Antioxidant, Hypolipidemic and Hepatic Protective Activities of Polysaccharides from Phascolosoma esculenta


 and
and
Abstract
:1. Introduction
2. Results
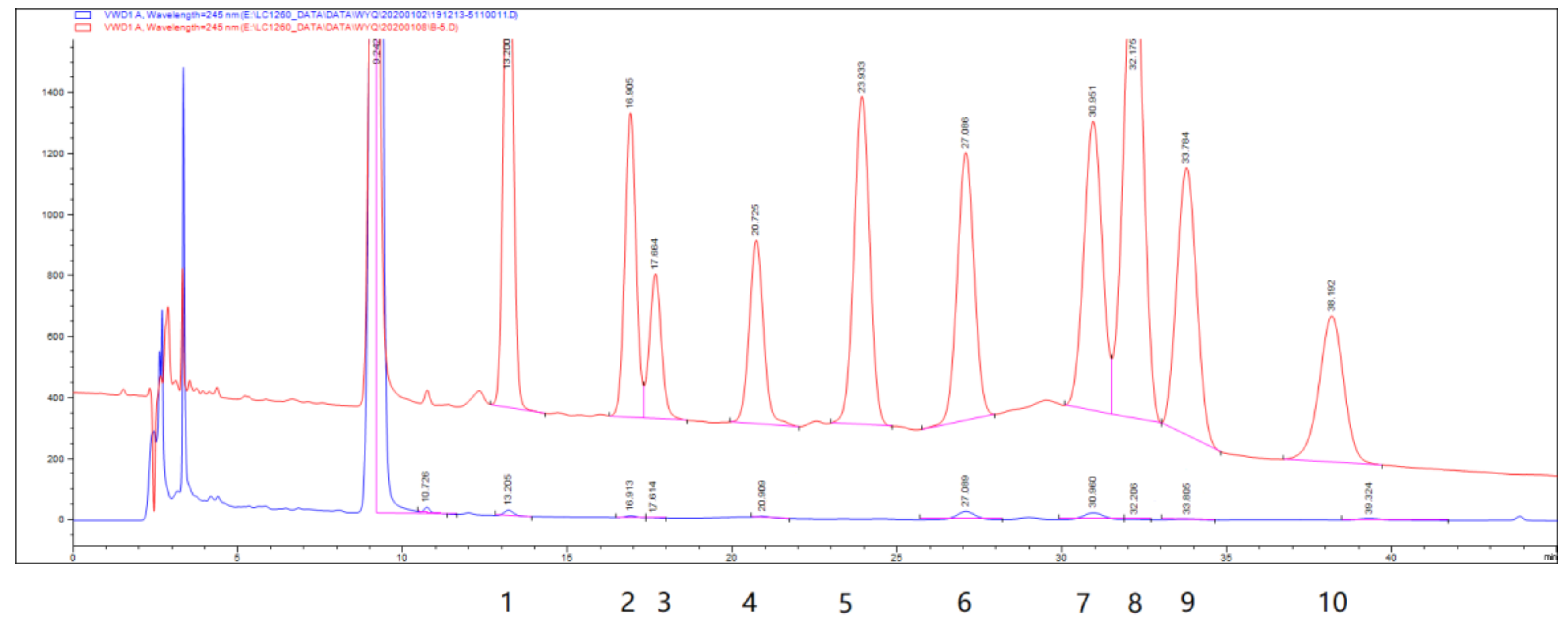
2.1. Characterization of PEP
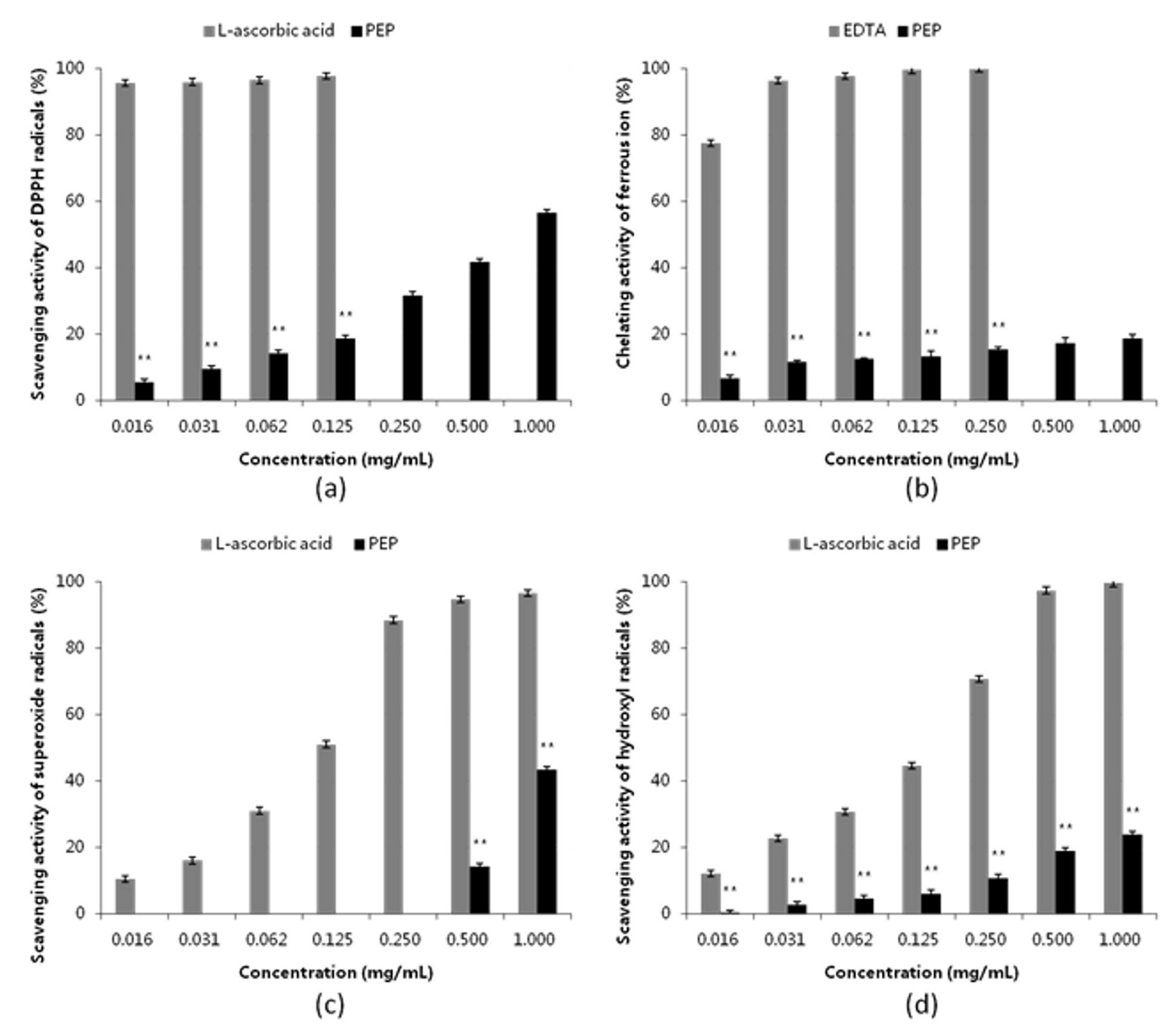
2.2. Antioxidant Activity of PEP in vitro
2.3. Effect of PEP on High-Fat Diet Fed Mice
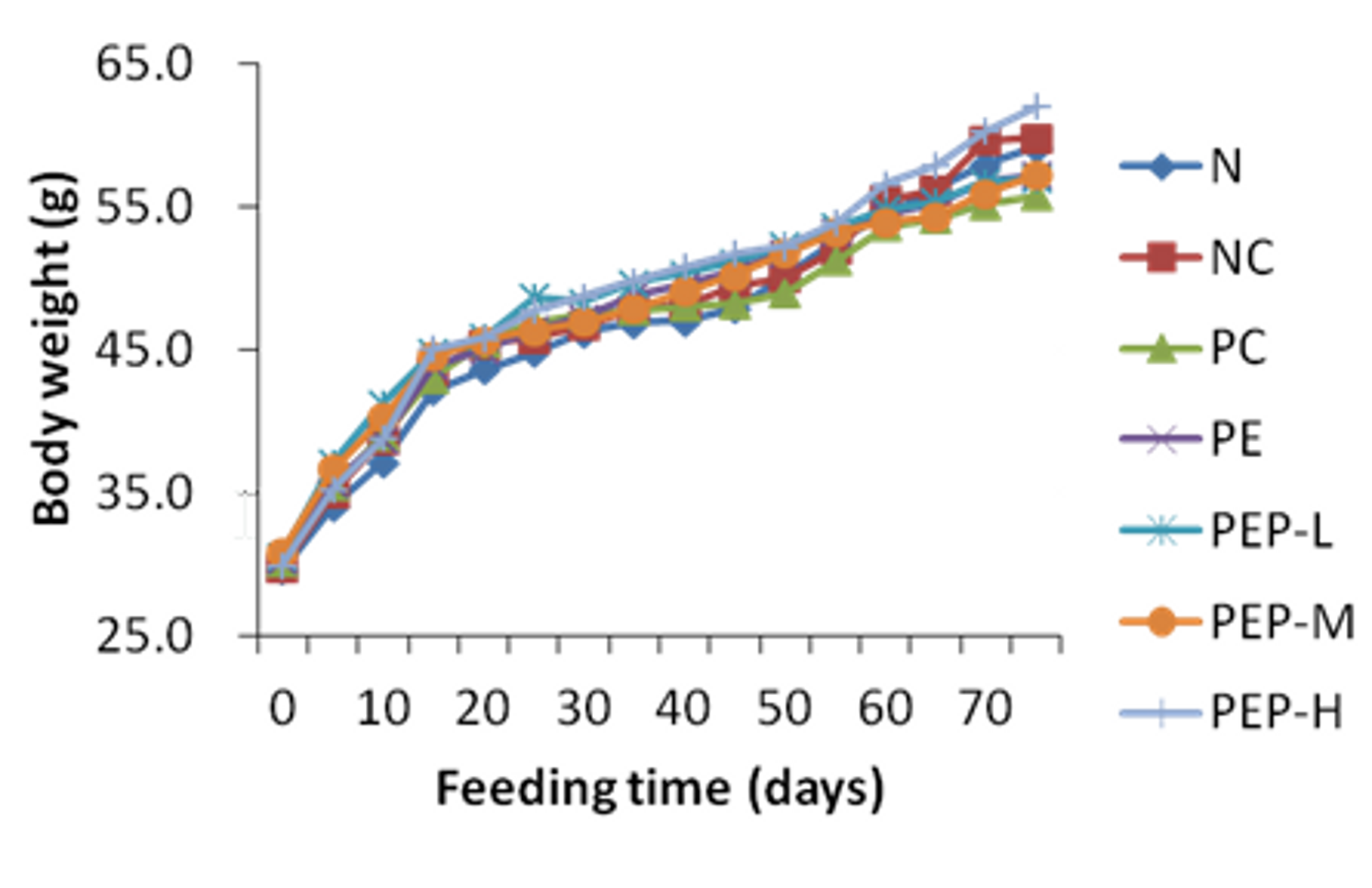
2.3.1. Effect of PEP on Body Weight
2.3.2. Effect of PEP on Serum TC, TG, LDL-C and HDL-C Levels
2.3.3. Effect of PEP on Liver Index and Liver Lipids
2.3.4. Effect of PEP on Levels of Aspartate Aminotransferase (AST) and Alanine Transaminase (ALT) Activity in Serum and Liver
2.3.5. Effect of PEP on Antioxidant Activity of Serum and Liver
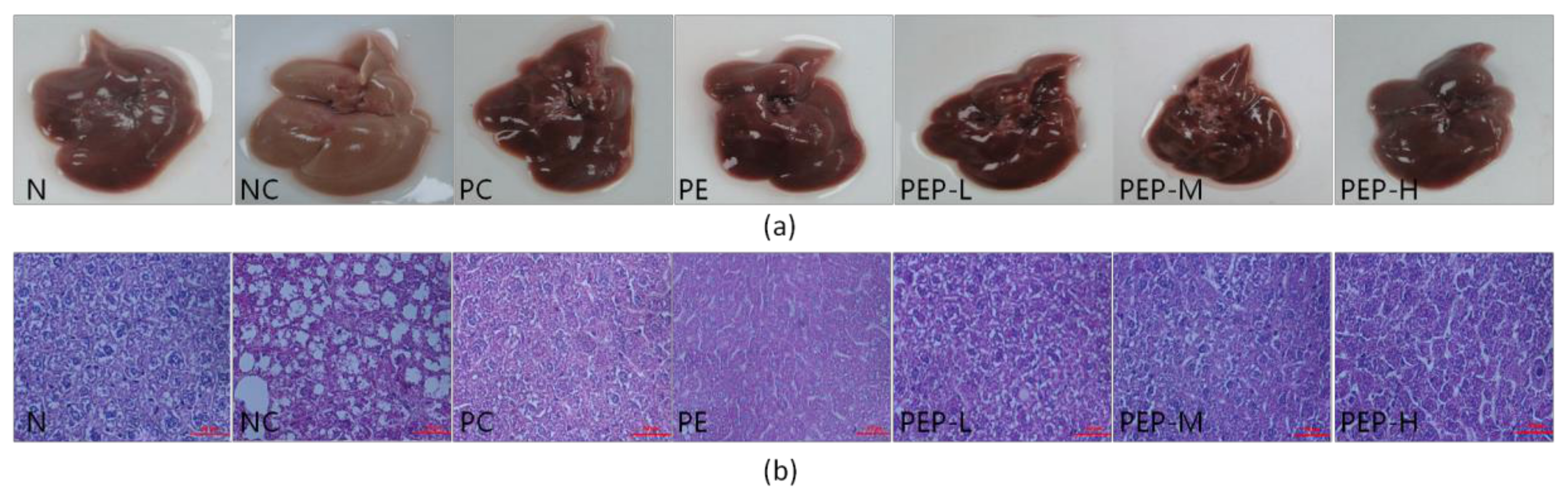
2.3.6. Changes in Morphology and Histopathology of Liver
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of Polysaccharides
4.3. Determination of in vitro Antioxidant Activities of PEP
4.3.1. DPPH Free Radical Scavenging Activity
4.3.2. Ferrous Ion Chelating Activity
4.3.3. Superoxide Anion Radical Scavenging Activity
4.3.4. Hydroxyl Radical Scavenging Activity
4.3.5. Determination of Total Phenolic Contents
4.4. Animals and Experimental Design
4.4.1. Animals and Diets
4.4.2. Biochemical Analysis of Serum
4.4.3. Biochemical Analysis of Liver
4.4.4. Hepatic Histology Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Surhio, M.M.; Wang, Y.; Xu, P.; Shah, F.; Li, J.; Ye, M. Antihyperlipidemic and hepatoprotective properties of selenium modified polysaccharide from Lachnum sp. Int. J. Boil. Macromol. 2017, 99, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, N.; Zhang, J.; Zhao, H.; Lin, L.; Jia, S.; Jia, L. Antihyperlipidemic and hepatoprotective activities of residue polysaccharide from Cordyceps militaris SU-12. Carbohydr. Polym. 2015, 131, 355–362. [Google Scholar] [CrossRef]
- Liu, X.; Sun, Z.; Zhang, M.; Meng, X.; Xia, X.; Yuan, W.; Xue, F.; Liu, C. Antioxidant and antihyperlipidemic activities of polysaccharides from sea cucumber Apostichopus japonicus. Carbohydr. Polym. 2012, 90, 1664–1670. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, M.J.; Laffin, L.J.; Davidson, M.H. Overcoming toxicity and side-effects of lipid-lowering therapies. Best Pr. Res. Clin. Endocrinol. Metab. 2014, 28, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhai, G.; Zhang, J.; Wang, L.; Ma, Z.; Jia, M.; Jia, L. Antihyperlipidemic and hepatoprotective activities of mycelia zinc polysaccharide from Pholiota nameko SW-02. Int. J. Boil. Macromol. 2014, 70, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Wei, B.; Wang, S.; Ke, S.; Chen, J.; Zhang, H.; Wang, H. The Antioxidant Activity of Polysaccharides Derived from Marine Organisms: An Overview. Mar. Drugs 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, R.M.; de Morais, A.M.; de Morais, R.M. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Manlusoc, J.; Hsieh, C.L.; Hsieh, C.Y.; Salac, E.; Lee, Y.T.; Tsai, P.W. Pharmacologic Application Potentials of Sulfated Polysaccharide from Marine Algae. Polymers 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Gao, H.; Wang, S.; Wen, S.; Qin, S. Hypolipidemic and antioxidant properties of a polysaccharide fraction from Enteromorpha prolifera. Int. J. Boil. Macromol. 2013, 58, 186–189. [Google Scholar] [CrossRef]
- Li, W.; Jiang, N.; Li, B.; Wan, M.; Chang, X.; Liu, H.; Zhang, L.; Yin, S.; Qi, H.; Liu, S. Antioxidant activity of purified ulvan in hyperlipidemic mice. Int. J. Boil. Macromol. 2018, 113, 971–975. [Google Scholar] [CrossRef]
- Qi, H.; Ji, X.; Liu, S.; Feng, D.; Dong, X.; He, B.; Srinivas, J.; Yu, C. Antioxidant and anti-dyslipidemic effects of polysaccharidic extract from sea cucumber processing liquor. Electron. J. Biotechnol. 2017, 28, 1–6. [Google Scholar] [CrossRef]
- Liu, H.H.; Ko, W.C.; Hu, M.L. Hypolipidemic effect of glycosaminoglycans from the sea cucumber Metriatyla scabra in rats fed a cholesterol-supplemented diet. J. Agric. Food Chem. 2002, 50, 3602–3606. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, C.; Su, X.; Wang, M.; Li, Y.; Li, Y.; Li, T. Cloning and characterization of hemerythrin gene from Sipuncula Phascolosoma esculenta. Genes Genom. 2013, 35, 95–100. [Google Scholar] [CrossRef]
- Wu, Y.; Fang, M.; Du, L.; Wu, H.; Liu, Y.; Guo, M.; Xie, J.; Wei, D. The nutritional composition and anti-hypertensive activity on spontaneously hypertensive rats of sipuncula Phascolosoma esculenta. Food Funct. 2014, 5, 2317–2323. [Google Scholar] [CrossRef]
- Du, L.; Fang, M.; Wu, H.; Xie, J.; Wu, Y.; Li, P.; Zhang, D.; Huang, Z.; Xia, Y.; Zhou, L.; et al. A novel angiotensin I-converting enzyme inhibitory peptide from Phascolosoma esculenta water-soluble protein hydrolysate. J. Funct. Foods 2013, 5, 475–483. [Google Scholar] [CrossRef]
- Liang, R. Orthogonal test design for optimization of the extraction of polysaccharides from Phascolosoma esulenta and evaluation of its immunity activity. Carbohydr. Polym. 2008, 73, 558–563. [Google Scholar] [CrossRef]
- Niu, R.; Tang, J. Research of anti-fatigue effect of Phascolosoma esculenta. Sci. Technol. Food Ind. 2012, 33, 389–391. [Google Scholar]
- Zhou, Y.; Yan, X.; Li, G.; Ma, Q.; Chen, H.; Zhu, P. Preparation of peptides from Phascolosoma esulenta and the anti-free radical activities of the peptides in physically trained rats. Chin. J. Sports Med. 2008, 27, 55–60. [Google Scholar]
- Xu, R.A.; Xiong, Y.; Cui, X. A marine Phascolosoma esculenta fibrinolytic enzyme and its preparation and application. Chinese patent 201310169730.X, 2013. [Google Scholar]
- Liu, L.; Cao, J.; Chen, J.; Zhang, X.; Wu, Z.; Xiang, H. Effects of peptides from Phascolosoma esculenta on spatial learning and memory via anti-oxidative character in mice. Neurosci. Lett. 2016, 631, 30–35. [Google Scholar] [CrossRef]
- Yu, Z.; Ni, X.; Bian, R. The mammotrophic action of a mixture from Phascolosoma and Penaeus on mice. Chin. J. Mar. Drugs 1992, 4, 4–7. [Google Scholar]
- Guo, M.; Chen, X.; Wu, Y.; Zhang, L.; Huang, W.; Yuan, Y.; Fang, M.; Xie, J.; Wei, D. Angiotensin I-converting enzyme inhibitory peptides from Sipuncula (Phascolosoma esculenta): Purification, identification, molecular docking and antihypertensive effects on spontaneously hypertensive rats. Process. Biochem. 2017, 63, 84–95. [Google Scholar] [CrossRef]
- Zhou, Y.; Ding, L.; Xu, J.; Li, G.; Yan, X. Analysis of main nutritional components in Phascolosoma esculenta. Acta Nutr. Sinica 2007, 29, 413–414. [Google Scholar]
- Wu, Y.; Jiang, H.; Liu, J.; Ma, Q.; Liu, J.; Xu, R.A. Advances of chemical components and pharmacological properties of Sipuncula. Chin. J. Mar. Drugs 2015, 34, 86–92. [Google Scholar]
- Kong, S.; Liu, L.; Lu, X.; Shi, Y.; Chen, J. Optimization of extraction process by response surface methodology of anti-oxidant polysaccharides from Phasolosma esculenta. Sci. Technol. Food Ind. 2014, 35, 233–237. [Google Scholar]
- Yang, Z.; Wang, J.; Li, J.; Xiong, L.; Chen, H.; Liu, X.; Wang, N.; Ouyang, K.; Wang, W. Antihyperlipidemic and hepatoprotective activities of polysaccharide fraction from Cyclocarya paliurus in high-fat emulsion-induced hyperlipidaemic mice. Carbohydr. Polym. 2018, 183, 11–20. [Google Scholar] [CrossRef]
- Choi, W.; Chang, S.; Kim, J.; Lee, S. Hypolipidemic effects of scoparone and its coumarin analogues in hyperlipidemia rats induced by high fat diet. J. Korean Soc. Appl. Boil. Chem. 2013, 56, 647–653. [Google Scholar] [CrossRef]
- Dong, Y.; Qi, Y.; Liu, M.; Song, X.; Zhang, C.; Jiao, X.; Wang, W.; Zhang, J.; Jia, L. Antioxidant, anti-hyperlipidemia and hepatic protection of enzyme-assisted Morehella esculenta polysaccharide. Int. J. Boil. Macromol. 2018, 120, 1490–1499. [Google Scholar] [CrossRef]
- Jiang, C.; Xiong, Q.; Gan, D.; Jiao, Y.; Liu, J.; Ma, L.; Zeng, X. Antioxidant activity and potential hepatoprotective effect of polysaccharides from Cyclina sinensis. Carbohydr. Polym. 2013, 91, 262–268. [Google Scholar] [CrossRef]
- Zeng, H.; Chen, P.; Chang, Q.; Zheng, B.; Zhang, Y. Hypolipidemic effect of polysaccharides from Fortunella margarita (Lour.) Swingle in hyperlipidemic rats. Food Chem. Toxicol. 2019, 132, 110663. [Google Scholar] [CrossRef]
- Wang, X.; Li, W.; Xiao, L.; Liu, C.; Qi, H.; Zhang, Z. In vivo antihyperlipidemic and antioxidant activity of porphyran in hyperlipidemic mice. Carbohydr. Polym. 2017, 174, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Shen, X.; Liu, Y.; Zhang, J.; He, Y.; Liu, Q.; Jiang, D.; Zong, J.; Li, J.; Hou, D.; et al. Isolation, characterization, and radiation protection of Sipunculus nudus L. polysaccharide. Int. J. Boil. Macromol. 2016, 83, 288–296. [Google Scholar] [CrossRef]
- Li, B.; Xu, H.; Wang, X.; Wan, Y.; Jiang, N.; Qi, H.; Liu, X. Antioxidant and antihyperlipidemic activities of high sulfate content purified polysaccharide from Ulva pertusa. Int. J. Boil. Macromol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Huang, H.; Yang, N.; Qin, M.; Shi, Z. Study on optimization of enzymolysis-alkali echelon extraction of polysaccharides from Phscolosoma esculenta, its antioxidant and antimicrobial activity. J. South. Agric. 2019, 50, 1085–1092. [Google Scholar]
- Barker, M.F.; Xu, R.A. Population differences in gonad and pyloric caeca cycles of the New Zealand seastar Sclerasterias mollis (Echinodermata: Asteroidea). Mar. Boil. 1991, 108, 97–103. [Google Scholar] [CrossRef]
- Barker, M.F.; Xu, R.A. Seasonal changes in biochemical composition of body walls, gonads and pyloric caeca in two populations of Sclerasterias mollis (Echinodermata: Asteroidea) during the annual reproductive cycle. Mar. Boil. 1991, 109, 27–34. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, Q.; Tong, T.; Xu, M. Extraction and Antioxidant Activity in Vitro of Polysaccharide from Sipunculus Nudus Coelomic Fluid. Food Res. Dev. 2015, 36, 46–49. [Google Scholar]
- Teng, Z.; Qian, L.; Zhou, Y. Hypolipidemic activity of the polysaccharides from Enteromorpha prolifera. Int. J. Boil. Macromol. 2013, 62, 254–256. [Google Scholar] [CrossRef]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Vamanu, E.; Gatea, F. Correlations between Microbiota Bioactivity and Bioavailability of Functional Compounds: A Mini-Review. Biomedicines 2020, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B. The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohydr. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Terho, T.T.; Hartiala, K. Method for determination of the sulfate content of glycosaminoglycans. Anal. Biochem. 1971, 41, 471–476. [Google Scholar] [CrossRef]
- Li, J.; Yuan, P.; Wang, X.; Aipire, A.; Li, M.; Yang, J.; Tao, H.; Ying, T.; Fu, C.; Wei, X.; et al. Purification, characterization and bioactivities of polysaccharides from Pleurotus ferulae. Food Funct. 2017, 8, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Akao, E.; Suzuki, S.; Okuda, M.; Kakehi, K.; Nakamura, J. High-performance liquid chromatography of reducing carbohydrates as strongly ultraviolet-absorbing and electrochemically sensitive 1-phenyl-3-methyl5-pyrazolone derivatives. Anal. Biochem. 1989, 180, 351–357. [Google Scholar] [CrossRef]
- Saija, A.; Tomaino, A.; Lo, C.R.; Rapisarda, P.; Dederen, J.C. In vitro antioxidant activity and in vivo photoprotective effect of a red orange extract. Int. J. Cosmet. Sci. 1998, 20, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinis, T.C.; Maderia, V.M.; Almeida, L.M. Action of phenolic derivatives (acetaminophen, salicylate, and 5-aminosalicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Liu, F.; Ng, T.B. Antioxidative and free radical scavenging activities of selected medicinal herbs. Life Sci. 2000, 66, 725–735. [Google Scholar] [CrossRef]
- Guo, T.; Wei, L.; Sun, J.; Hou, C.; Fan, L. Antioxidant activities of extract and fractions from Tuber indicum Cooke & Massee. Food Chem. 2011, 127, 1634–1640. [Google Scholar] [CrossRef]
- Xu, R.A.; Sun, X.; Tse, L.Y.; Li, H.; Chan, P.C.; Xu, S.; Xiao, W.; Kung, H.F.; Krissansen, G.W.; Fan, S.T. Long-term expression of angiostatin suppresses metastatic liver cancer in mice. Hepatology 2003, 37, 1451–1460. [Google Scholar] [CrossRef]
- Shi, J.; Zheng, D.; Liu, Y.; Sham, M.H.; Tam, P.; Farzaneh, F.; Xu, R.A. Overexpression of soluble TRAIL induces apoptosis in human lung adenocarcinoma and inhibits growth of tumor xenografts in nude mice. Cancer Res. 2005, 65, 1687–1692. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | TG (mmol·L−1) | TC (mmol·L−1) | LDL-C (mmol·L−1) | HDL-C (mmol·L−1) | AI |
|---|---|---|---|---|---|
| N | 1.19 ± 0.27 | 4.02 ± 0.66 | 0.27 ± 0.11 | 2.15 ± 0.68 | 1.09 ± 0.91 |
| NC | 2.52 ± 0.66 ## | 7.79 ± 1.70 ## | 1.08 ± 0.45 ## | 0.89 ± 0.23 ## | 8.40 ± 3.15 ## |
| PC | 1.56 ± 0.57 ** | 5.35 ± 0.99 ** | 0.47 ± 0.24 ** | 1.47 ± 0.38 ** | 2.86 ± 1.21 ** |
| PE | 1.53 ± 0.32 ** | 5.40 ± 0.74 ** | 0.45 ± 0.35 ** | 1.38 ± 0.54 * | 3.25 ± 1.14 ** |
| PEP-L | 1.90 ± 0.55 * | 5.45 ± 1.41 ** | 0.47 ± 0.41 ** | 1.31 ± 0.39 * | 3.36 ± 1.27 ** |
| PEP-M | 1.25 ± 0.33 ** | 5.00 ± 1.02** | 0.55 ± 0.15 ** | 1.38 ± 0.45 ** | 2.92 ± 1.35 ** |
| PEP-H | 1.24 ± 0.49 ** | 5.35 ± 1.07 ** | 0.53 ± 0.33 ** | 1.42 ± 0.40 ** | 3.09 ± 1.47 ** |
| Groups | Liver index (%) | TC (mmol·gprot−1) | TG (mmol·gprot−1) |
|---|---|---|---|
| N | 3.63 ± 0.38 | 0.078 ± 0.004 | 0.051 ± 0.006 |
| NC | 4.43 ± 0.20 ## | 0.152 ± 0.008 ## | 0.096 ± 0.007 ## |
| PC | 3.66 ± 0.26 ** | 0.101 ± 0.02 ** | 0.060 ± 0.009 ** |
| PE | 3.75 ± 0.35 ** | 0.119 ± 0.014 ** | 0.066 ± 0.006 * |
| PEP-L | 4.07 ± 0.35 * | 0.109 ± 0.012 ** | 0.070 ± 0.005 |
| PEP-M | 3.70 ± 0.43 ** | 0.108 ± 0.013 ** | 0.061 ± 0.005 ** |
| PEP-H | 3.65 ± 0.50 ** | 0.120 ± 0.015 ** | 0.074 ± 0.009 |
| Group | Serum AST (U·L−1) | Serum ALT (U·L−1) | Liver AST (U·gprot−1) | Liver ALT (U·gprot−1) |
|---|---|---|---|---|
| N | 13.98 ± 0.80 | 4.27 ± 0.73 | 3.16 ± 1.37 | 5.08 ± 0.39 |
| NC | 28.68 ± 1.99 ## | 14.55 ± 1.51 ## | 12.15 ± 1.46 ## | 24.50 ± 1.35 ## |
| PC | 17.08 ± 1.96 ** | 9.71 ± 0.98 ** | 5.36 ± 0.91 ** | 8.92 ± 0.92 ** |
| PE | 20.67 ± 2.03 ** | 8.76 ± 0.94 ** | 6.84 ± 0.85 ** | 15.93 ± 1.23 ** |
| PEP-L | 23.32 ± 2.00 ** | 8.34 ± 1.02 ** | 6.25 ± 0.94 ** | 16.84 ± 1.01 ** |
| PEP-M | 22.66 ± 2.66 ** | 8.08 ± 0.76 ** | 5.10 ± 0.88 ** | 14.15 ± 1.46 ** |
| PEP-H | 20.19 ± 2.10 ** | 7.86 ± 0.92 ** | 7.00 ± 0.97 ** | 15.62 ± 1.07 ** |
| Group | Serum SOD (U·mL−1) | Serum GSH-Px (U·mL−1) | Serum MDA (nmol·mL−1) | Liver SOD (U·gprot−1) | Liver GSH-Px (U·mgprot−1) | Liver MDA (nmol·mgprot−1) |
|---|---|---|---|---|---|---|
| N | 51.95 ± 8.14 | 2798.36 ± 186.85 | 10.37 ± 0.94 | 28.35 ± 4.91 | 371.92 ± 104.69 | 2.65 ± 0.35 |
| NC | 41.77 ± 7.20 ## | 2317.42 ± 330.65 ## | 13.72 ± 1.87## | 19.31 ± 3.78 ## | 268.11 ± 35.81 # | 3.47 ± 0.58 ## |
| PC | 50.14 ± 6.28 * | 2691.38 ± 119.74 ** | 10.96 ± 0.75** | 24.57 ± 4.05 ** | 347.23 ± 84.93 * | 2.90 ± 0.61 * |
| PE | 49.55 ± 5.54 * | 2607.26 ± 209.84 * | 11.29 ± 1.95 * | 24.20 ± 4.15 * | 352.80 ± 99.33 * | 2.91 ± 0.51 * |
| PEP-L | 46.03 ± 5.99 | 2673.23 ± 348.06 * | 11.53 ± 1.49 * | 22.93 ± 4.49 | 345.59 ± 92.85 * | 2.92 ± 0.64 |
| PEP-M | 51.02 ± 8.09 * | 2655.42 ± 381.07 * | 10.37 ± 0.97 ** | 24.50 ± 4.39 * | 349.73 ± 111.13 * | 2.89 ± 0.61 * |
| PEP-H | 50.56 ± 6.50 * | 2594.17 ± 348.20 | 10.57 ± 0.92 ** | 23.97 ± 3.49 * | 339.52 ± 129.96 | 2.95 ± 0.73 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Jiang, H.; Lin, J.-S.; Liu, J.; Wu, C.-J.; Xu, R. Antioxidant, Hypolipidemic and Hepatic Protective Activities of Polysaccharides from Phascolosoma esculenta. Mar. Drugs 2020, 18, 158. https://doi.org/10.3390/md18030158
Wu Y, Jiang H, Lin J-S, Liu J, Wu C-J, Xu R. Antioxidant, Hypolipidemic and Hepatic Protective Activities of Polysaccharides from Phascolosoma esculenta. Marine Drugs. 2020; 18(3):158. https://doi.org/10.3390/md18030158
Chicago/Turabian StyleWu, Yaqing, Hongying Jiang, Jyuan-Siou Lin, Jia Liu, Chang-Jer Wu, and Ruian Xu. 2020. "Antioxidant, Hypolipidemic and Hepatic Protective Activities of Polysaccharides from Phascolosoma esculenta" Marine Drugs 18, no. 3: 158. https://doi.org/10.3390/md18030158