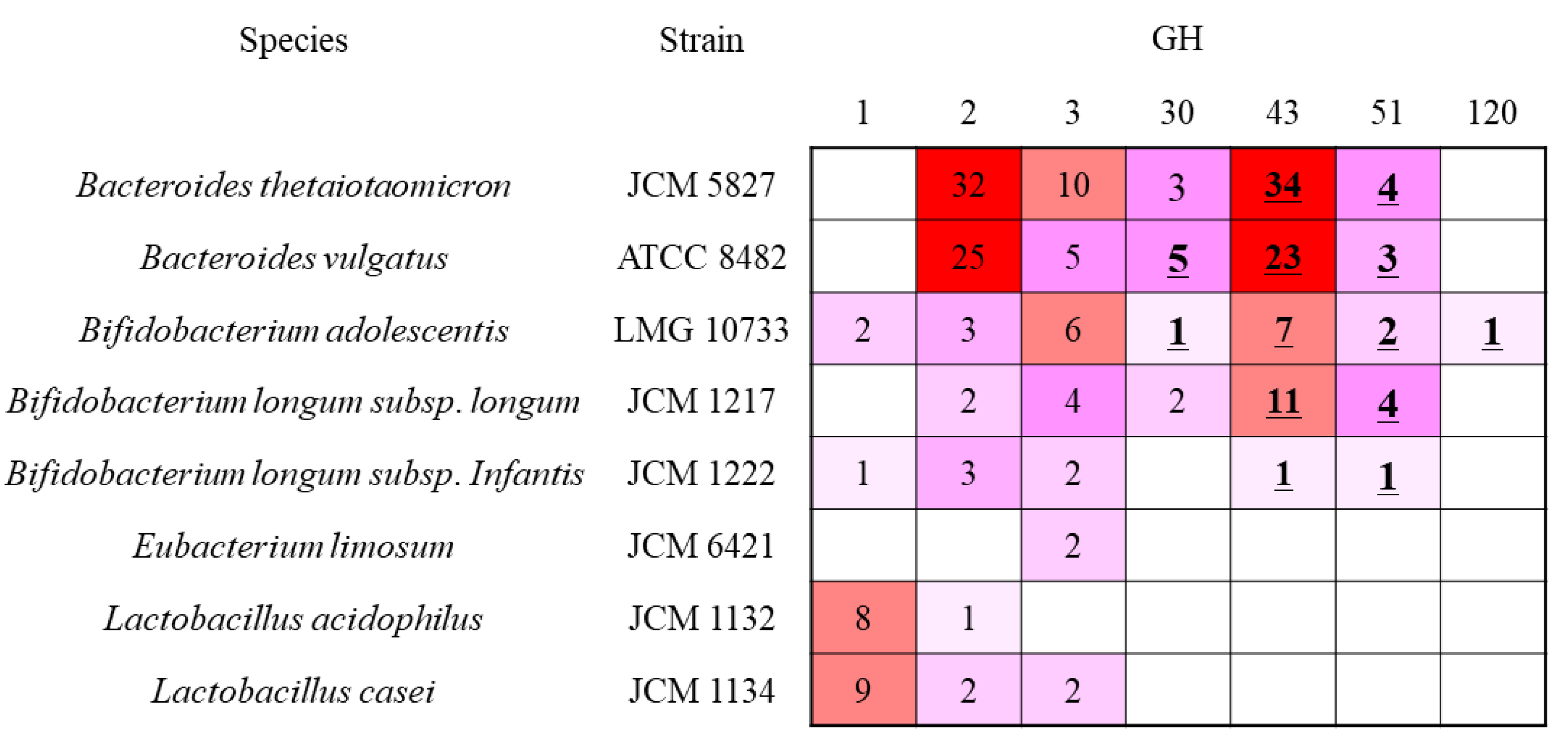
2.3. Comparison of β-Xylosidase Genes in the Tested Bacterium
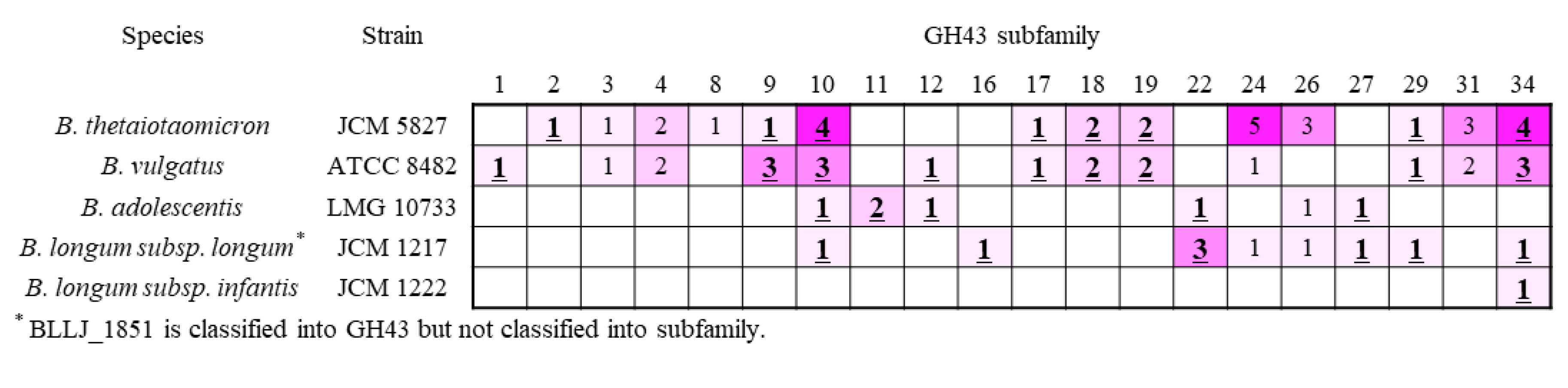
2.4. β-Xylosidase in GH43 Subfamily
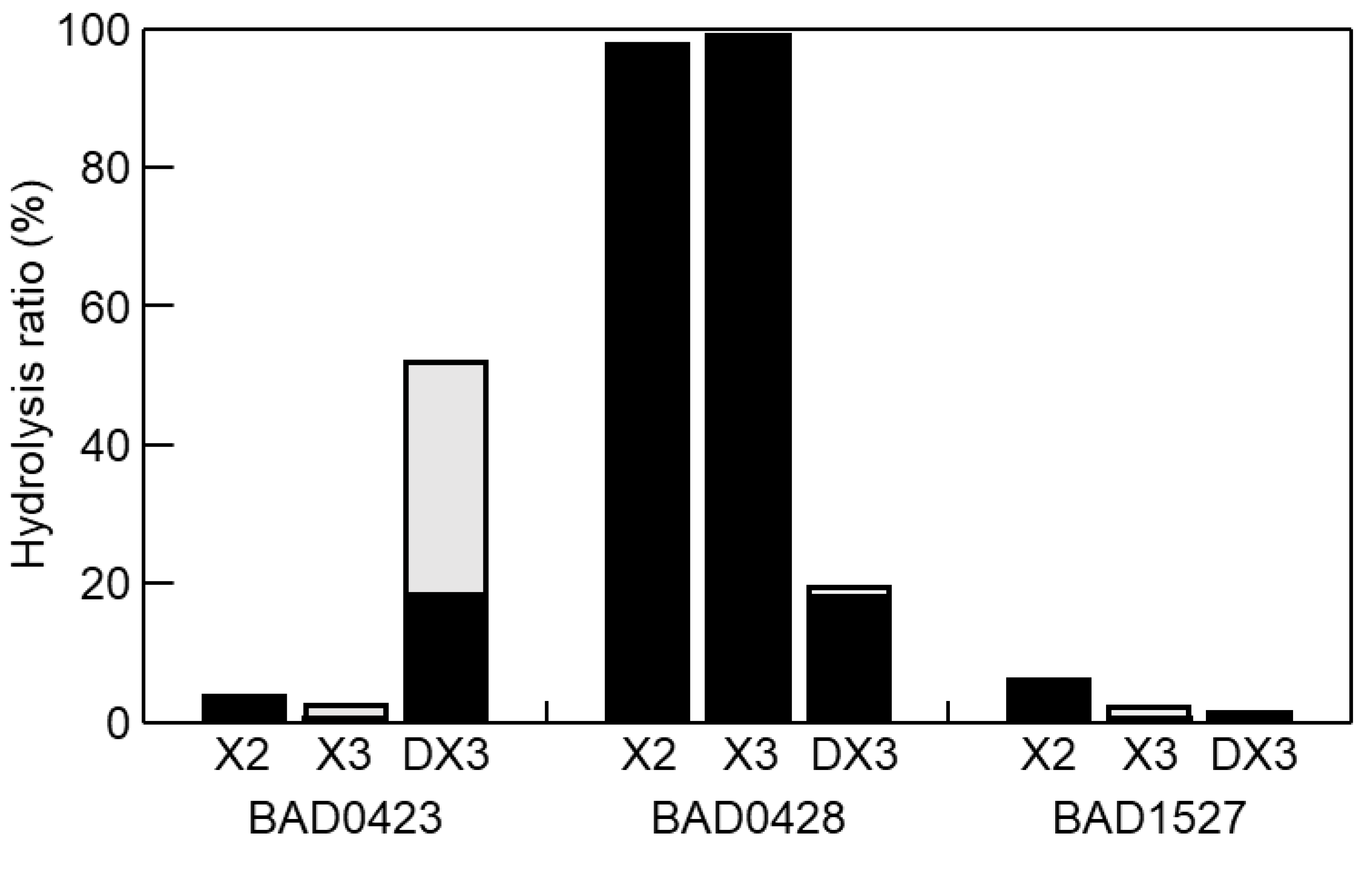
2.5. Enzymatic Character of GH43 Enzymes
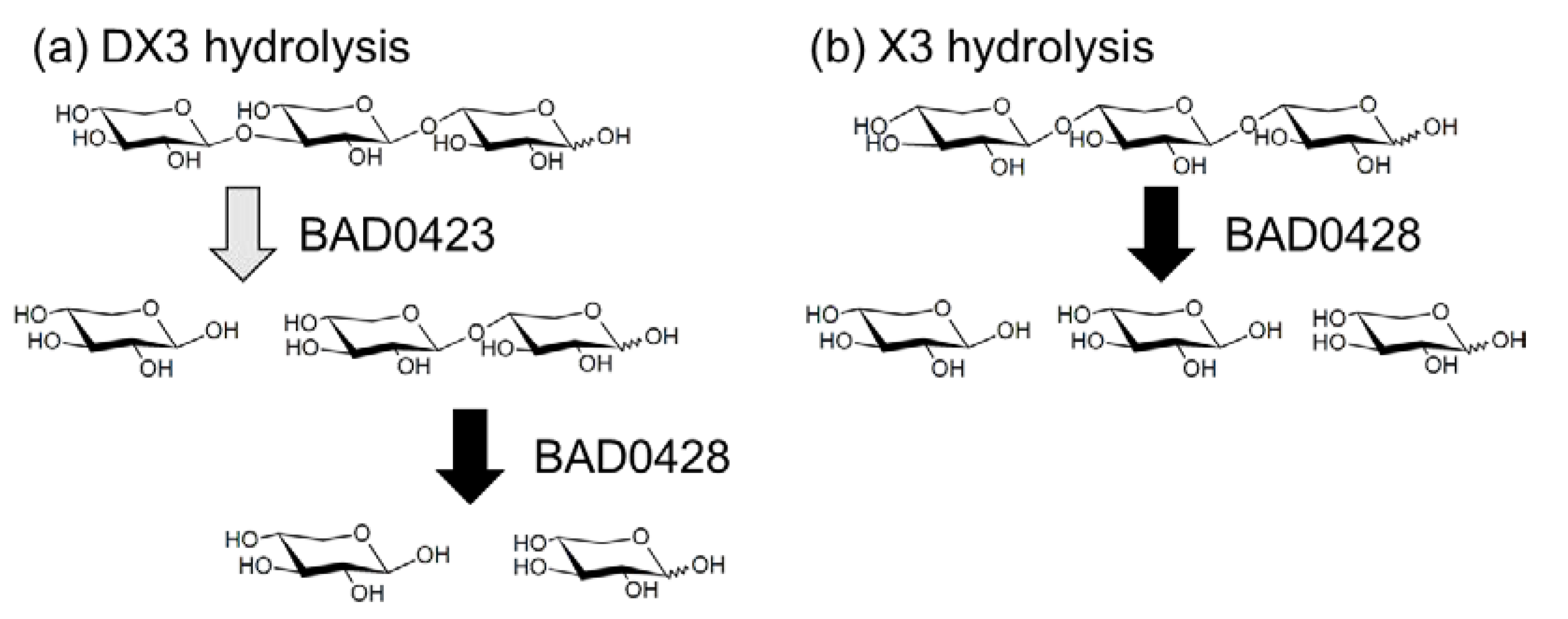
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of DX3 from Red Alga Dulse
4.3. Bacterial Growth
4.4. Bioinformatics
4.5. Cloning, Expression, and Purification of Enzymes
4.6. Activity Assay
4.7. HPLC Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ebringerová, A.; Hromádková, Z.; Heinze, T. Hemicellulose. In Polysaccharides I: Structure, Characterization and Use; Heinze, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 1–67. [Google Scholar]
- Carvalho, A.F.A.; de Oliva Neto, P.; da Silva, D.F.; Pastore, G.M. Xylo-oligosaccharides from lignocellulosic materials: Chemical structure, health benefits and production by chemical and enzymatic hydrolysis. Food Res. Int. 2013, 51, 75–85. [Google Scholar] [CrossRef]
- Singh, R.D.; Banerjee, J.; Arora, A. Prebiotic potential of oligosaccharides: A focus on xylan derived oligosaccharides. Bioact. Carbohydrates Diet. Fibre 2015, 5, 19–30. [Google Scholar] [CrossRef]
- Manisseri, C.; Gudipati, M. Prebiotic activity of purified xylobiose obtained from Ragi (Eleusine coracana, Indaf-15) bran. Indian J. Microbiol. 2012, 52, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.-Y.; Zhao, L.; Ge, X.-R.; Tang, Y.-L.; Chen, L.-J.; Pang, W.; Zhang, Y. Preparation, characterization and bioactivity of xylobiose and xylotriose from corncob xylan by xylanase. Eur. Food Res. Technol. 2015, 241, 27–35. [Google Scholar] [CrossRef]
- Sheu, W.H.-H.; Lee, I.-T.; Chen, W.; Chan, Y.-C. Effects of Xylooligosaccharides in type 2 diabetes mellitus. J. Nutr. Sci. Vitaminol. 2008, 54, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, M.; Fujikawa, S.; Matsumoto, N. Effect of xylooligosaccharide on the growth of Bifidobacteria. Bifidobact. Microflora 1990, 9, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Aachary, A.A.; Prapulla, S.G. Xylooligosaccharides (XOS) as an emerging prebiotic: Microbial synthesis, utilization, structural characterization, bioactive properties, and applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Akpinar, O.; Erdogan, K.; Bostanci, S. Production of xylooligosaccharides by controlled acid hydrolysis of lignocellulosic materials. Carbohydr. Res. 2009, 344, 660–666. [Google Scholar] [CrossRef]
- Martone, P.T.; Estevez, J.M.; Lu, F.; Ruel, K.; Denny, M.W.; Somerville, C.; Ralph, J. Discovery of lignin in seaweed reveals convergent evolution of cell-wall architecture. Curr. Biol. 2009, 19, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Hong, I.K.; Jeon, H.; Lee, S.B. Comparison of red, brown and green seaweeds on enzymatic saccharification process. J. Ind. Eng. Chem. 2014, 20, 2687–2691. [Google Scholar] [CrossRef]
- McCandless, E.L.; Craigie, J.S. Sulfated polysaccharides in red and brown algae. Ann. Rev. Plant Physiol. 1979, 30, 41–53. [Google Scholar] [CrossRef]
- Pereira, M.G.; Benevides, N.M.B.; Melo, M.R.S.; Valente, A.P.; Melo, F.R.; Mourão, P.A.S. Structure and anticoagulant activity of a sulfated galactan from the red alga, Gelidium crinale. Is there a specific structural requirement for the anticoagulant action? Carbohydr. Res. 2005, 340, 2015–2023. [Google Scholar] [CrossRef] [PubMed]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; McSorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from Seaweeds: An Ocean of Opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahaye, M.; Rondeau-Mouro, C.; Deniaud, E.; Buléon, A. Solid-state 13C NMR spectroscopy studies of xylans in the cell wall of Palmaria palmata (L. Kuntze, Rhodophyta). Carbohydr. Res. 2003, 338, 1559–1569. [Google Scholar] [CrossRef]
- Morgan, K.C.; Wright, J.L.C.; Simpson, F.J. Review of chemical constituents of the red alga Palmaria palmata (dulse). Econ. Bot. 1980, 34, 27–50. [Google Scholar] [CrossRef]
- Lahaye, M.; Michel, C.; Barry, J.L. Chemical, physicochemical and in-vitro fermentation characteristics of dietary fibres from Palmaria palmata (L.) Kuntze. Food Chem. 1993, 47, 29–36. [Google Scholar] [CrossRef]
- Deniaud, E.; Quemener, B.; Fleurence, J.; Lahaye, M. Structural studies of the mix-linked β-(1→3)/β-(1→4)-d-xylans from the cell wall of Palmaria palmata (Rhodophyta). Int. J. Biol. Macromol. 2003, 33, 9–18. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kishimura, H.; Kinoshita, Y.; Saburi, W.; Kumagai, Y.; Yasui, H.; Ojima, T. Enzymatic production of xylooligosaccharides from red alga dulse (Palmaria sp.) wasted in Japan. Process Biochem. 2019, 82, 117–122. [Google Scholar] [CrossRef]
- van den Broek, L.A.; Hinz, S.W.; Beldman, G.; Vincken, J.P.; Voragen, A.G. Bifidobacterium carbohydrases-their role in breakdown and synthesis of (potential) prebiotics. Mol. Nutr. Food Res. 2008, 52, 146–163. [Google Scholar] [CrossRef]
- Amaretti, A.; Bernardi, T.; Leonardi, A.; Raimondi, S.; Zanoni, S.; Rossi, M. Fermentation of xylo-oligosaccharides by Bifidobacterium adolescentis DSMZ 18350: Kinetics, metabolism, and β-xylosidase activities. Appl. Microbiol. Biotechnol. 2013, 97, 3109–3117. [Google Scholar] [CrossRef]
- Ejby, M.; Fredslund, F.; Vujicic-Zagar, A.; Svensson, B.; Slotboom, D.J.; Abou Hachem, M. Structural basis for arabinoxylo-oligosaccharide capture by the probiotic Bifidobacterium animalis subsp. lactis Bl-04. Mol. Microbiol. 2013, 90, 1100–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umemoto, Y.; Onishi, R.; Araki, T. Cloning of a novel gene encoding β-1,3-xylosidase from a marine bacterium, Vibrio sp. strain XY-214, and characterization of the gene product. Appl. Environ. Microbiol. 2008, 74, 305–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phuengmaung, P.; Fujiwara, D.; Sukhumsirichart, W.; Sakamoto, T. Identification and characterization of the first β-1,3-D-xylosidase from a gram-positive bacterium, Streptomyces sp. SWU10. Enzyme Microb. Technol. 2018, 112, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mewis, K.; Lenfant, N.; Lombard, V.; Henrissat, B. Dividing the large glycoside hydrolase family 43 into subfamilies: A motivation for detailed enzyme characterization. Appl. Environ. Microbiol. 2016, 82, 1686–1692. [Google Scholar] [CrossRef] [Green Version]
- Miyabe, Y.; Furuta, T.; Takeda, T.; Kanno, G.; Shimizu, T.; Tanaka, Y.; Gai, Z.; Yasui, H.; Kishimura, H. Structural properties of phycoerythrin from dulse Palmaria palmata. J. Food Biochem. 2017, 41, e12301. [Google Scholar] [CrossRef] [Green Version]
- Kitade, Y.; Miyabe, Y.; Yamamoto, Y.; Takeda, H.; Shimizu, T.; Yasui, H.; Kishimura, H. Structural characteristics of phycobiliproteins from red alga Mazzaella japonica. J. Food Biochem. 2018, 42, e12436. [Google Scholar] [CrossRef] [Green Version]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef]
- Kumagai, Y.; Miyabe, Y.; Takeda, T.; Adachi, K.; Yasui, H.; Kishimura, H. In silico analysis of relationship between proteins from plastid genome of red alga Palmaria sp. (Japan) and angiotensin I converting enzyme inhibitory peptides. Mar. Drugs 2019, 17, 190. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, Y.; Ryota, T.; Miyabe, Y.; Takeda, T.; Adachi, K.; Yasui, H.; Kishimura, H. Complete sequence of mitochondrial DNA of red alga dulse Palmaria palmata (Linnaeus) Weber & Mohr in Japan. Mitochondrial DNA Part B Resour. 2019, 4, 3177–3178. [Google Scholar]
- Sato, N.; Furuta, T.; Takeda, T.; Miyabe, Y.; Ura, K.; Takagi, Y.; Yasui, H.; Kumagai, Y.; Kishimura, H. Antioxidant activity of proteins extracted from red alga dulse harvested in Japan. J. Food Biochem. 2019, 43, e12709. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, B.; Cao, Y.; Wang, C. In vitro fermentation of xylooligosaccharides from wheat bran insoluble dietary fiber by Bifidobacteria. Carbohydr. Polym. 2010, 82, 419–423. [Google Scholar] [CrossRef]
- van Zanten, G.C.; Knudsen, A.; Röytiö, H.; Forssten, S.; Lawther, M.; Blennow, A.; Lahtinen, S.J.; Jakobsen, M.; Svensson, B.; Jespersen, L. The effect of selected synbiotics on microbial composition and short-chain fatty acid production in a model system of the Human colon. PLoS ONE 2012, 7, e47212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, E.J.; Smith, N.L.; Turkenburg, J.P.; D’Souza, S.; Gilbert, H.J.; Davies, G.J. Structural insight into the ligand specificity of a thermostable family 51 arabinofuranosidase, Araf51, from Clostridium thermocellum. Biochem. J. 2006, 395, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hövel, K.; Shallom, D.; Niefind, K.; Belakhov, V.; Shoham, G.; Baasov, T.; Shoham, Y.; Schomburg, D. Crystal structure and snapshots along the reaction pathway of a family 51 α-l-arabinofuranosidase. EMBO J. 2003, 22, 4922–4932. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Akiyama, T.; Hatano, H.; Kimura, K. In Vitro assessment of oligosaccharides assimilation by intestinal anaerobic bacteria. Milk Sci. 2015, 64, 87–98. [Google Scholar]
- Moura, P.; Barata, R.; Carvalheiro, F.; Gírio, F.; Loureiro-Dias, M.C.; Esteves, M.P. In vitro fermentation of xylo-oligosaccharides from corn cobs autohydrolysis by Bifidobacterium and Lactobacillus strains. LWT Food Sci. Technol. 2007, 40, 963–972. [Google Scholar] [CrossRef]
- Duranti, S.; Milani, C.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Sanchez, B.; Margolles, A.; et al. Evaluation of genetic diversity among strains of the human gut commensal Bifidobacterium adolescentis. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Komeno, M.; Hayamizu, H.; Fujita, K.; Ashida, H. Two novelα-l-arabinofuranosidases from Bifidobacterium longum subsp. longum belonging to glycoside hydrolase family 43 cooperatively degrade arabinan. Appl. Environ. Microbiol. 2019, 85, e02582-18. [Google Scholar]
- Lee, J.H.; Hyun, Y.J.; Kim, D.H. Cloning and characterization of α-l-arabinofuranosidase and bifunctional α-l-arabinopyranosidase/β-d-galactopyranosidase from Bifidobacterium longum H-1. J. Appl. Microbiol. 2011, 111, 1097–1107. [Google Scholar] [CrossRef]
- Milani, C.; Andrea Lugli, G.; Duranti, S.; Turroni, F.; Mancabelli, L.; Ferrario, C.; Mangifesta, M.; Hevia, A.; Viappiani, A.; Scholz, M.; et al. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut. Sci. Rep. 2015, 5, 15782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viborg, A.H.; Sørensen, K.I.; Gilad, O.; Steen-Jensen, D.B.; Dilokpimol, A.; Jacobsen, S.; Svensson, B. Biochemical and kinetic characterisation of a novel xylooligosaccharide-upregulated GH43 β-d-xylosidase/α-l-arabinofuranosidase (BXA43) from the probiotic Bifidobacterium animalis subsp. lactis BB-12. AMB Express 2013, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Sakamoto, A.; Kaneko, S.; Kotake, T.; Tsumuraya, Y.; Kitahara, K. Degradative enzymes for type II arabinogalactan side chains in Bifidobacterium longum subsp. longum. Appl. Microbiol. Biotechnol. 2019, 103, 1299–1310. [Google Scholar] [CrossRef] [PubMed]
- Van Laere, K.M.J.; Voragen, C.H.L.; Kroef, T.; Van Den Broek, L.A.M.; Beldman, G.; Voragen, A.G.J. Purification and mode of action of two different arabinoxylan arabinofuranohydrolases from Bifidobacterium adolescentis DSM 20083. Appl. Microbiol. Biotechnol. 1999, 51, 606–613. [Google Scholar] [CrossRef]
- Lagaert, S.; Pollet, A.; Delcour, J.A.; Lavigne, R.; Courtin, C.M.; Volckaert, G. Substrate specificity of three recombinant α-l-arabinofuranosidases from Bifidobacterium adolescentis and their divergent action on arabinoxylan and arabinoxylan oligosaccharides. Biochem. Biophys. Res. Commun. 2010, 402, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Lagaert, S.; Pollet, A.; Delcour, J.A.; Lavigne, R.; Courtin, C.M.; Volckaert, G. Characterization of two β-xylosidases from Bifidobacterium adolescentis and their contribution to the hydrolysis of prebiotic xylooligosaccharides. Appl. Microbiol. Biotechnol. 2011, 92, 1179–1185. [Google Scholar] [CrossRef]
- Andersen, J.M.; Barrangou, R.; Hachem, M.A.; Lahtinen, S.J.; Goh, Y.J.; Svensson, B.; Klaenhammer, T.R. Transcriptional analysis of oligosaccharide utilization by Bifidobacterium lactis Bl-04. BMC Genom. 2013, 14, 312. [Google Scholar] [CrossRef] [Green Version]
- Gilad, O.; Jacobsen, S.; Stuer-Lauridsen, B.; Pedersen, M.B.; Garrigues, C.; Svensson, B. Combined transcriptome and proteome analysis of Bifidobacterium animalis subsp. lactis BB-12 grown on xylo-oligosaccharides and a model of their utilization. Appl. Environ. Microbiol. 2010, 76, 7285–7291. [Google Scholar]
- Gilad, O.; Svensson, B.; Viborg, A.H.; Stuer-Lauridsen, B.; Jacobsen, S. The extracellular proteome of Bifidobacterium animalis subsp. lactis BB-12 reveals proteins with putative roles in probiotic effects. Proteomics 2011, 11, 2503–2514. [Google Scholar]
- Hyun, Y.J.; Kim, B.; Kim, D.H. Cloning and characterization of ginsenoside Ra1-hydrolyzing β-d-xylosidase from Bifidobacterium breve K-110. J. Microbiol. Biotechnol. 2012, 22, 535–540. [Google Scholar] [CrossRef] [Green Version]
- Margolles, A.; de los Reyes-Gavilán, C.G. Purification and Functional Characterization of a Novel α-l-Arabinofuranosidase from Bifidobacterium longum B667. Appl. Environ. Microbiol. 2003, 69, 5096–5103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, H.R.; Jørgensen, C.T.; Hansen, C.H.; Jørgensen, C.I.; Pedersen, S.; Meyer, A.S. A novel GH43 α-l-arabinofuranosidase from Humicola insolens: Mode of action and synergy with GH51 α-l-arabinofuranosidases on wheat arabinoxylan. Appl. Microbiol. Biotechnol. 2006, 73, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Pastell, H.; Westermann, P.; Meyer, A.S.; Päivi, T.; Tenkanen, M. In vitro fermentation of arabinoxylan-derived carbohydrates by bifidobacteria and mixed fecal microbiota. J. Agric. Food Chem. 2009, 57, 8598–8606. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.S.; Laville, E.; Xiao, Y.; Nouaille, S.; Le Bourgeois, P.; Heux, S.; Portais, J.C.; Monsan, P.; Martens, E.C.; Potocki-Veronese, G.; et al. Functional characterization of a gene locus from an uncultured gut Bacteroides conferring xylo-oligosaccharides utilization to Escherichia coli. Mol. Microbiol. 2016, 102, 579–592. [Google Scholar] [CrossRef]
- Kumagai, Y.; Usuki, H.; Yamamoto, Y.; Yamasato, A.; Arima, J.; Mukaihara, T.; Hatanaka, T. Characterization of calcium ion sensitive region for β-Mannanase from Streptomyces thermolilacinus. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 1127–1133. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, M.; Kumagai, Y.; Yamamoto, Y.; Yasui, H.; Kishimura, H. Identification of a Key Enzyme for the Hydrolysis of β-(1→3)-Xylosyl Linkage in Red Alga Dulse Xylooligosaccharide from Bifidobacterium Adolescentis. Mar. Drugs 2020, 18, 174. https://doi.org/10.3390/md18030174
Kobayashi M, Kumagai Y, Yamamoto Y, Yasui H, Kishimura H. Identification of a Key Enzyme for the Hydrolysis of β-(1→3)-Xylosyl Linkage in Red Alga Dulse Xylooligosaccharide from Bifidobacterium Adolescentis. Marine Drugs. 2020; 18(3):174. https://doi.org/10.3390/md18030174
Chicago/Turabian StyleKobayashi, Manami, Yuya Kumagai, Yohei Yamamoto, Hajime Yasui, and Hideki Kishimura. 2020. "Identification of a Key Enzyme for the Hydrolysis of β-(1→3)-Xylosyl Linkage in Red Alga Dulse Xylooligosaccharide from Bifidobacterium Adolescentis" Marine Drugs 18, no. 3: 174. https://doi.org/10.3390/md18030174