The Biological and Chemical Diversity of Tetramic Acid Compounds from Marine-Derived Microorganisms


{kind=link}
{kind=link}
{kind=link}
{kind=link}
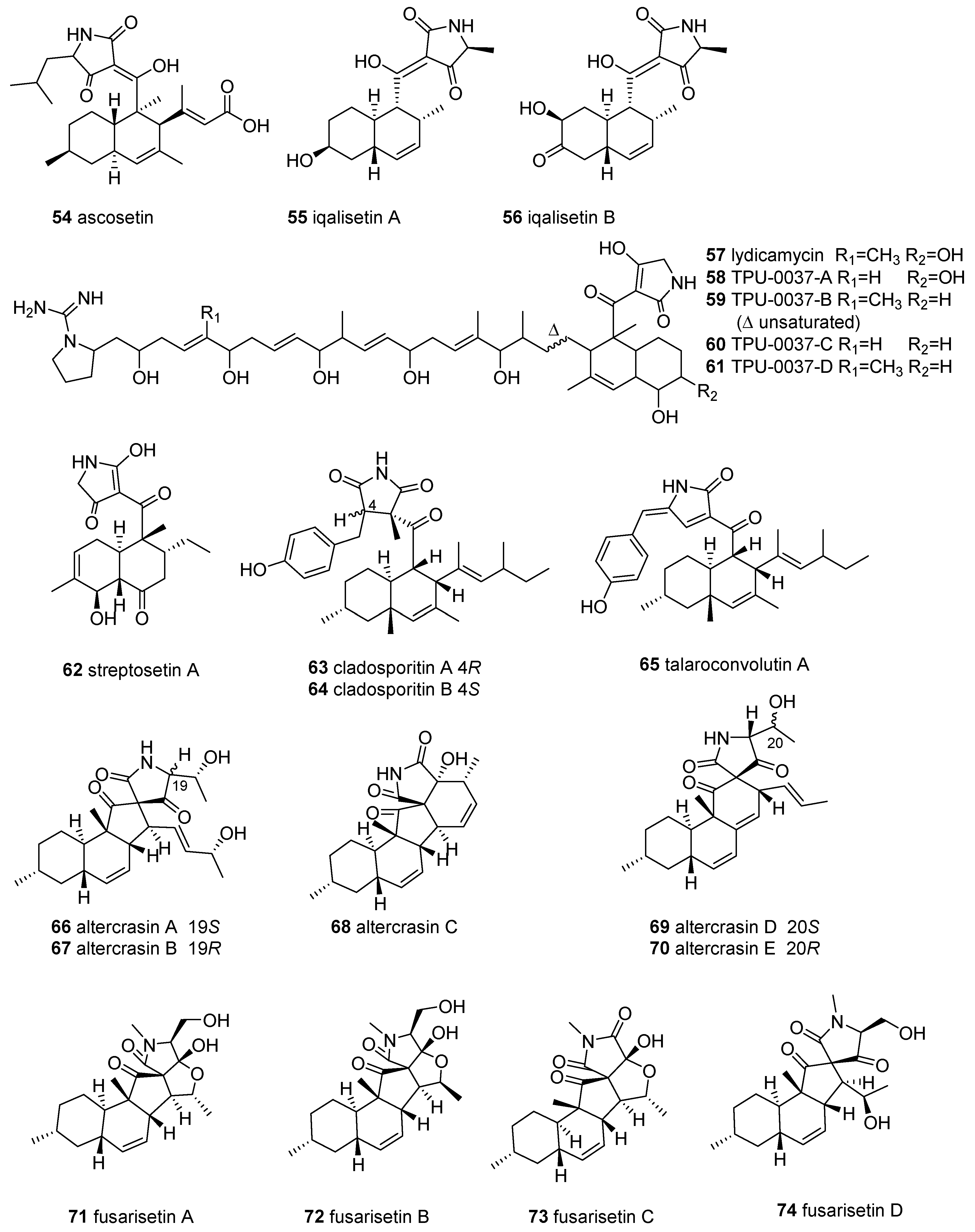{kind=link}
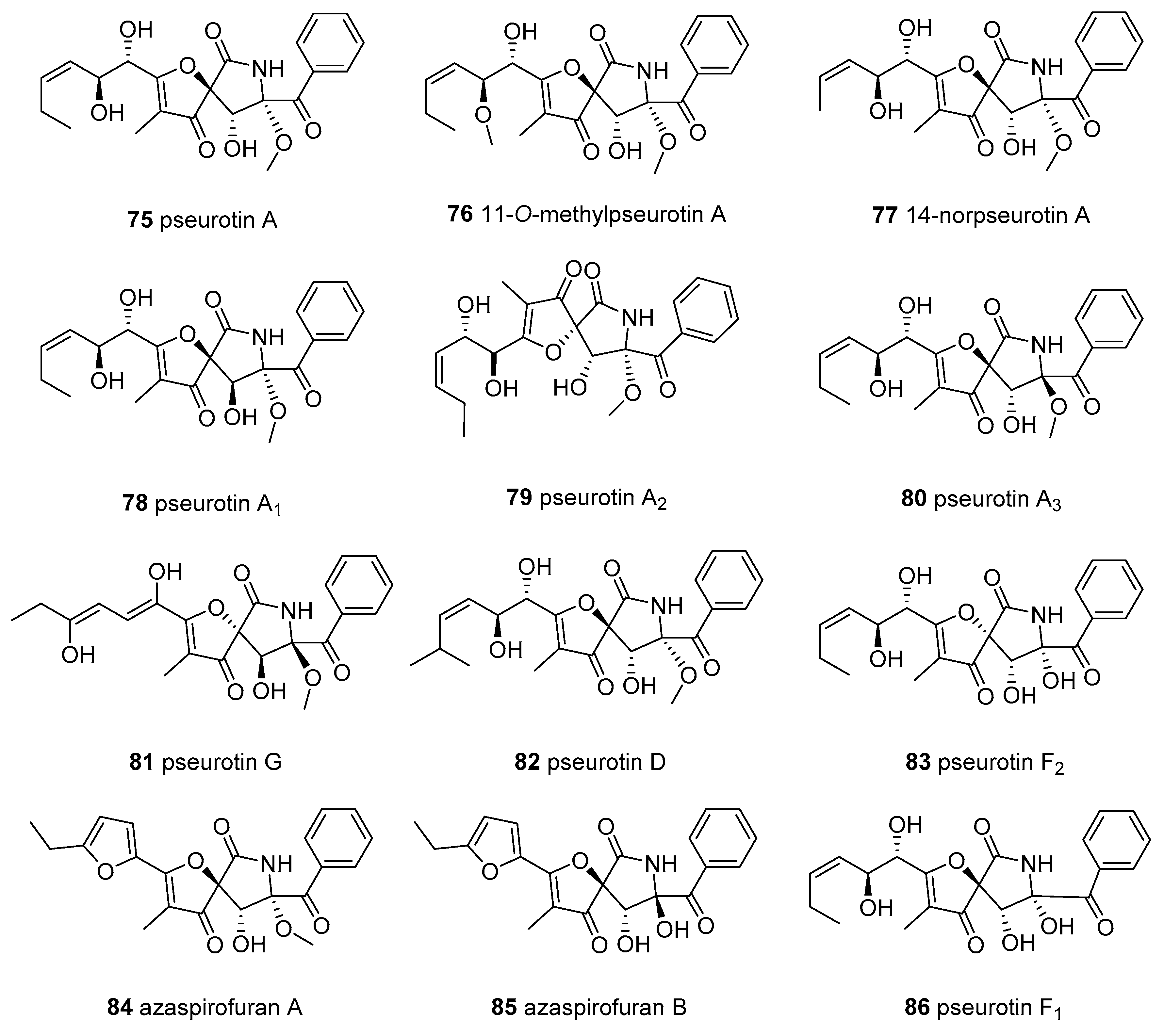{kind=link}
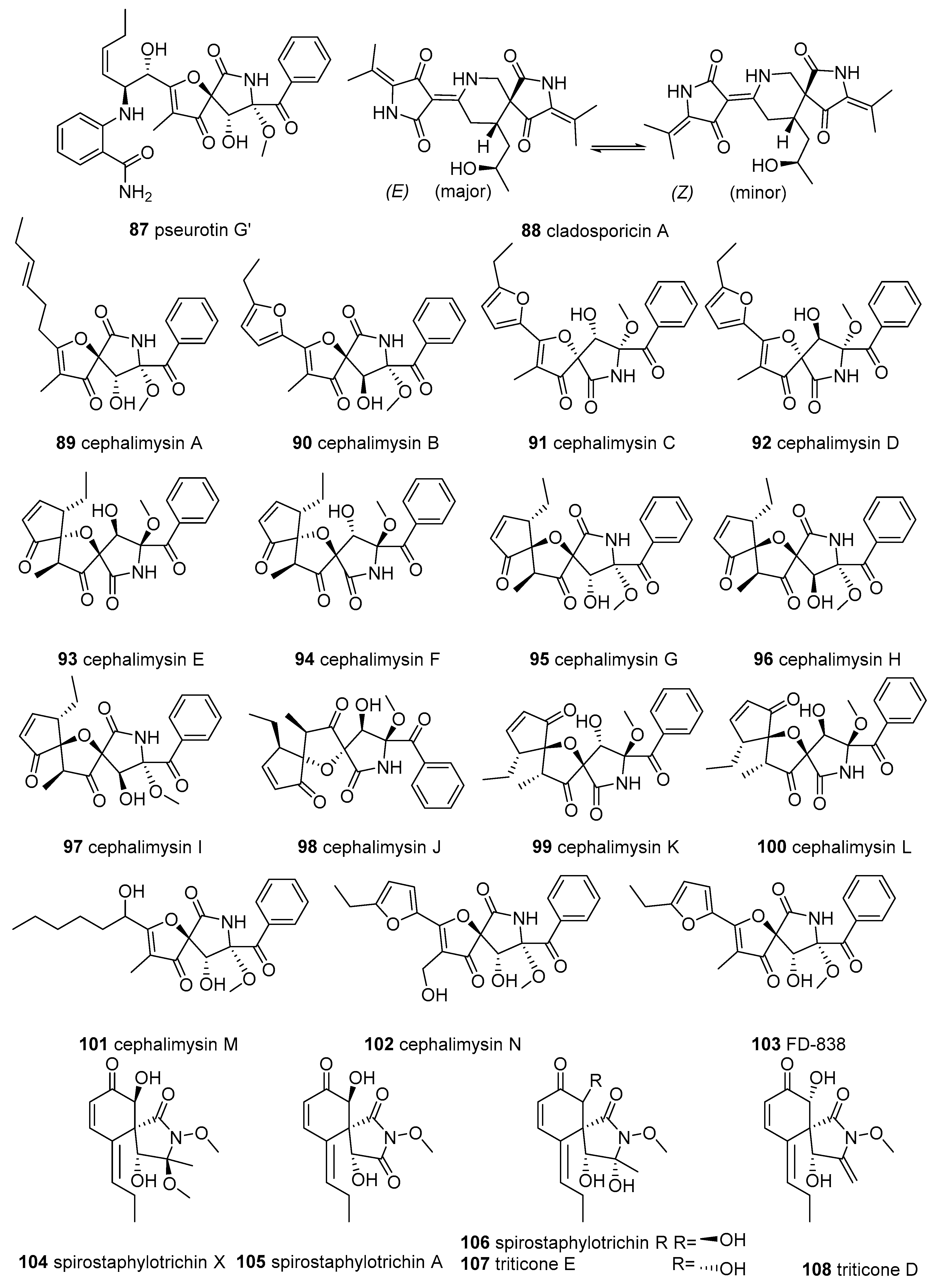{kind=link}
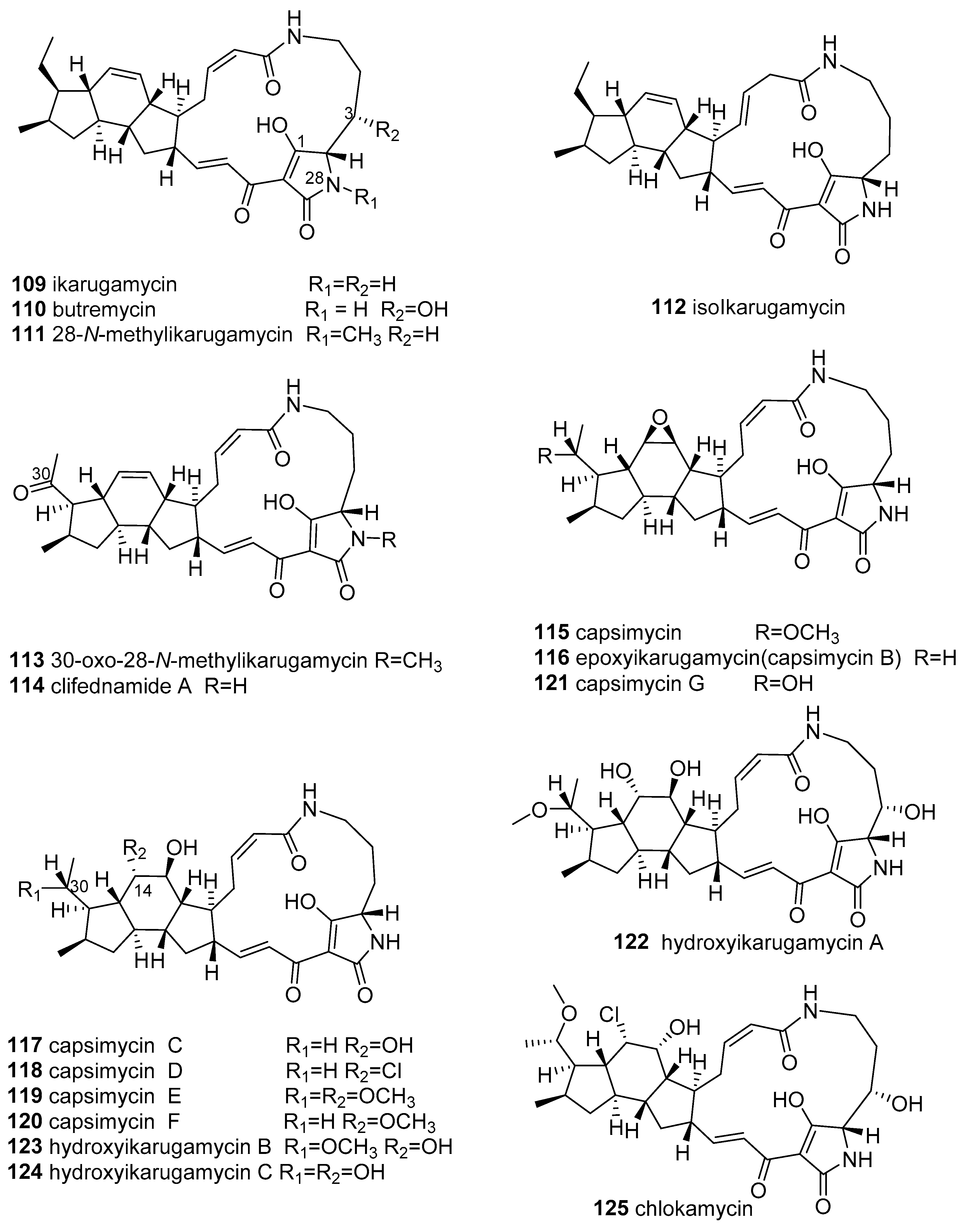{kind=link}
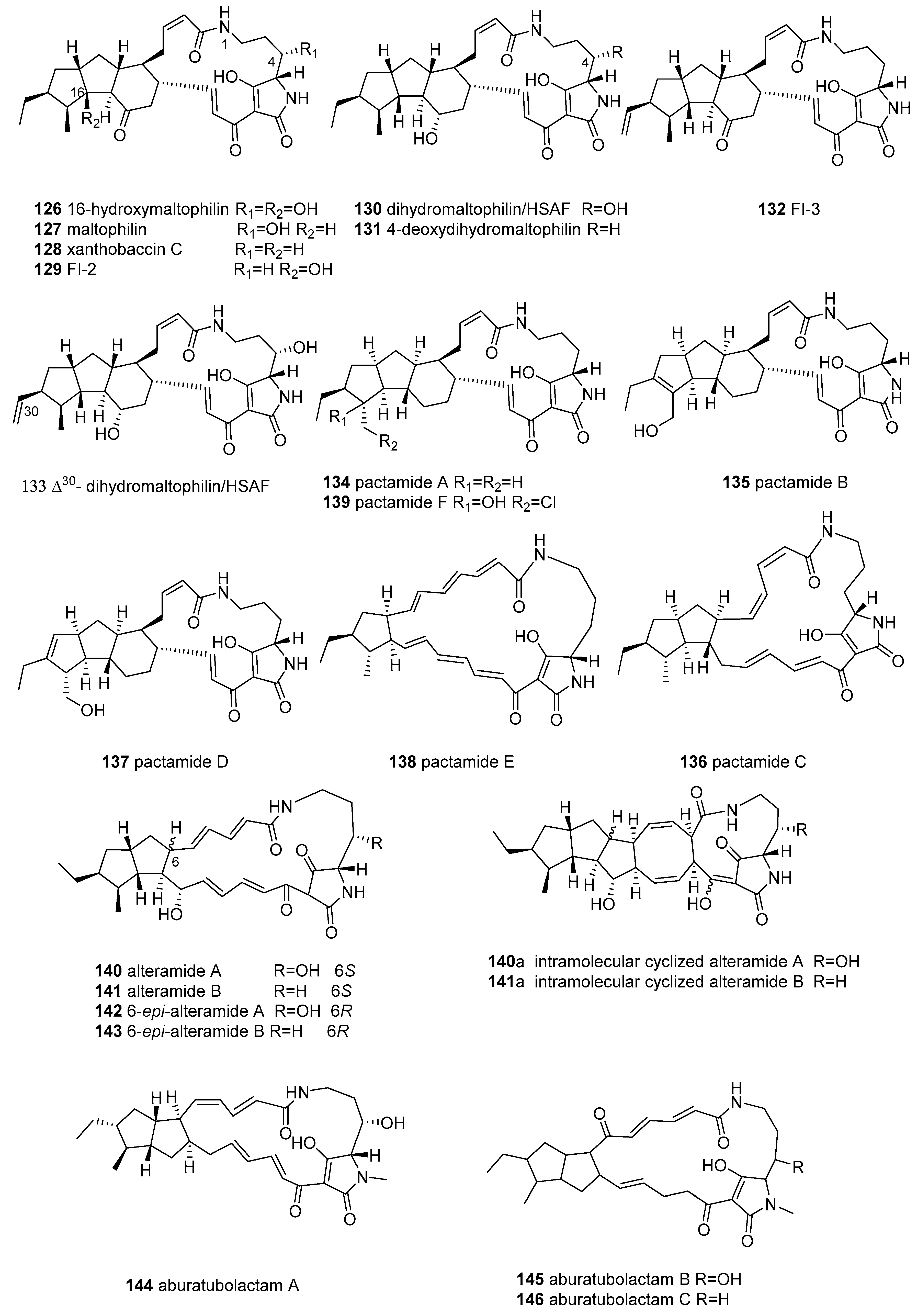{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
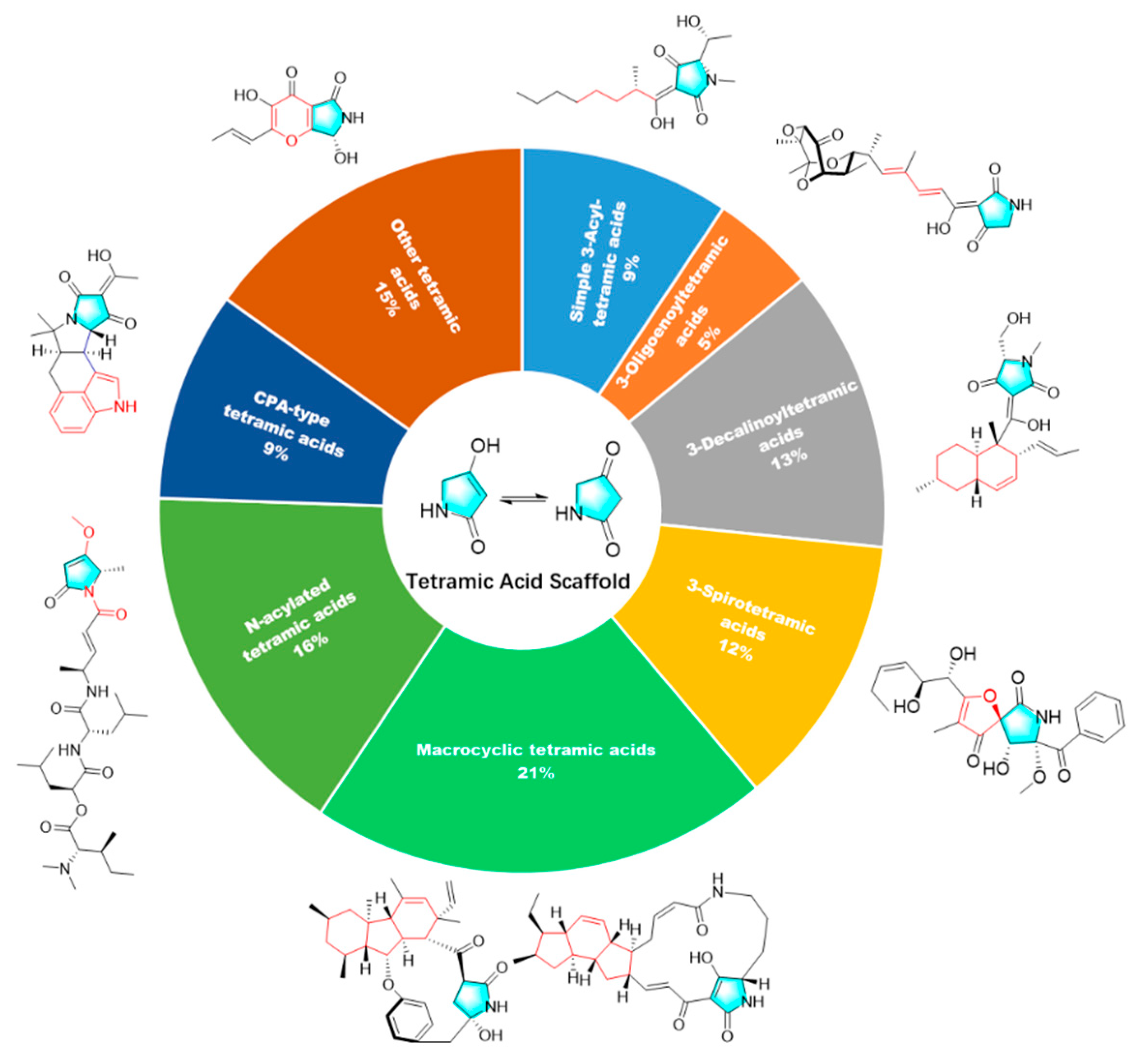
2. Isolation, Structure, and Bioactivities of Tetramic Acid Products from Marine Microbes
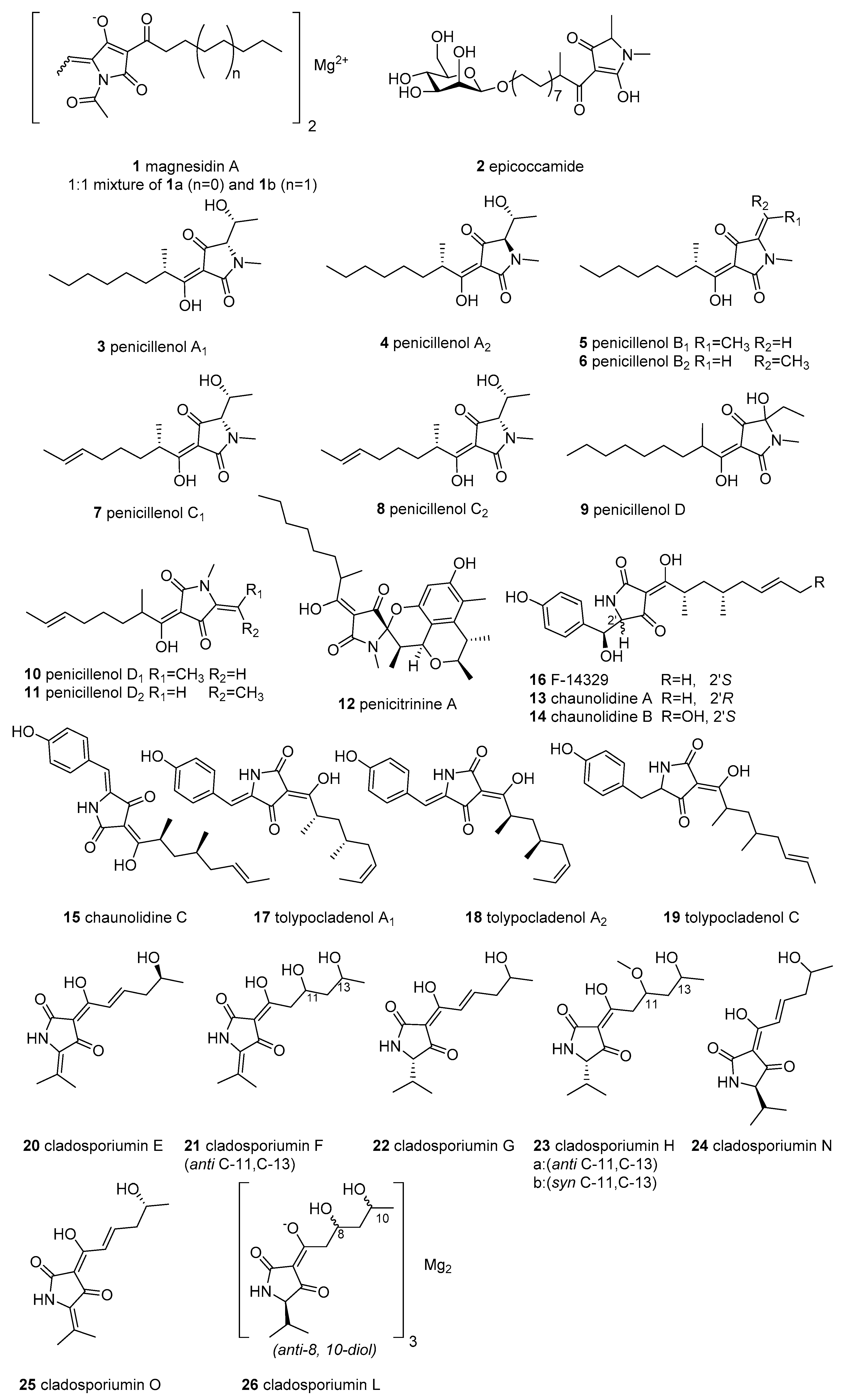
2.1. Simple 3-acyl Tetramic Acid
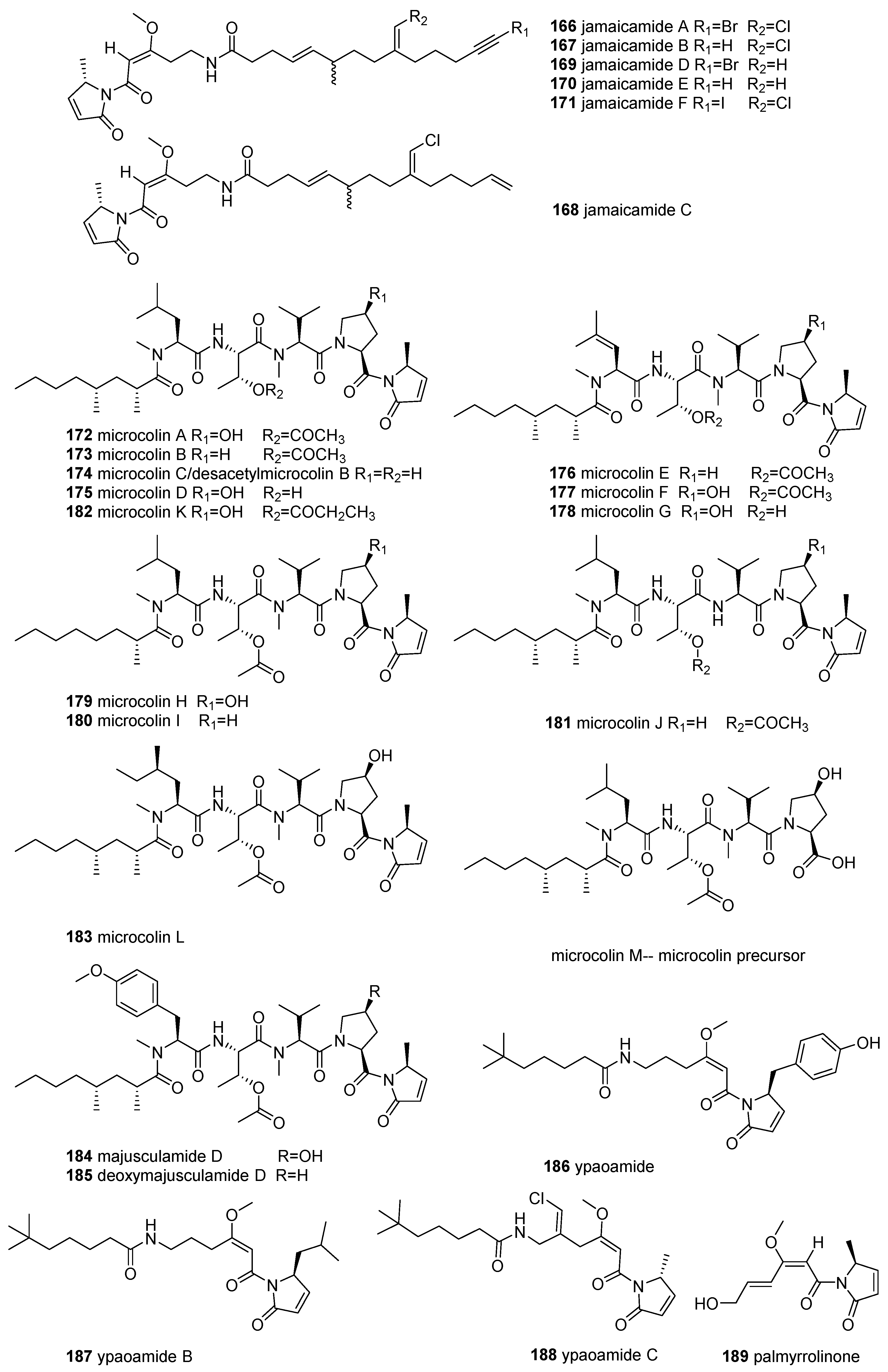
2.2. 3-Oligoenoyltetramic Acids
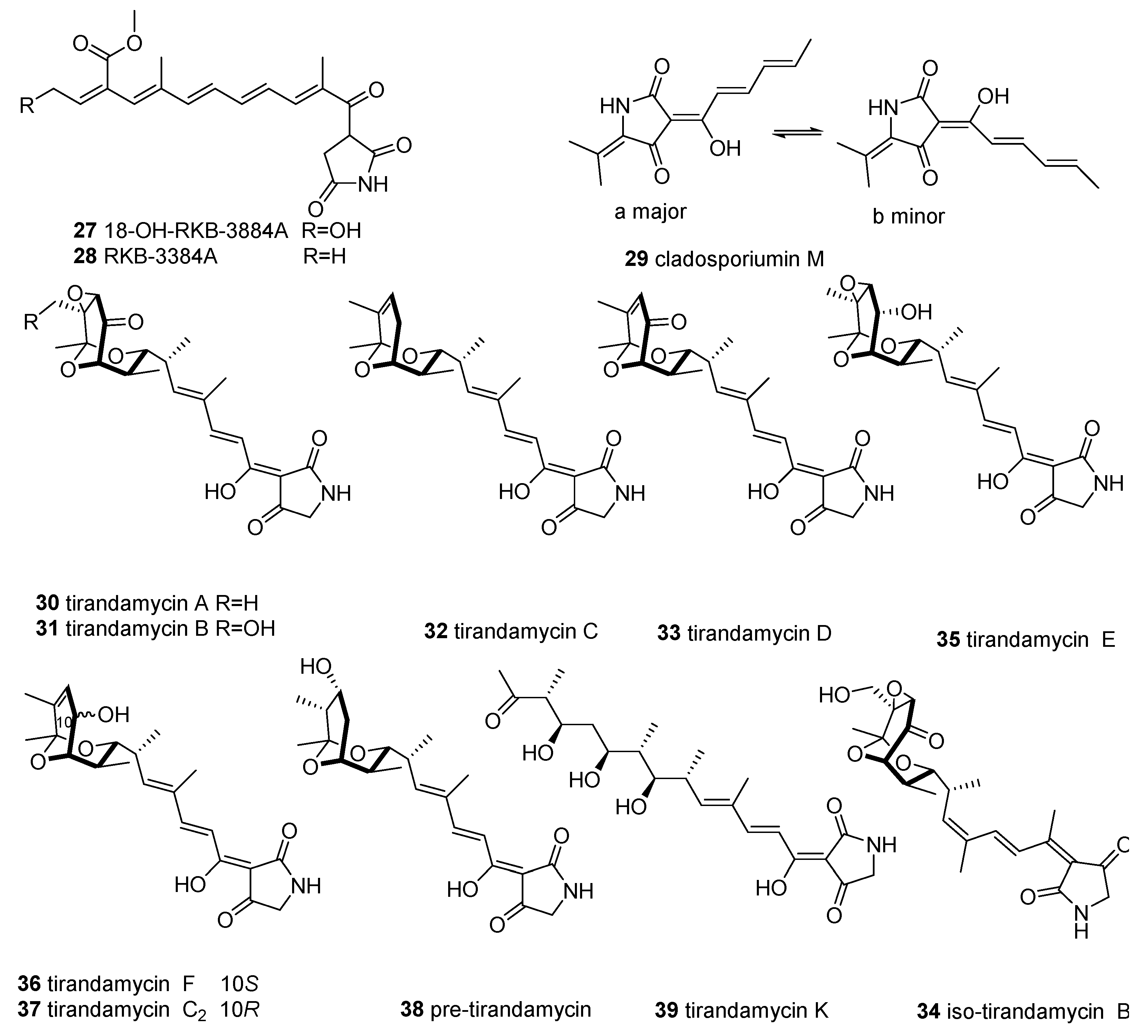
2.3. 3-Decalinoyltetramic Acids
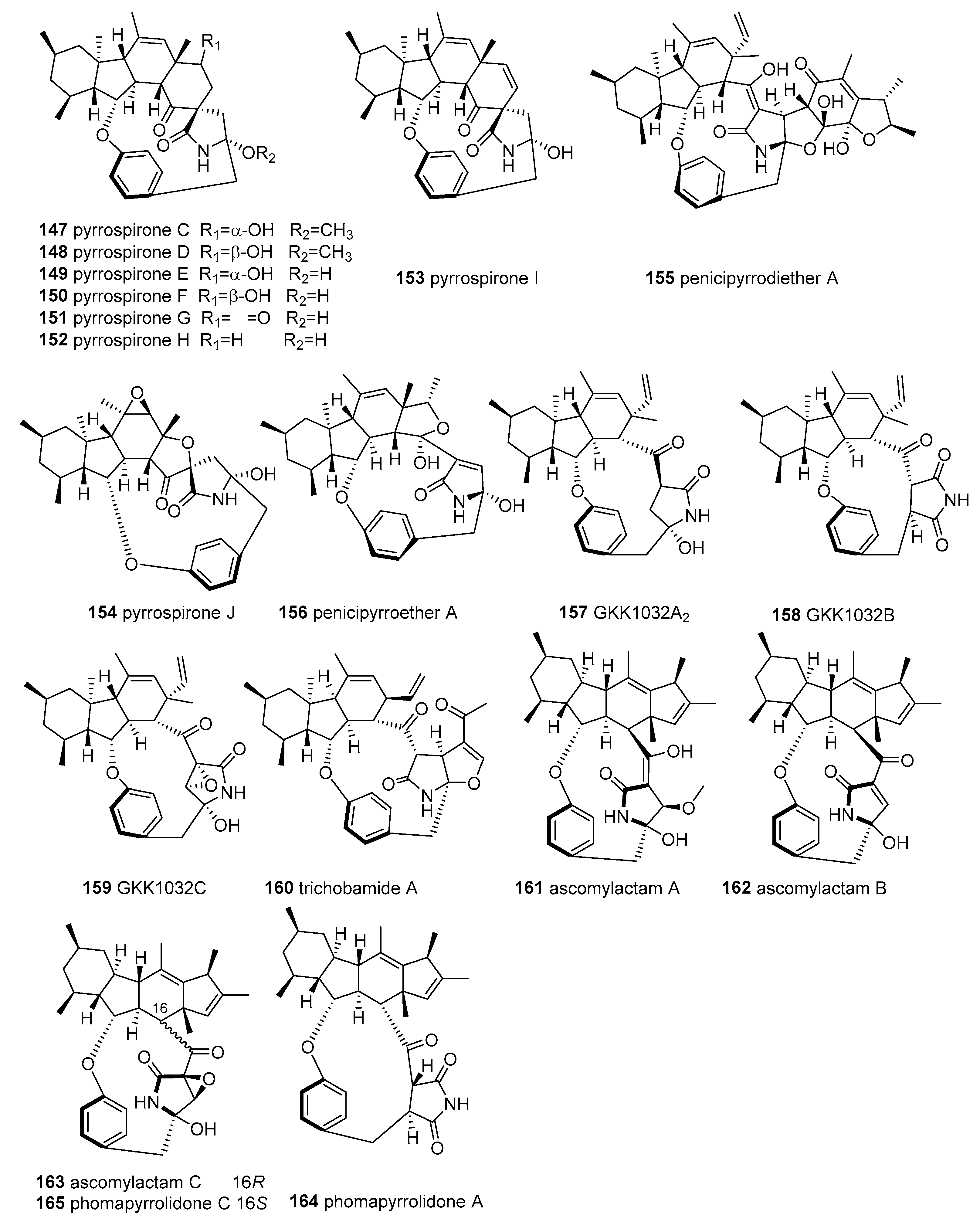
2.4. 3-Spirotetramic Acids
2.5. Macrocyclic Tetramic Acids
2.5.1. Polycyclic Tetramate Macrolactams (PTMs)
2.5.2. Pyrrocidine Tetramate Alkaloids (PTAs)
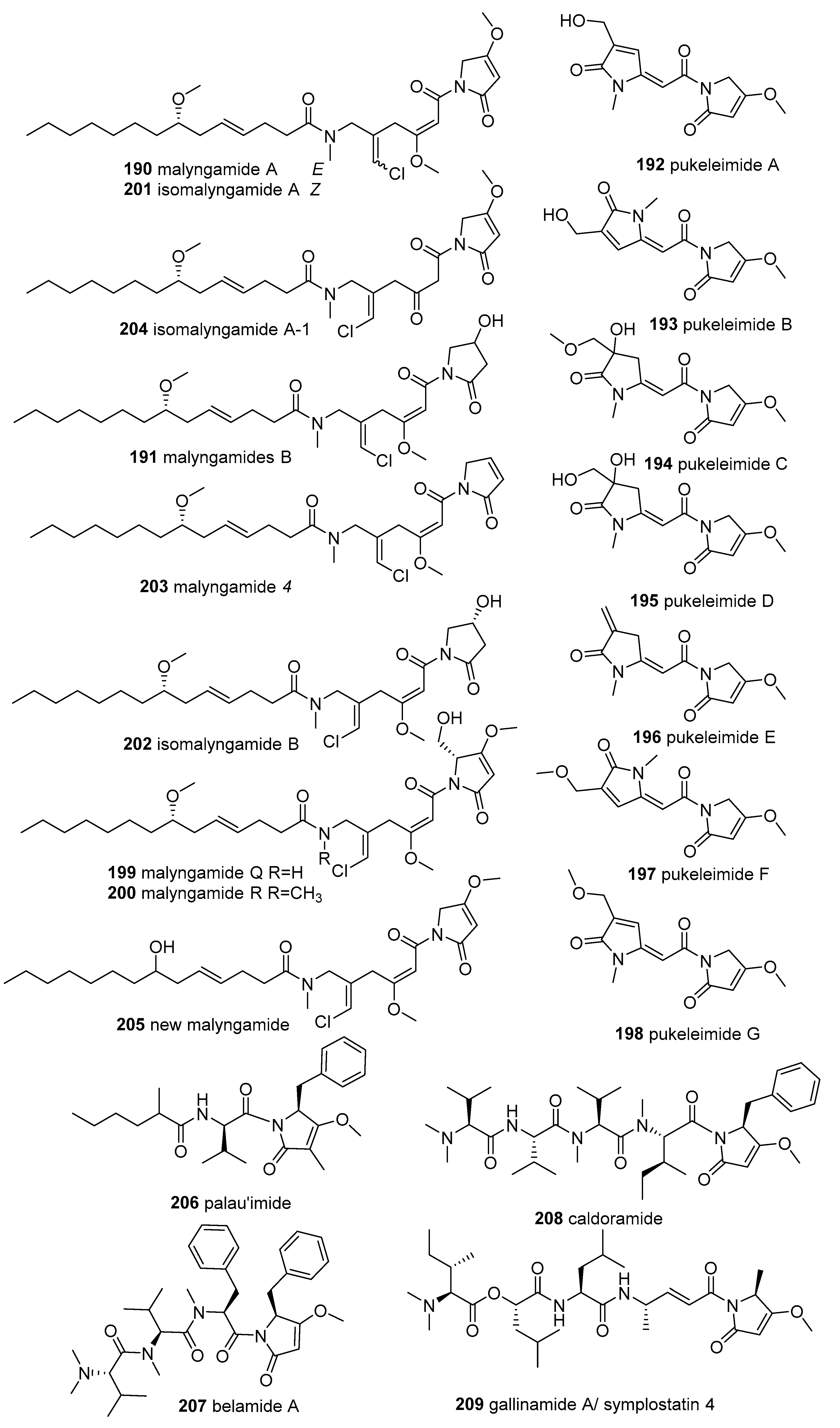
2.6. N-acylated Tetramic Acids
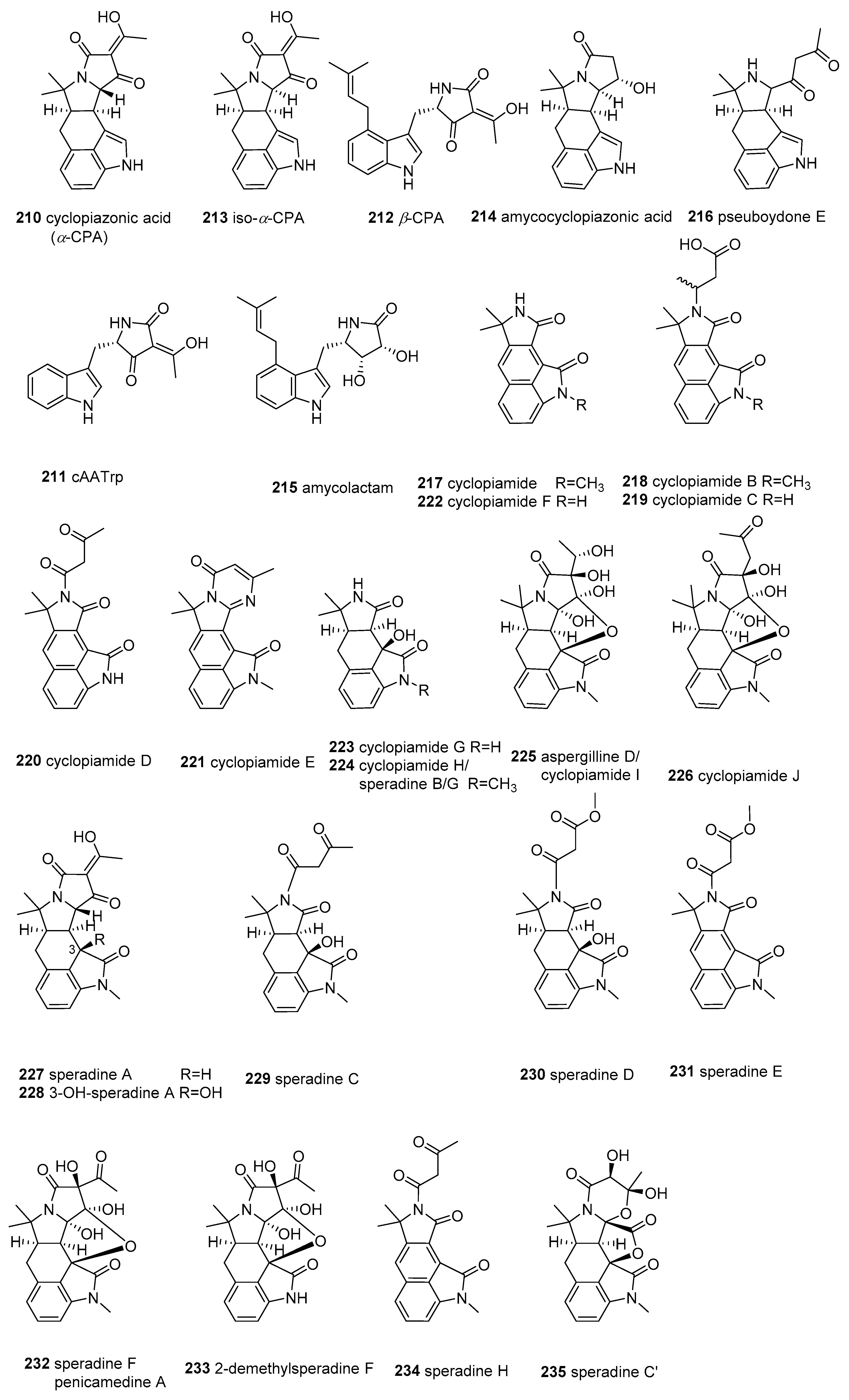
2.7. α-Cyclopiazonic acid (CPA)-type Tetramic Acids
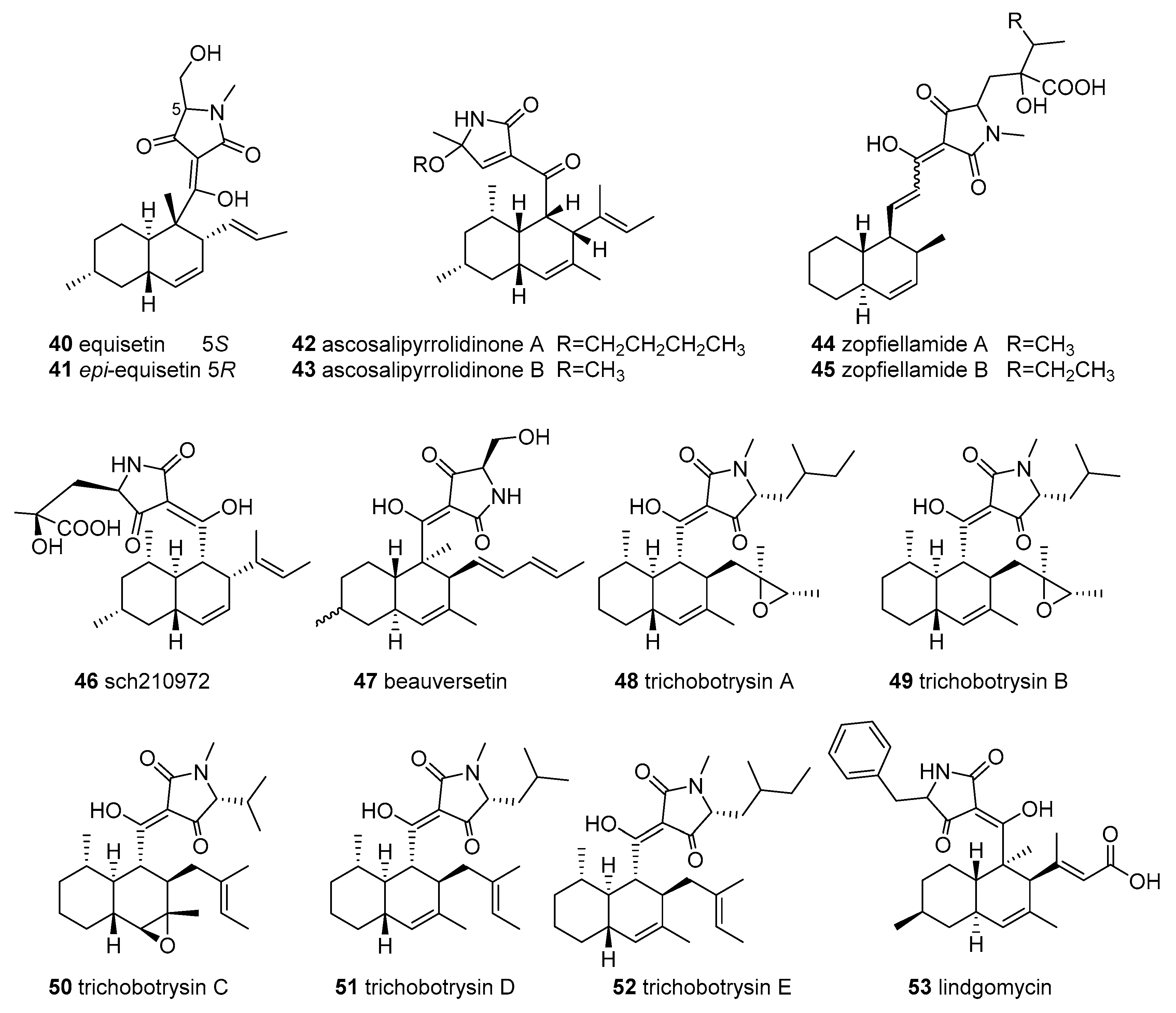
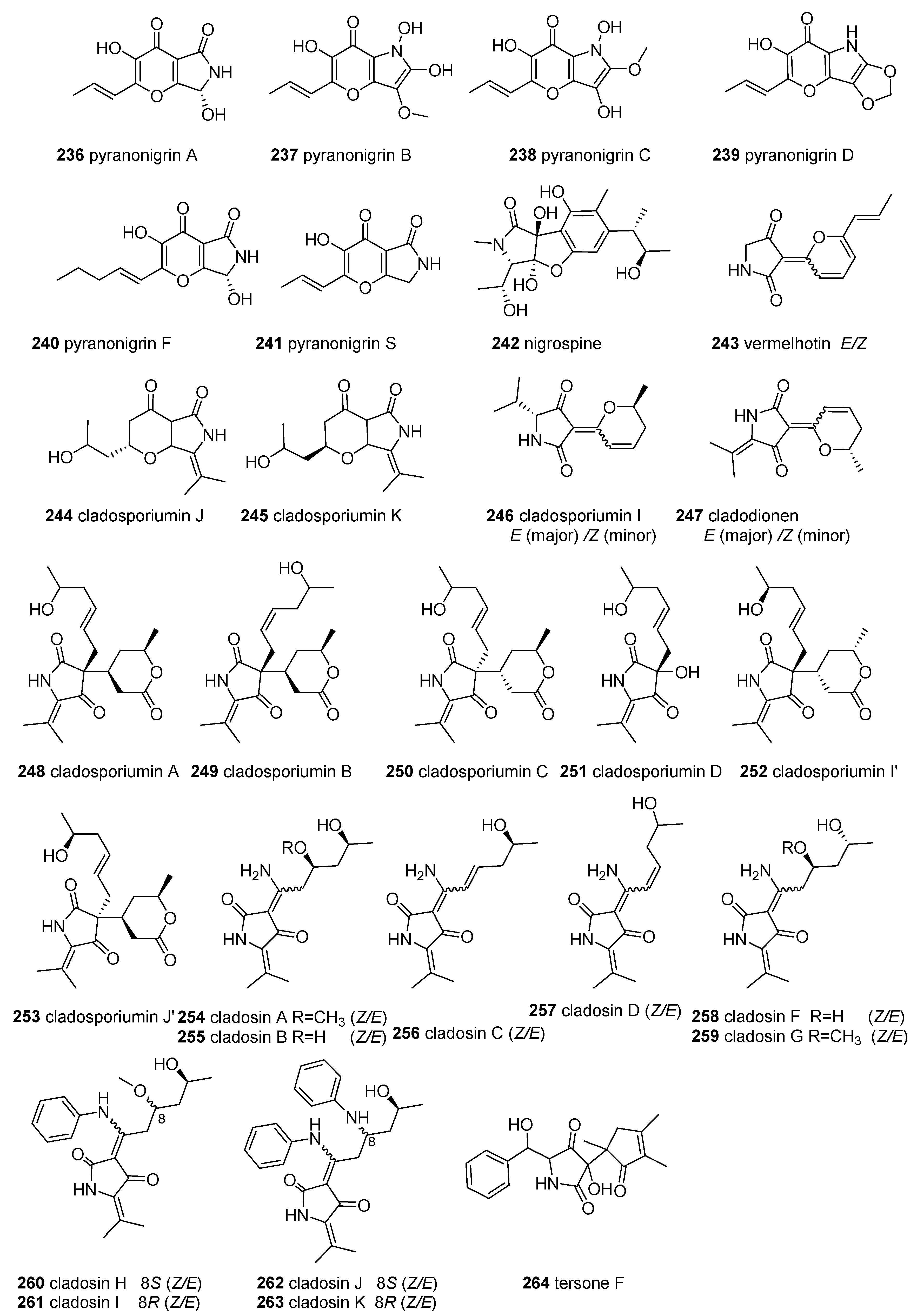
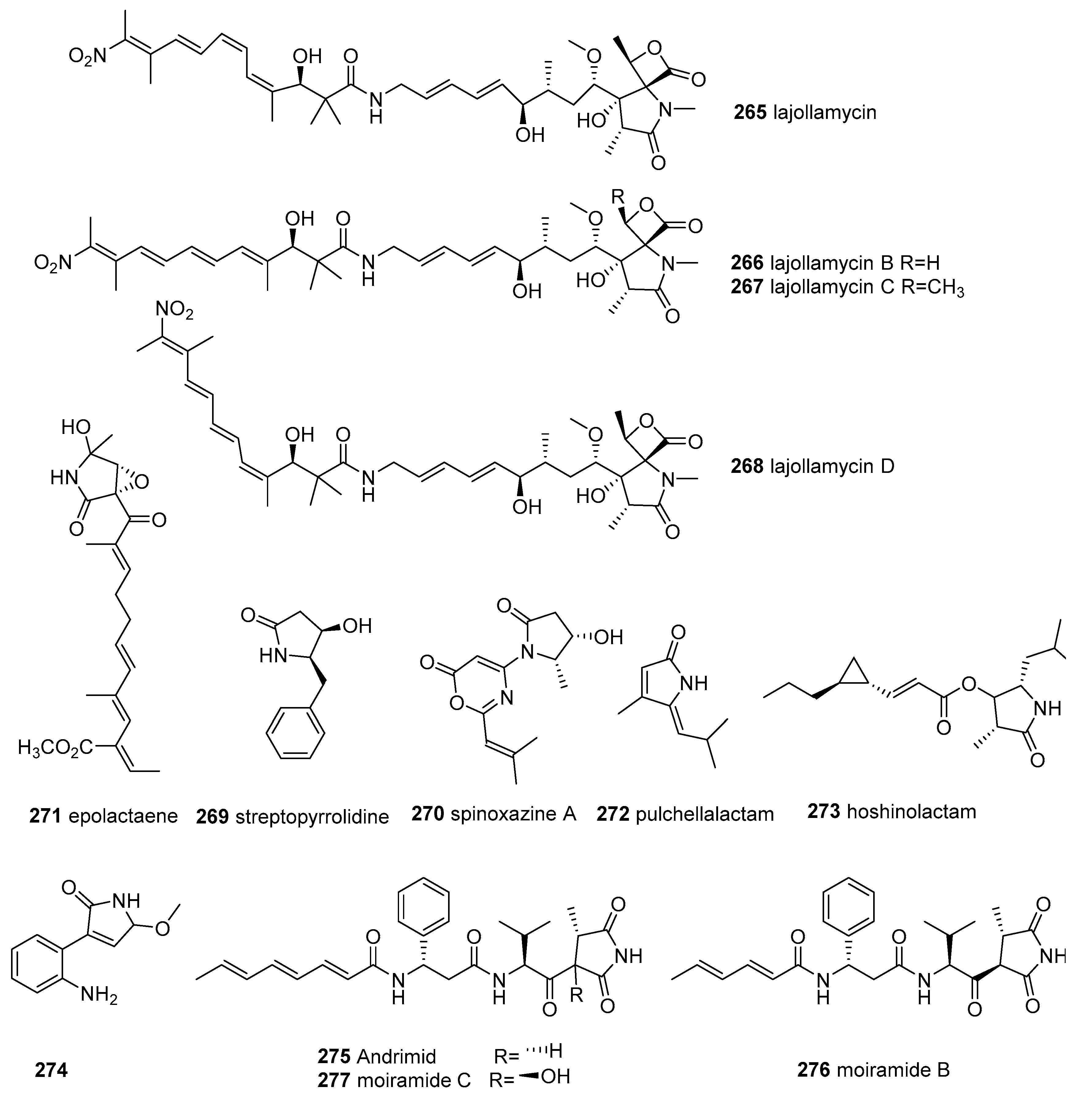
2.8. Other Tetramic Acids
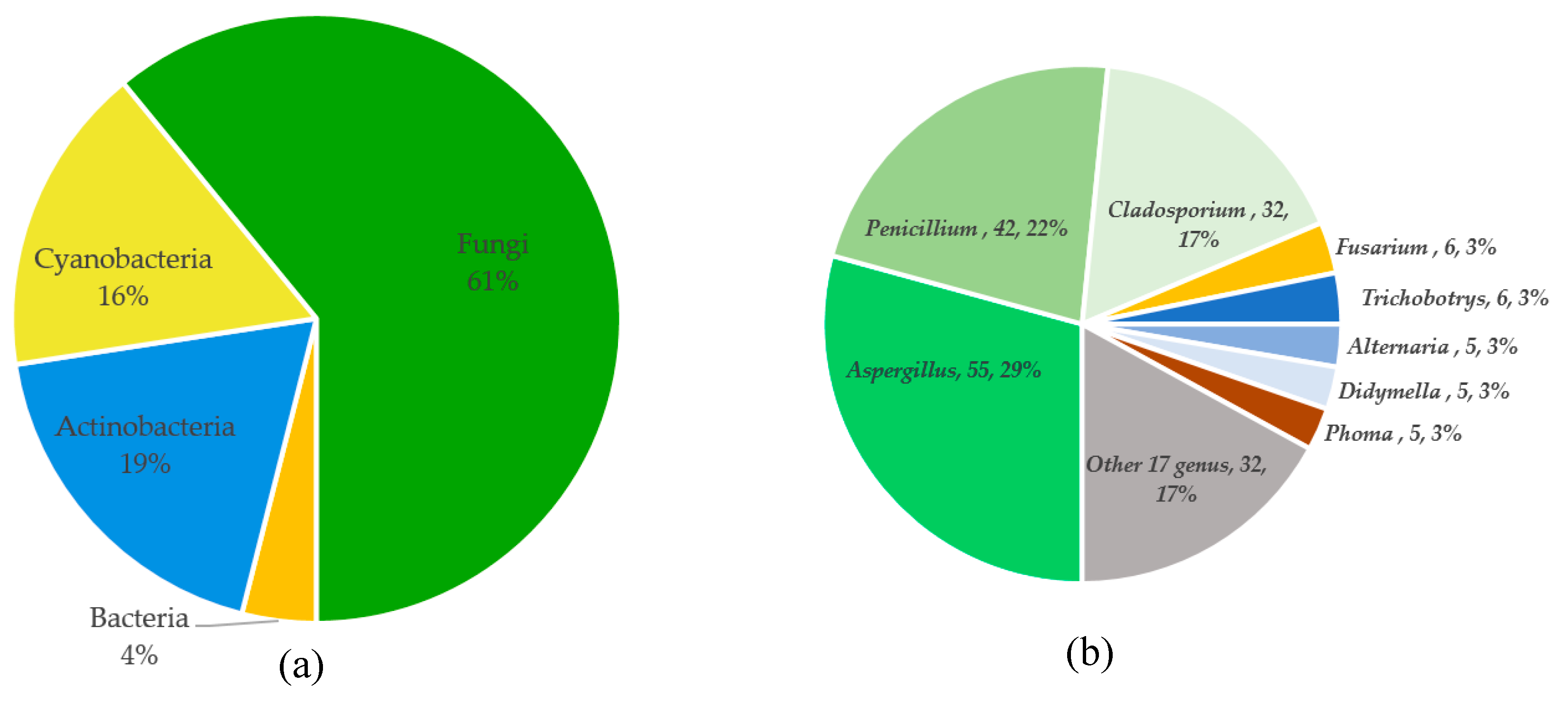
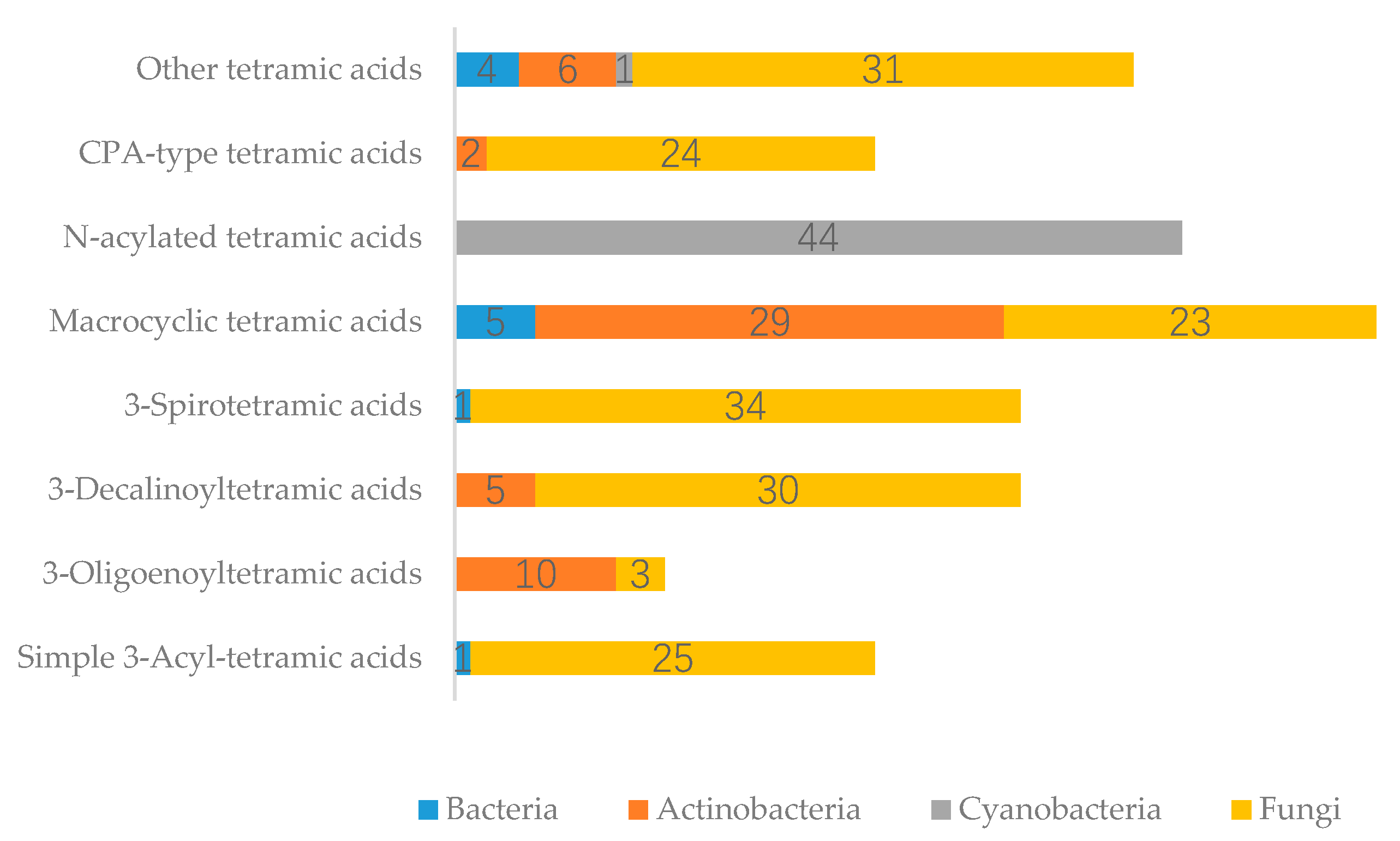
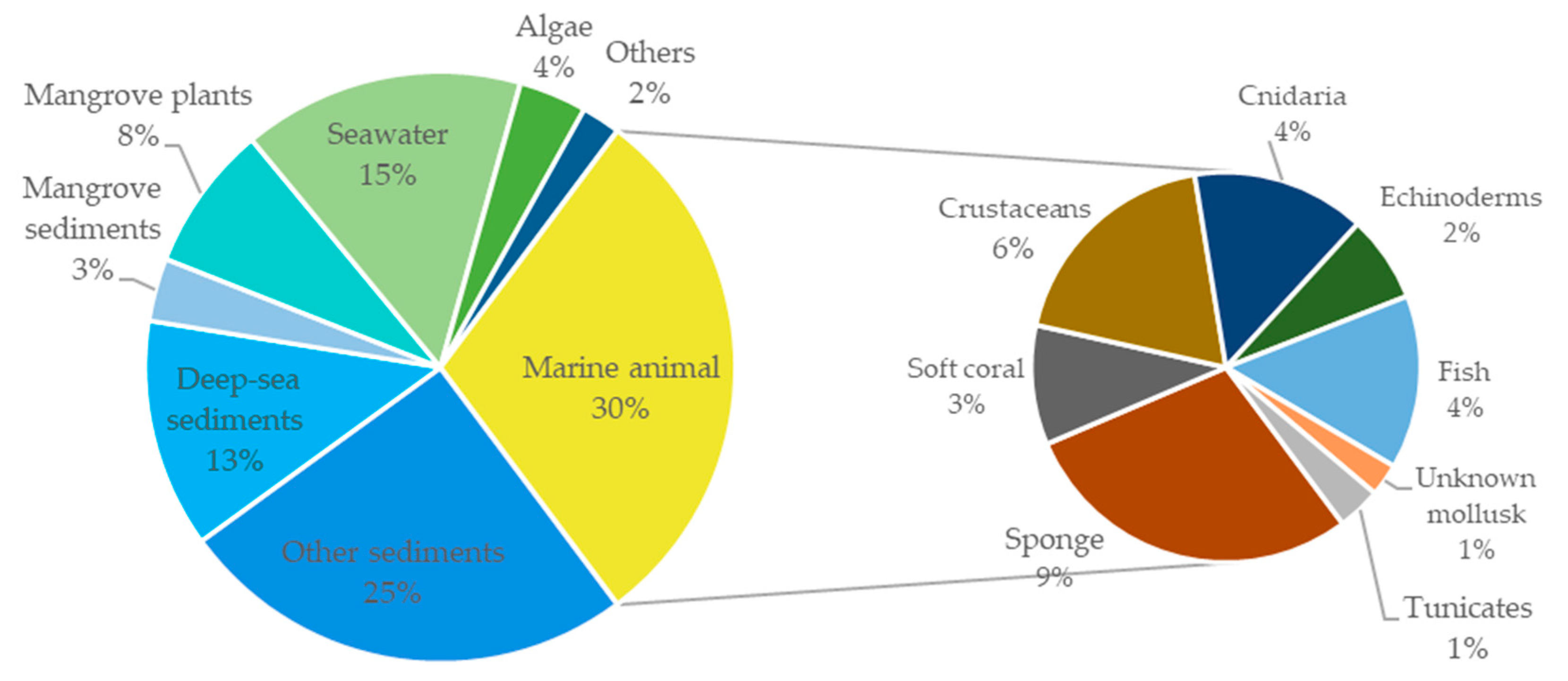
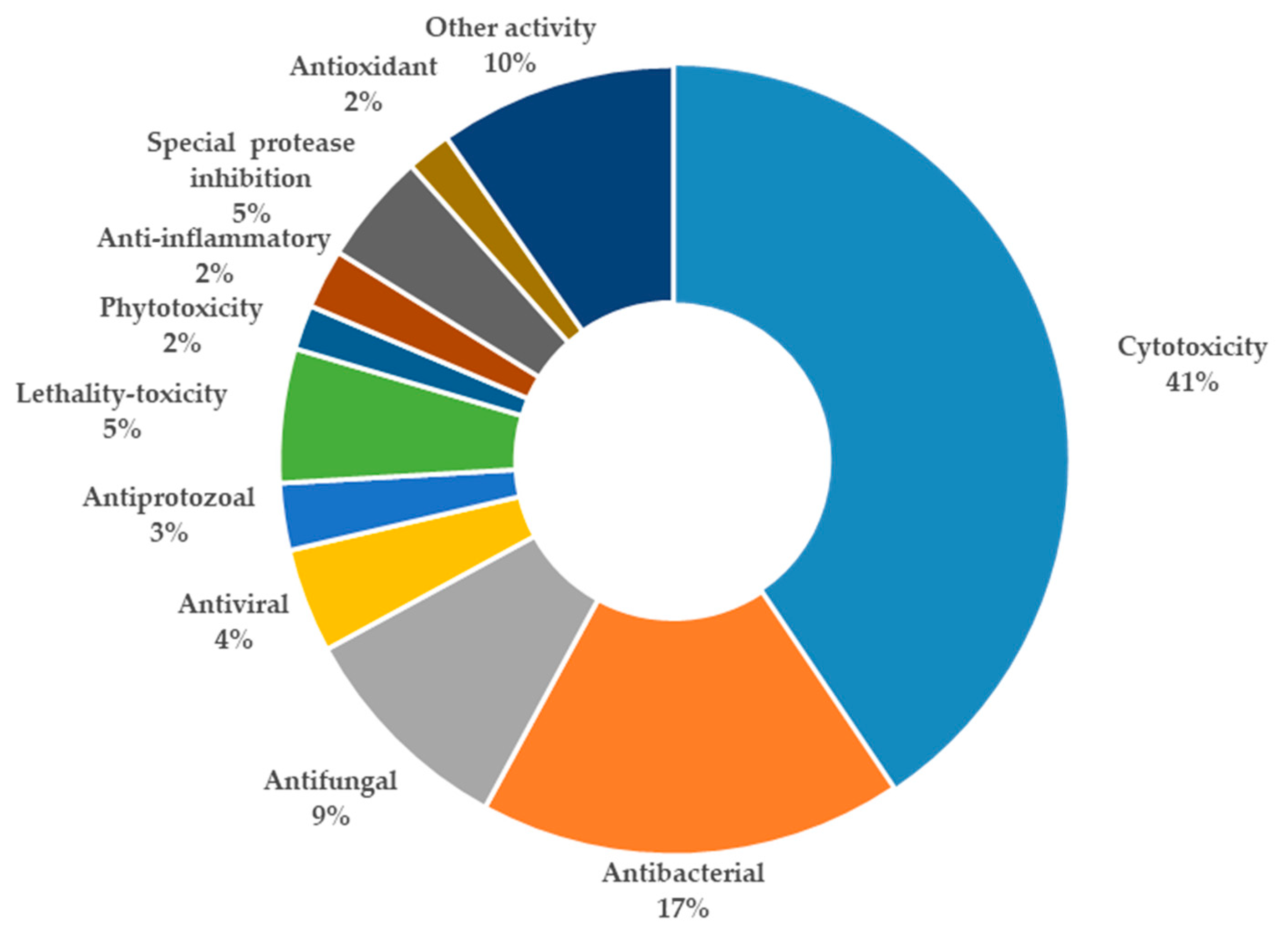
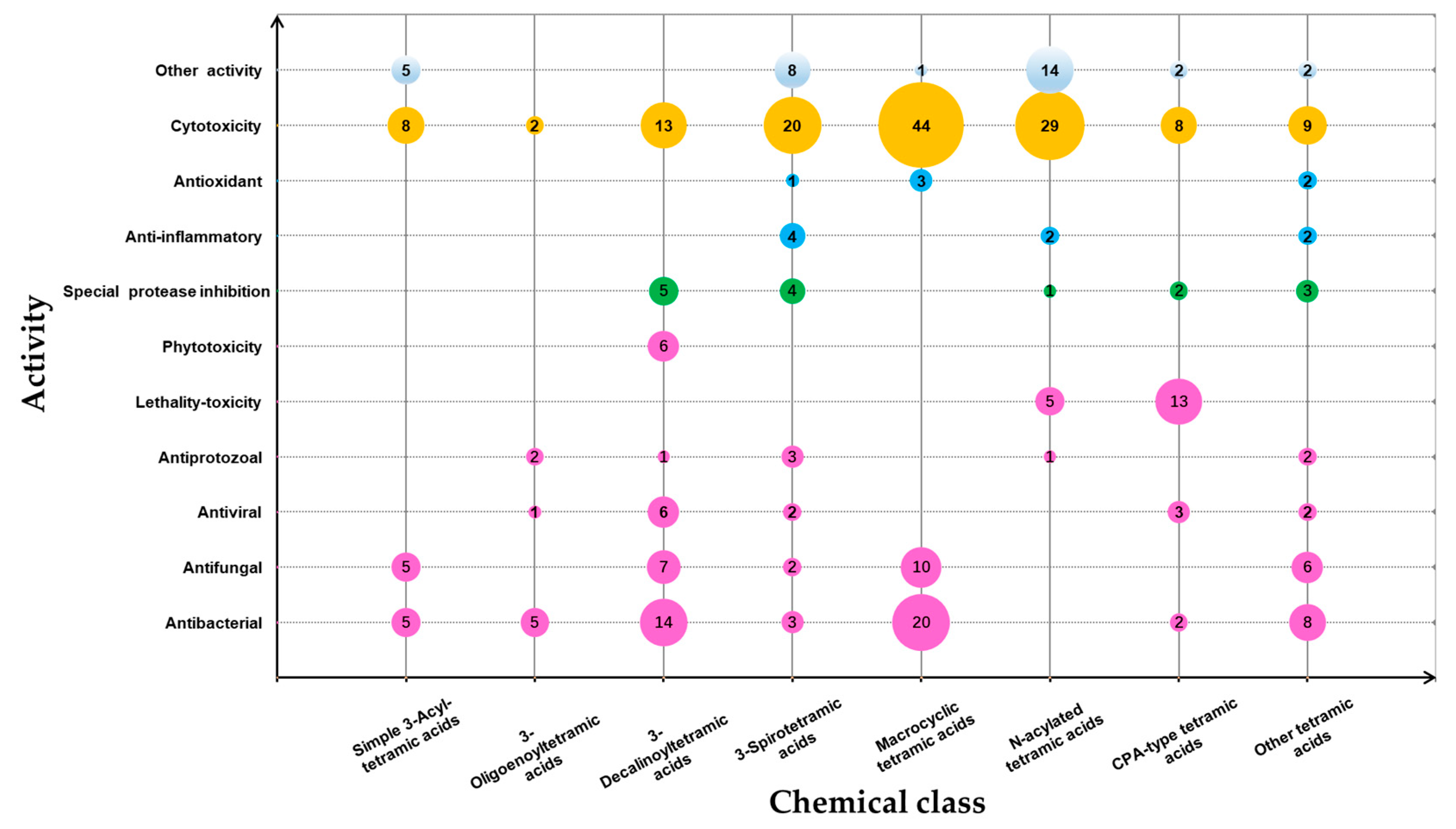
3. Summary of Tetramic Acid Products from Marine Microbes
4. Conclusions and Outlooks
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Royles, B.J. Naturally occurring tetramic acids: Structure, isolation, and synthesis. Chem. Rev. 1995, 95, 1981–2001. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. Bioactive Tetramic Acid Metabolites. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2003; Volume 28, pp. 109–163. [Google Scholar]
- Gossauer, A. Monopyrrolic natural compounds including tetramic acid derivatives. Chem. Org. Nat. 2003, 86, 1–188. [Google Scholar]
- Schobert, R.; Schlenk, A. Tetramic and tetronic acids: An update on new derivatives and biological aspects. Bioorg. Med. Chem. 2008, 16, 4203–4221. [Google Scholar] [CrossRef]
- Mo, X.; Li, Q.; Ju, J. Naturally occurring tetramic acid products: Isolation, structure elucidation and biological activity. RSC Adv. 2014, 4, 50566–50593. [Google Scholar] [CrossRef]
- Boettger, D.; Hertweck, C. Molecular diversity sculpted by fungal PKS-NRPS hybrids. Chembiochem. 2013, 14, 28–42. [Google Scholar] [CrossRef]
- Fisch, K.M. Biosynthesis of natural products by microbial iterative hybrid PKS–NRPS. RSC Adv. 2013, 3, 18228–18247. [Google Scholar] [CrossRef] [Green Version]
- Li, X.W.; Ear, A.; Nay, B. Hirsutellones and beyond: Figuring out the biological and synthetic logics toward chemical complexity in fungal PKS-NRPS compounds. Nat. Prod. Rep. 2013, 30, 765–782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, W.; Saha, S.; Zhang, C. Recent advances in discovery, biosynthesis and genome mining of medicinally relevant polycyclic tetramate macrolactams. Curr. Top. Med. Chem. 2016, 16, 1727–1739. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimundo, I.; Silva, S.G.; Costa, R.; Keller-Costa, T. Bioactive Secondary Metabolites from Octocoral-Associated Microbes-New Chances for Blue Growth. Mar. Drugs 2018, 16, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jessen, H.J.; Gademann, K. 4-Hydroxy-2-pyridone alkaloids: Structures and synthetic approaches. Nat. Prod. Rep. 2010, 27, 1168–1185. [Google Scholar] [CrossRef] [PubMed]
- Scherlach, K.; Boettger, D.; Remme, N.; Hertweck, C. The chemistry and biology of cytochalasans. Nat. Prod. Rep. 2010, 27, 869–886. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Zhu, M.; Deng, Z.; Sun, Y. Biosynthesis of tetronate antibiotics: A growing family of natural products with broad biological activities. Sci. China Chem. 2013, 56, 1364–1371. [Google Scholar] [CrossRef]
- Vieweg, L.; Reichau, S.; Schobert, R.; Leadlay, P.F.; Süssmuth, R.D. Recent advances in the field of bioactive tetronates. Nat. Prod. Rep. 2014, 31, 1554–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skellam, E. The biosynthesis of cytochalasans. Nat. Prod. Rep. 2017, 34, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Imamura, N.; Adachi, K.; Sano, H. Magnesidin A, a component of marine antibiotic magnesidin, produced by Vibrio gazogenes ATCC29988. J. Antibiot. 1994, 47, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.; Nazareth, J.; Divekar, P.; Kohl, H.; De Souza, N. Magnesidin, a novel magnesium-containing antibiotic. J. Antibiot. 1973, 26, 797–798. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.D.; Osterhage, C.; Konig, G.M. Epicoccamide, a novel secondary metabolite from a jellyfish-derived culture of Epicoccum purpurascens. Org. Biomol. Chem. 2003, 1, 507–510. [Google Scholar] [CrossRef]
- Lin, Z.-J.; Lu, Z.-Y.; Zhu, T.-J.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Penicillenols from Penicillium sp. GQ-7, an endophytic fungus associated with Aegiceras corniculatum. Chem. Pharm. Bull. 2008, 56, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-Q.; Chen, L.; Hu, X.; Gong, M.-W.; Zhang, W.-W.; Zheng, Q.-H.; Liu, Q.-Y. Novel Cytotoxic Metabolites from the Marine-derived Fungus Trichoderma citrinoviride. Heterocycles 2014, 89, 189–196. [Google Scholar]
- Zhang, Q.-Q.; Chen, L.; Zhou, T.; Zhao, Y.-Y.; Gong, M.-W.; Xia, Q.-W.; Ying, M.-G.; Zheng, Q.-H.; Liu, Q.-Y. Four New Penicitrinols and Two New Penicillenols from the Marine-Derived Fungus Penicillium citrinum. Heterocycles 2015, 91, 1007–1016. [Google Scholar]
- Kempf, K.; Schmitt, F.; Bilitewski, U.; Schobert, R. Synthesis, stereochemical assignment, and bioactivity of the Penicillium metabolites penicillenols B1 and B2. Tetrahedron 2015, 71, 5064–5068. [Google Scholar] [CrossRef]
- Zhang, Q.-Q.; Chen, L.; Huang, K.; Zhong, P.; Hu, X.; Fang, Z.-X.; Wu, J.-L. Tumonoic Acids K and L, Novel Metabolites from the Marine-Derived Fungus Penicillium citrinum. Heterocycles 2012, 85, 413–419. [Google Scholar]
- Wang, J.-F.; Qin, X.; Xu, F.-Q.; Zhang, T.; Liao, S.; Lin, X.; Yang, B.; Liu, J.; Wang, L.; Tu, Z.; et al. Tetramic acid derivatives and polyphenols from sponge-derived fungus and their biological evaluation. Nat. Prod. Res. 2015, 29, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yao, Q.F.; Amin, M.; Nong, X.H.; Zhang, X.Y.; Qi, S.H. Penicillenols from a deep-sea fungus Aspergillus restrictus inhibit Candida albicans biofilm formation and hyphal growth. J. Antibiot. 2017, 70, 763–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.Y.; Zhou, T.; Zhao, Y.Y.; Chen, L.; Gong, M.W.; Xia, Q.W.; Ying, M.G.; Zheng, Q.H.; Zhang, Q.Q. Antitumor Effects and Related Mechanisms of Penicitrinine A, a Novel Alkaloid with a Unique Spiro Skeleton from the Marine Fungus Penicillium citrinum. Mar. Drugs 2015, 13, 4733. [Google Scholar] [CrossRef]
- Shang, Z.; Li, L.; Esposito, B.P.; Salim, A.A.; Khalil, Z.G.; Quezada, M.; Bernhardt, P.V.; Capon, R.J. New PKS-NRPS tetramic acids and pyridinone from an Australian marine-derived fungus, Chaunopycnis sp. Org. Biomol. Chem. 2015, 13, 7795–7802. [Google Scholar] [CrossRef]
- Nakada, T.; Nakajima, M.; Kobayashi, H.; Takahashi, M.; Tanaka, I. Novel compound F-14329, its manufacture, Chaunopycnis species producing the compound, and its use for pharmaceuticals. Japanese Kokai Tokkyo Koho JP2007-153840, 21 June 2007. [Google Scholar]
- Kebede, B.; Wrigley, S.K.; Prashar, A.; Rahlff, J.; Wolf, M.; Reinshagen, J.; Gribbon, P.; Imhoff, J.F.; Silber, J.; Labes, A.; et al. Establishing the Secondary Metabolite Profile of the Marine Fungus: Tolypocladium geodes sp. MF458 and Subsequent Optimisation of Bioactive Secondary Metabolite Production. Mar. Drugs 2017, 15, 84. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.-H.; Nong, X.-H.; Liang, X.; Qi, S.-H. New tetramic acid derivatives from the deep-sea-derived fungus Cladosporium sp. SCSIO z0025. Tetrahedron 2018, 74, 2620–2626. [Google Scholar] [CrossRef]
- Liang, X.; Huang, Z.-H.; Ma, X.; Qi, S.-H. Unstable tetramic acid derivatives from the deep- sea-derived fungus Cladosporium sphaerospermum EIODSF 008. Mar. Drugs 2018, 16, 448. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Wang, L.; Wang, Y.; Wang, W.; Hao, J.; Zhu, W. Bioactive Natural Products of Aspergillus sp. OUCMDZ-1914, an Aciduric Fungus from the Mangrove Soils. Chin. J. Org. Chem. 2015, 35, 1955–1960. [Google Scholar] [CrossRef] [Green Version]
- Reusser, F. Tirandamycin, an inhibitor of bacterial ribonucleic acid polymerase. Antimicrob. Agents Chem. 1976, 10, 618–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reusser, F. Tirandamycin: Inhibition of oxidative phosphorylation in rat liver mitochondria. Infect. Immun. 1970, 2, 82–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, Y.; Kondo, K.; Ikeda, A.; Harada, R.; Dairi, T. Identification of tirandamycins as specific inhibitors of the futalosine pathway. J. Antibiot. 2017, 70, 798–800. [Google Scholar] [CrossRef]
- Carlson, J.C.; Li, S.; Burr, D.A.; Sherman, D.H. Isolation and characterization of tirandamycins from a marine-derived Streptomyces sp. J. Nat. Prod. 2009, 72, 2076–2079. [Google Scholar] [CrossRef] [Green Version]
- Duan, C.; Yao, Y.; Wang, Z.; Tian, X.; Zhang, S.; Zhang, C.; Ju, J. Fermentation optimization, isolation and identification of tirandamycins A and B from marine-derived Streptomyces sp. SCSIO 1666. Chin. J. Mar. Drugs 2010, 29, 12–20. [Google Scholar]
- Huang, X.; Kong, F.; Zhou, S.; Huang, D.; Zheng, J.; Zhu, W. Streptomyces tirandamycinicus sp. nov., a novel marine sponge-derived actinobacterium with antibacterial potential against Streptococcus agalactiae. Front. Microbiol. 2019, 10, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Rateb, M.E.; Yu, Z.; Yan, Y.; Yang, D.; Huang, T.; Vodanovic-Jankovic, S.; Kron, M.A.; Shen, B. Medium optimization of Streptomyces sp. 17944 for tirandamycin B production and isolation and structural elucidation of tirandamycins H., I and J. J. Antibiot. 2014, 67, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, A.; Socha, A.M.; Ryke, E.; Rowley, D.C. Antiamoebic properties of the actinomycete metabolites echinomycin A and tirandamycin A. Parasitol. Res. 2012, 111, 2473–2477. [Google Scholar] [CrossRef]
- Cong, Z.; Huang, X.; Liu, Y.; Liu, Y.; Wang, P.; Liao, S.; Yang, B.; Zhou, X.; Huang, D.; Wang, J. Cytotoxic anthracycline and antibacterial tirandamycin analogues from a marine-derived Streptomyces sp. SCSIO 41399. J. Antibiot. 2019, 72, 45–49. [Google Scholar] [CrossRef]
- Mo, X.; Wang, Z.; Wang, B.; Ma, J.; Huang, H.; Tian, X.; Zhang, S.; Zhang, C.; Ju, J. Cloning and characterization of the biosynthetic gene cluster of the bacterial RNA polymerase inhibitor tirandamycin from marine-derived Streptomyces sp. SCSIO1666. Biochem. Biophys. Res. Commun. 2011, 406, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.; Ma, J.; Huang, H.; Wang, B.; Song, Y.; Zhang, S.; Zhang, C.; Ju, J. Delta(11,12) double bond formation in tirandamycin biosynthesis is atypically catalyzed by TrdE, a glycoside hydrolase family enzyme. J. Am. Chem. Soc. 2012, 134, 2844–2847. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.; Huang, H.; Ma, J.; Wang, Z.; Wang, B.; Zhang, S.; Zhang, C.; Ju, J. Characterization of TrdL as a 10-hydroxy dehydrogenase and generation of new analogues from a tirandamycin biosynthetic pathway. Org. Lett. 2011, 13, 2212–2215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Z.; Du, L.; Chlipala, G.E.; Lopez, P.C.; Zhang, W.; Sherman, D.H.; Li, S. Identification of an unexpected shunt pathway product provides new insights into tirandamycin biosynthesis. Tetrahedron Lett. 2016, 57, 5919–5923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Han, X.; Wang, D.; Liu, M.; Gou, J.; Peng, Y.; Liu, J.; Li, Y.; Cao, F.; Zhang, C. Bioactive 3-decalinoyltetramic acids derivatives from a marine-derived strain of the fungus Fusarium equiseti D39. Front. Microbiol. 2019, 10, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Ming, Y.; Wang, L.; Li, Y.; Li, B.; Chen, J.; Shi, S. Local delivery of deep marine fungus-derived equisetin from polyvinylpyrrolidone (PVP) nanofibers for anti-MRSA activity. Chem. Eng. J. 2018, 350, 157–163. [Google Scholar] [CrossRef]
- Osterhage, C.; Kaminsky, R.; König, G.M.; Wright, A.D. Ascosalipyrrolidinone a, an antimicrobial alkaloid, from the obligate marine fungus Ascochyta s alicorniae. J. Org. Chem. 2000, 65, 6412–6417. [Google Scholar] [CrossRef]
- Daferner, M.; Anke, T.; Sterner, O. Zopfiellamides A and B, antimicrobial pyrrolidinone derivatives from the marine fungus Zopfiella latipes. Tetrahedron 2002, 58, 7781–7784. [Google Scholar] [CrossRef]
- Neumann, K.; Kehraus, S.; Guetschow, M.; Koenig, G.M. Cytotoxic and HLE-inhibitory tetramic acid derivatives from marine-derived fungi. Nat. Prod. Commun. 2009, 4, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.W.; Mierzwa, R.; Terracciano, J.; Patel, M.; Gullo, V.; Wagner, N.; Baroudy, B.; Puar, M.; Chan, T.M.; McPhail, A.T.; et al. Chemokine receptor CCR-5 inhibitors produced by Chaetomium globosum. J. Nat. Prod. 2006, 69, 1025–1028. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Wang, J.; Wang, Y.-F.; Zhang, X.-Y.; Nong, X.-H.; Chen, M.-Y.; Xu, X.-Y.; Qi, S.-H. Cytotoxic and antiviral tetramic acid derivatives from the deep-sea-derived fungus Trichobotrys effusa DFFSCS021. Tetrahedron 2015, 71, 9328–9332. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Labes, A.; Kramer, A.; Schmaljohann, R.; Imhoff, J.F. Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar Drugs 2015, 13, 4617–4632. [Google Scholar] [CrossRef] [Green Version]
- Ondeyka, J.G.; Smith, S.K.; Zink, D.L.; Vicente, F.; Basilio, A.; Bills, G.F.; Polishook, J.D.; Garlisi, C.; McGuinness, D.; Smith, E.; et al. Isolation, structure elucidation and antibacterial activity of a new tetramic acid, ascosetin. J. Antibiot. 2014, 67, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, A.L.; Berrué, F.; Overy, D.P.; Kerr, R.G. Isolation of iqalisetins A and B from a Tolypocladium sp. isolated from marine sediment from Frobisher Bay in Canada’s arctic. Can. J. Chem. 2016, 94, 444–448. [Google Scholar] [CrossRef]
- Furumai, T.; Eto, K.; Sasaki, T.; Higuchi, H.; Onaka, H.; Saito, N.; Fujita, T.; Naoki, H.; Igarashi, Y. TPU-0037-A, B, C and D, novel lydicamycin congeners with anti-MRSA activity from Streptomyces platensis TP-A0598. J. Antibiot. 2002, 55, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, Y.; Kanamaru, N.; Shimazu, A.; Seto, H. Lydicamycin, a new antibiotic of a novel skeletal type. I. Taxonomy, fermentation, isolation and biological activity. J. Antibiot. 1991, 44, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Amagata, T.; Xiao, J.; Chen, Y.P.; Holsopple, N.; Oliver, A.G.; Gokey, T.; Guliaev, A.B.; Minoura, K. Creation of an HDAC-based yeast screening method for evaluation of marine-derived actinomycetes: Discovery of streptosetin A. J. Nat. Prod. 2012, 75, 2193–2199. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Cui, Y.; Cai, C.; Chen, H.; Dai, Y.; Chen, P.; Kong, F.; Yuan, J.; Song, X.; Mei, W.; et al. Two New Succinimide Derivatives Cladosporitins A and B from the Mangrove-derived Fungus Cladosporium sp. HNWSW-1. Mar. Drugs 2018, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Kikuchi, T.; Tanaka, R. Altercrasin A, a novel decalin derivative with spirotetramic acid, produced by a sea urchin-derived Alternaria sp. Tetrahedron Lett. 2015, 56, 1229–1232. [Google Scholar] [CrossRef]
- Yamada, T.; Tanaka, A.; Nehira, T.; Nishii, T.; Kikuchi, T. Altercrasins A–E, Decalin Derivatives, from a Sea-Urchin-Derived Alternaria sp.: Isolation and Structural Analysis Including Stereochemistry. Mar. Drugs 2019, 17, 218. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.H.; Asami, Y.; Jang, J.P.; Kim, S.O.; Moon, D.O.; Shin, K.S.; Hashizume, D.; Muroi, M.; Saito, T.; Oh, H.; et al. Fusarisetin A, an acinar morphogenesis inhibitor from a soil fungus, Fusarium sp. FN080326. J. Am. Chem. Soc. 2011, 133, 6865–6867. [Google Scholar] [CrossRef]
- Caro-Diaz, E.J.; Aung, A.; Xu, J.; Varghese, S.; Theodorakis, E.A. Fusarisetins: Structure-function studies on a novel class of cell migration inhibitors. Org. Chem. Front. 2014, 1, 135–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Caro-Diaz, E.J.; Lacoske, M.H.; Hung, C.I.; Jamora, C.; Theodorakis, E.A. Fusarisetin A: Scalable Total Synthesis and Related Studies. Chem. Sci. 2012, 3, 3378–3386. [Google Scholar] [CrossRef] [Green Version]
- Bloch, P.; Tamm, C.; Bollinger, P.; Petcher, T.J.; Weber, H.P. Pseurotin, a New Metabolite of Pseudeurotium ovalis STOLK Having an Unusual Hetero-Spirocyclic System. Helv. Chim. Acta. 1976, 59, 133–137. [Google Scholar] [CrossRef]
- Boot, C.M.; Gassner, N.C.; Compton, J.E.; Tenney, K.; Tamble, C.M.; Lokey, R.S.; Holman, T.R.; Crews, P. Pinpointing pseurotins from a marine-derived Aspergillus as tools for chemical genetics using a synthetic lethality yeast screen. J. Nat. Prod. 2007, 70, 1672–1675. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.-L.; Fang, Y.-C.; Zhu, T.-J.; Gu, Q.-Q.; Zhu, W.-M. Cytotoxic alkaloids and antibiotic nordammarane triterpenoids from the marine-derived fungus Aspergillus sydowi. J. Nat. Prod. 2008, 71, 985–989. [Google Scholar] [CrossRef]
- Saraiva, N.N.; Rodrigues, B.S.; Jimenez, P.C.; Guimaraes, L.A.; Torres, M.C.; Rodrigues-Filho, E.; Pfenning, L.H.; Abreu, L.M.; Mafezoli, J.; de Mattos, M.C.; et al. Cytotoxic compounds from the marine-derived fungus Aspergillus sp. recovered from the sediments of the Brazilian coast. Nat. Prod. Res. 2015, 29, 1545–1550. [Google Scholar] [CrossRef]
- Lee, M.S.; Wang, S.W.; Wang, G.J.; Pang, K.L.; Lee, C.K.; Kuo, Y.H.; Cha, H.J.; Lin, R.K.; Lee, T.H. Angiogenesis Inhibitors and Anti-Inflammatory Agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef]
- Anjum, K.; Bi, H.; Chai, W.; Lian, X.Y.; Zhang, Z. Antiglioma pseurotin A from marine Bacillus sp. FS8D regulating tumour metabolic enzymes. Nat. Prod. Res. 2018, 32, 1353–1356. [Google Scholar] [CrossRef]
- Maebayashi, Y.; Horie, Y.; Satoh, Y.; Yamazaki, M. Isolation of pseurotin A and a new pyrazine from Pseudallescheria boydii. JSM Mycotoxins 1985, 1985, 33–34. [Google Scholar] [CrossRef] [Green Version]
- Wink, J. Biologically active pseurotin A and D, new metabolites from Aspergillus fumigatus, process for their preparation and their use as apomorphine antagonists. European Patent EP0546475, 16 June 2003. [Google Scholar]
- Wenke, J.; Anke, H.; Sterner, O. Pseurotin A and 8-O-demethylpseurotin A from Aspergillus fumigatus and their inhibitory activities on chitin synthase. Biosci. Biotechnol. Biochem. 1993, 57, 961–964. [Google Scholar] [CrossRef]
- Komagata, D.; Fujita, S.; Yamashita, N.; Saito, S.; Morino, T. Novel neuritogenic activities of pseurotin A and penicillic acid. J. Antibiot. 1996, 49, 958–959. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, A.; Fujioka, S.; Nukina, M.; Kawano, T.; Shimada, A.; Kimura, Y. Fumiquinones A and B, nematicidal quinones produced by Aspergillus fumigatus. Biosci. Biotechnol. Biochem. 2007, 71, 1697–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, M.; Ninomiya, T.; Akabane, H.; Kushida, N.; Tsujiuchi, G.; Ohyama, M.; Gomi, S.; Shito, K.; Murata, T. Pseurotin A and its analogues as inhibitors of immunoglobulin E [correction of immunoglobuline E] production. Bioorg. Med. Chem. Lett. 2009, 19, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Luis, S.; Cherigo, L.; Arnold, E.; Spadafora, C.; Gerwick, W.H.; Cubilla-Rios, L. Antiparasitic and anticancer constituents of the endophytic fungus Aspergillus sp. strain F1544. Nat. Prod. Commun. 2012, 7, 165–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, E.A.A.; Carvalho, J.M.; dos Santos, D.C.P.; Feitosa, A.d.O.; Marinho, P.S.B.; Guilhon, G.M.S.P.; de Souza, A.D.L.; da Silva, F.M.A.; Marinho, A.M.d.R. Antibacterial activity of alkaloids produced by endophytic fungus Aspergillus sp. EJC08 isolated from medical plant Bauhinia guianensis. Nat. Prod. Res. 2013, 27, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Mehedi, M.A.U.; Molla, A.H.; Khondkar, P.; Sultana, S.; Islam, M.A.; Rashid, M.A.; Chowdhury, R. Pseurotin A: An Antibacterial Secondary Metabolite from Aspergillus fumigatus. Asian J. Chem. 2010, 22, 2611–2614. [Google Scholar]
- Lu, Q.Q.; Tian, J.M.; Wei, J.; Gao, J.M. Bioactive metabolites from the mycelia of the basidiomycete Hericium erinaceum. Nat. Prod. Res. 2014, 28, 1288–1292. [Google Scholar] [CrossRef]
- Chen, K.; Qiu, P.; Yuan, Y.; Zheng, L.; He, J.; Wang, C.; Guo, Q.; Kenny, J.; Liu, Q.; Zhao, J.; et al. Pseurotin A Inhibits Osteoclastogenesis and Prevents Ovariectomized-Induced Bone Loss by Suppressing Reactive Oxygen Species. Theranostics 2019, 9, 1634–1650. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Bai, J.; Chen, Z.; Hua, H.; Liu, T. Alkaloids from the Marine-Derived Fungus Aspergillus fumigatus YK-7 and Their Antitumor Activities. Chin. Pharm. J. 2017, 52, 1308–1312. [Google Scholar]
- Wang, F.-Z.; Li, D.-H.; Zhu, T.-J.; Zhang, M.; Gu, Q.-Q. Pseurotin A1 and A2, two new 1-oxa-7-azaspiro[4.4]non-2-ene-4,6-diones from the holothurian-derived fungus Aspergillus fumigatus WFZ-25. Can. J. Chem. 2011, 89, 72–76. [Google Scholar] [CrossRef]
- Yamada, T.; Ohshima, M.; Yuasa, K.; Kikuchi, T.; Tanaka, R. Assignment of the CD Cotton Effect to the Chiral Center in Pseurotins, and the Stereochemical Revision of Pseurotin A (2). Mar. Drugs 2016, 14, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wink, J. Pseurotin F1/F2, new metabolites from Aspergillus fumigatus, process for their preparation and their use as apomorphine antagonists. European Patent Application JP EP546474, 16 June 1993. [Google Scholar]
- Ren, H.; Liu, R.; Chen, L.; Zhu, T.; Zhu, W.M.; Gu, Q.Q. Two new hetero-spirocyclic gamma-lactam derivatives from marine sediment-derived fungus Aspergillus sydowi D2-6. Arch. Pharm. Res. 2010, 33, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Tao, G.-Y.; Liu, J.; Jung, J.H.; Guo, W.; Wen, X.-Q.; Liu, Y. Compounds from a jellyfish-derived fungus Aspergillus fumigates. Nat. Prod. Sci. 2016, 22, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Rateb, M.; Tabudravu, J.N.; Perez-Bonilla, M.; Dirkx, N.; Vallorani, R.; Diaz, C.; Perez Del Palacio, J.; Smith, A.J.; Ebel, R.; et al. Zebrafish-Based Discovery of Antiseizure Compounds from the Red Sea: Pseurotin A2 and Azaspirofuran, A. ACS Chem. Neurosci. 2018, 9, 1652–1662. [Google Scholar] [CrossRef] [Green Version]
- Wakefield, J.; Hassan, H.M.; Jaspars, M.; Ebel, R.; Rateb, M.E. Dual Induction of New Microbial Secondary Metabolites by Fungal Bacterial Co-cultivation. Front. Microbiol. 2017, 8, 1284–1293. [Google Scholar] [CrossRef] [Green Version]
- Rischer, M.; Lee, S.R.; Eom, H.J.; Park, H.B.; Vollmers, J.; Kaster, A.-K.; Shin, Y.-H.; Oh, D.-C.; Kim, K.H.; Beemelmanns, C. Spirocyclic cladosporicin A and cladosporiumins I and J from a Hydractinia-associated Cladosporium sphaerospermum SW67. Org. Chem. Front. 2019, 6, 1084–1093. [Google Scholar] [CrossRef]
- Yamada, T.; Imai, E.; Nakatuji, K.; Numata, A.; Tanaka, R. Cephalimysin A, a potent cytotoxic metabolite from an Aspergillus species separated from a marine fish. Tetrahedron Lett. 2007, 48, 6294–6296. [Google Scholar] [CrossRef]
- Yamada, T.; Kitada, H.; Kajimoto, T.; Numata, A.; Tanaka, R. The relationship between the CD Cotton effect and the absolute configuration of FD-838 and its seven stereoisomers. J. Org. Chem. 2010, 75, 4146–4153. [Google Scholar] [CrossRef]
- Yamada, T.; Kimura, H.; Arimitsu, K.; Kajimoto, T.; Kikuchi, T.; Tanaka, R. Absolute Configuration of Eight Cephalimysins Isolated from the Marine-Derived Aspergillus fumigatus. Chem. Sel. 2017, 2, 10936–10940. [Google Scholar] [CrossRef]
- Yamada, T.; Kajimoto, T.; Kikuchi, T.; Tanaka, R. Elucidation of the Relationship between CD Cotton Effects and the Absolute Configuration of Sixteen Stereoisomers of Spiroheterocyclic-Lactams. Mar. Drugs 2018, 16, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoue, K.; Okazaki, T.; Hanada, K.; Amamoto, T.; Yamagishi, M.; Omura, S. Physiologically active substance FD-838 and process for preparing the same. Europe Patent EP0216607, 1 April 1987. [Google Scholar]
- Xu, X.; Han, J.; Wang, Y.; Lin, R.; Yang, H.; Li, J.; Wei, S.; Polyak, S.W.; Song, F. Two New Spiro-Heterocyclic gamma-Lactams from A Marine-Derived Aspergillus fumigatus Strain CUGBMF170049. Mar. Drugs 2019, 17, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, F.; Liu, Y.; Liu, Y.; Li, K.; Yang, X.; Liu, S.; Zhou, X.; Yang, J. Spirostaphylotrichin x from a marine-derived fungus as an anti-influenza agent targeting RNA polymerase PB2. J. Nat. Prod. 2018, 81, 2722–2730. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Luo, M.; Liu, F.; Wang, D.; Pang, X.; Zhao, T.; Xu, L.; Wu, X.; Xia, M.; Yang, X. Cytochalasan and tyrosine-derived alkaloids from the marine sediment-derived fungus Westerdykella dispersa and their bioactivities. Sci. Rep. 2017, 7, 11956. [Google Scholar] [CrossRef] [Green Version]
- Quezada, M.; Licona-Cassani, C.; Cruz-Morales, P.; Salim, A.A.; Marcellin, E.; Capon, R.J.; Barona-Gomez, F. Diverse cone-snail species harbor closely related Streptomyces species with conserved chemical and genetic profiles, including polycyclic tetramic acid macrolactams. Front. Microbiol. 2017, 8, 2305–2317. [Google Scholar] [CrossRef]
- Li, Y.; Chen, H.; Ding, Y.; Xie, Y.; Wang, H.; Cerny, R.L.; Shen, Y.; Du, L. Iterative assembly of two separate polyketide chains by the same single-module bacterial polyketide synthase in the biosynthesis of HSAF. Angew. Chem. Int. Ed. Engl 2014, 53, 7524–7530. [Google Scholar] [CrossRef] [Green Version]
- Blodgett, J.A.; Oh, D.C.; Cao, S.; Currie, C.R.; Kolter, R.; Clardy, J. Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 11692–11697. [Google Scholar] [CrossRef] [Green Version]
- Jomon, K.; Kuroda, Y.; Ajisaka, M.; Sakai, H. A new antibiotic, ikarugamycin. J. Antibiot. 1972, 25, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Hirata, Y. The structure of ikarugamycin, an acyltetramic acid antibiotic possessing a unique as-hydrindacene skeleton. Bull. Chem. Soc. Jpn. 1977, 50, 1813–1820. [Google Scholar] [CrossRef] [Green Version]
- Bertasso, M.; Holzenkaempfer, M.; Zeeck, A.; Stackebrandt, E.; Beil, W.; Fiedler, H.-P. Ripromycin and other polycyclic macrolactams from Streptomyces sp. Tü 6239. J. Antibiot. 2003, 56, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Lacret, R.; Oves-Costales, D.; Gomez, C.; Diaz, C.; de la Cruz, M.; Perez-Victoria, I.; Vicente, F.; Genilloud, O.; Reyes, F. New ikarugamycin derivatives with antifungal and antibacterial properties from Streptomyces zhaozhouensis. Mar. Drugs 2014, 13, 128–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, R.; Heiss, E.H.; Ferk, F.; Peschel, A.; Knasmueller, S.; Dirsch, V.M.; Krupitza, G.; Kopp, B. Ikarugamycin induces DNA damage, intracellular calcium increase, p38 MAP kinase activation and apoptosis in HL-60 human promyelocytic leukemia cells. Mutat. Res. Fund. Mol. Mech. Mutagen. 2011, 709, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Dhaneesha, M.; Hasin, O.; Sivakumar, K.C.; Ravinesh, R.; Naman, C.B.; Carmeli, S.; Sajeevan, T.P. DNA binding and molecular dynamic studies of polycyclic tetramate macrolactams (PTM) with potential anticancer activity isolated from a sponge-associated Streptomyces zhaozhouensis subsp. mycale subsp. nov. Mar. Biotechnol. 2019, 21, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.L.; Jiang, S.H.; Bu, X.L.; Wang, J.H.; Weng, J.Y.; Yang, X.M.; He, K.Y.; Zhang, Z.G.; Ao, P.; Xu, J.; et al. Structural diversity of anti-pancreatic cancer capsimycins identified in mangrove-derived Streptomyces xiamenensis 318 and post-modification via a novel cytochrome P450 monooxygenase. Sci. Rep. 2017, 7, 40689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, G.; Zhang, L.; Liu, W.; Jiang, X.; Jin, H.; Liu, Z.; Zhang, H.; Zhou, A.; Zhang, C. New polycyclic tetramate macrolactams from marine-derived Streptomyces sp. SCSIO 40060. Tetrahedron 2018, 74, 6839–6845. [Google Scholar] [CrossRef]
- Hasumi, K.; Shinohara, C.; Naganuma, S.; Endo, A. Inhibition of the uptake of oxidized low-density lipoprotein in macrophage J774 by the antibiotic ikarugamycin. Eur. J. Biochem. 1992, 205, 841–846. [Google Scholar] [CrossRef]
- Luo, T.; Fredericksen, B.L.; Hasumi, K.; Endo, A.; Garcia, J.V. Human immunodeficiency virus type 1 Nef-induced CD4 cell surface downregulation is inhibited by ikarugamycin. J. Virol. 2001, 75, 2488–2492. [Google Scholar] [CrossRef] [Green Version]
- Moscatelli, A.; Ciampolini, F.; Rodighiero, S.; Onelli, E.; Cresti, M.; Santo, N.; Idilli, A. Distinct endocytic pathways identified in tobacco pollen tubes using charged nanogold. J. Cell. Sci. 2007, 120, 3804–3819. [Google Scholar] [CrossRef] [Green Version]
- Kyeremeh, K.; Acquah, K.S.; Sazak, A.; Houssen, W.; Tabudravu, J.; Deng, H.; Jaspars, M. Butremycin, the 3-hydroxyl derivative of ikarugamycin and a protonated aromatic tautomer of 5′-methylthioinosine from a Ghanaian Micromonospora sp. K310. Mar. Drugs 2014, 12, 999–1012. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, S.; Akutsu, H.; Satomi, T.; Nagatsu, T.; Taguchi, R.; Seino, A. Capsimycin, a new antibiotic. I. Production, isolation and properties. J. Antibiot. 1979, 32, 193–196. [Google Scholar] [CrossRef]
- Fukuda, T.; Takahashi, M.; Kasai, H.; Nagai, K.; Tomoda, H. Chlokamycin, a New Chloride from the Marine-derived Streptomyces sp MA2-12. Nat. Prod. Commun. 2017, 12, 1223–1226. [Google Scholar] [CrossRef] [Green Version]
- Mei, X.; Wang, L.; Wang, D.; Fan, J.; Zhu, W. Polycyclic Tetramate Macrolactams from the Marine-Derived Actinoalloteichus cyanogriseus WH1-2216-6. Chin. J. Org. Chem. 2017, 37, 2352–2360. [Google Scholar] [CrossRef] [Green Version]
- Graupner, P.; Thornburgh, S.; Mathieson, J.; Chapin, E.; Kemmitt, G.; Brown, J.; Snipes, C. Dihydromaltophilin; A novel fungicidal tetramic acid containing metabolite from Streptomyces sp. J. Antibiot. 1997, 50, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zaleta-Rivera, K.; Zhu, X.; Huffman, J.; Millet, J.C.; Harris, S.D.; Yuen, G.; Li, X.C.; Du, L. Structure and biosynthesis of heat-stable antifungal factor (HSAF), a broad-spectrum antimycotic with a novel mode of action. Antimicrob. Agents Chemother. 2007, 51, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Zhang, W.; Zhang, G.; Zhu, Y.; Chen, Y.; Liu, W.; Yuan, C.; Zhang, Q.; Zhang, H.; Zhang, L.; et al. Activation and characterization of a cryptic gene cluster reveals a cyclization cascade for polycyclic tetramate macrolactams. Chem. Sci. 2017, 8, 1607–1612. [Google Scholar] [CrossRef] [Green Version]
- Shigemori, H.; Bae, M.A.; Yazawa, K.; Sasaki, T.; Kobayashi, J. Alteramide A, a new tetracyclic alkaloid from a bacterium Alteromonas sp. associated with the marine sponge Halichondria okadai. J. Org. Chem. 1992, 57, 4317–4320. [Google Scholar] [CrossRef]
- Moree, W.J.; McConnell, O.J.; Nguyen, D.D.; Sanchez, L.M.; Yang, Y.L.; Zhao, X.; Liu, W.T.; Boudreau, P.D.; Srinivasan, J.; Atencio, L.; et al. Microbiota of healthy corals are active against fungi in a light-dependent manner. ACS Chem. Biol. 2014, 9, 2300–2308. [Google Scholar] [CrossRef]
- Olano, C.; Garcia, I.; Gonzalez, A.; Rodriguez, M.; Rozas, D.; Rubio, J.; Sanchez-Hidalgo, M.; Brana, A.F.; Mendez, C.; Salas, J.A. Activation and identification of five clusters for secondary metabolites in Streptomyces albus J1074. Microb. Biotechnol. 2014, 7, 242–256. [Google Scholar] [CrossRef] [Green Version]
- Bae, M.; Yamada, K.; Uemura, D.; Seu, J.-H.; Kim, Y.-H. Aburatubolactam C, a novel apoptosis-inducing substance produced by marine Streptomyces sp. SCRC A-20. J. Microbiol. Biotech. 1998, 8, 455–460. [Google Scholar]
- Bae, M.-A.; Yamada, K.; Ijuin, Y.; Tsuji, T.; Yazawa, K.; Uemura, D. Aburatubolactam A, a novel inhibitor of superoxide anion generation from a marine microorganism. Heterocycl. Commun. 1992, 2, 315–318. [Google Scholar] [CrossRef]
- Yamada, K.; Kuramoto, M.; Uemura, D. Aburatubolactams and Zoanthamines, naturally occurring bioactive alkaloids. Recent Res. Devel. Pure. Appl. Chem 1999, 3, 245–254. [Google Scholar]
- Song, T.; Chen, M.; Chai, W.; Zhang, Z.; Lian, X.-Y. New bioactive pyrrospirones C−I from a marine-derived fungus Penicillium sp. ZZ380. Tetrahedron 2018, 74, 884–891. [Google Scholar] [CrossRef]
- Song, T.; Chen, M.; Ge, Z.W.; Chai, W.; Li, X.C.; Zhang, Z.; Lian, X.Y. Bioactive Penicipyrrodiether A, an Adduct of GKK1032 Analogue and Phenol A Derivative, from a Marine-Sourced Fungus Penicillium sp. ZZ380. J. Org. Chem. 2018, 83, 13395–13401. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Tang, M.; Ge, H.; Chen, M.; Lian, X.; Zhang, Z. Novel Bioactive Penicipyrroether A and Pyrrospirone J from the Marine-Derived Penicillium sp. ZZ380. Mar. Drugs 2019, 17, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Li, X.; Zhao, J.; He, N.; Li, Y.; Zhang, T.; Wang, S.; Yu, L.; Xie, Y. GKK1032C, a new alkaloid compound from the endophytic fungus Penicillium sp. CPCC 400817 with activity against methicillin-resistant S. aureus. J. Antibiot. 2019, 72, 237–240. [Google Scholar] [CrossRef]
- Chen, S.; Shen, H.; Zhang, P.; Cheng, H.; Dai, X.; Liu, L. Anti-glioma trichobamide A with an unprecedented tetrahydro-5H-furo[2,3-b]pyrrol-5-one functionality from ascidian-derived fungus Trichobotrys effuse 4729. Chem. Commun. 2019, 55, 1438–1441. [Google Scholar] [CrossRef]
- Wijeratne, E.M.; He, H.; Franzblau, S.G.; Hoffman, A.M.; Gunatilaka, A.A. Phomapyrrolidones A-C, antitubercular alkaloids from the endophytic fungus Phoma sp. NRRL 46751. J. Nat. Prod. 2013, 76, 1860–1865. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Z.; Huang, Y.; Liu, L.; He, J.; Wang, L.; Yuan, J.; She, Z. Ascomylactams A-C, Cytotoxic 12- or 13-Membered-Ring Macrocyclic Alkaloids Isolated from the Mangrove Endophytic Fungus Didymella sp. CYSK-4, and Structure Revisions of Phomapyrrolidones A and C. J. Nat. Prod. 2019, 82, 1752–1758. [Google Scholar] [CrossRef]
- Luesch, H.; Harrigan, G.; Goetz, G.; Horgen, F. The cyanobacterial origin of potent anticancer agents originally isolated from sea hares. Curr. Med. Chem. 2002, 9, 1791–1806. [Google Scholar] [CrossRef]
- Edwards, D.J.; Marquez, B.L.; Nogle, L.M.; McPhail, K.; Goeger, D.E.; Roberts, M.A.; Gerwick, W.H. Structure and biosynthesis of the jamaicamides, new mixed polyketide-peptide neurotoxins from the marine cyanobacterium Lyngbya majuscula. Chem. Biol. 2004, 11, 817–833. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, P.D.; Monroe, E.A.; Mehrotra, S.; Desfor, S.; Korobeynikov, A.; Sherman, D.H.; Murray, T.F.; Gerwick, L.; Dorrestein, P.C.; Gerwick, W.H. Expanding the Described Metabolome of the Marine Cyanobacterium Moorea producens JHB through Orthogonal Natural Products Workflows. PLoS ONE 2015, 10, e0133297. [Google Scholar] [CrossRef] [PubMed]
- Koehn, F.E.; Longley, R.E.; Reed, J.K. Microcolins A and B, new immunosuppressive peptides from the blue-green alga Lyngbya majuscula. J. Nat. Prod. 1992, 55, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Meickle, T.; Matthew, S.; Ross, C.; Luesch, H.; Paul, V. Bioassay-guided isolation and identification of desacetylmicrocolin B from Lyngbya cf. polychroa. Plant. Med. 2009, 75, 1427–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.B.; Glukhov, E.; Li, Y.; Iwasaki, A.; Gerwick, L.; Dorrestein, P.C.; Jiao, B.H.; Gerwick, W.H. Cytotoxic Microcolin Lipopeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2019, 82, 2608–2619. [Google Scholar] [CrossRef]
- Takamatsu, S.; Nagle, D.G.; Gerwick, W.H. Secondary metabolites from marine cyanobacteria and algae inhibit LFA-1/ICAM-1 mediated cell adhesion. Plant. Med. 2004, 70, 127–131. [Google Scholar]
- Zhang, L.H.; Longley, R.E.; Koehn, F.E. Antiproliferative and immunosuppressive properties of microcolin A, a marine-derived lipopeptide. Life Sci. 1997, 60, 751–762. [Google Scholar] [CrossRef]
- Moore, R.E.; Entzeroth, M. Majusculamide D and deoxymajusculamide D, two cytotoxins from Lyngbya majuscula. Phytochemistry 1988, 27, 3101–3103. [Google Scholar] [CrossRef]
- Caro-Diaz, E.J.E.; Valeriote, F.A.; Gerwick, W.H. Highly Convergent Total Synthesis and Assignment of Absolute Configuration of Majusculamide D, a Potent and Selective Cytotoxic Metabolite from Moorea sp. Org. Lett. 2019, 21, 793–796. [Google Scholar] [CrossRef]
- Nagle, D.G.; Paul, V.J.; Roberts, M.A. Ypaoamide, a new broadly acting feeding deterrent from the marine cyanobacterium Lyngbya majuscula. Tetrahedron Lett. 1996, 37, 6263–6266. [Google Scholar] [CrossRef]
- Sueyoshi, K.; Yamada, M.; Yamano, A.; Ozaki, K.; Sumimoto, S.; Iwasaki, A.; Suenaga, K.; Teruya, T. Ypaoamides B and C, Linear Lipopeptides from an Okeania sp. Marine Cyanobacterium. J. Nat. Prod. 2018, 81, 1103–1107. [Google Scholar] [CrossRef]
- Pereira, A.R.; Etzbach, L.; Engene, N.; Muller, R.; Gerwick, W.H. Molluscicidal metabolites from an assemblage of Palmyra Atoll cyanobacteria. J. Nat. Prod. 2011, 74, 1175–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Malyngamide A, a novel chlorinated metabolite of the marine cyanophyte Lyngbya majuscula. J. Am. Chem. Soc. 1979, 101, 240–242. [Google Scholar] [CrossRef]
- Dalietos, D.; Marner, F.-J.; Mynderse, J.S.; Moore, R.E. (–)-Trans-7 (S)-methoxytetradec-4-enoic acid and related amides from the marine cyanophyte Lyngbya majuscula. Phytochemistry 1978, 17, 2091–2095. [Google Scholar]
- Simmons, C.J.; Marner, F.-J.; Cardellina II, J.H.; Moore, R.E.; Seff, K. Pukeleimide C, a novel pyrrolic compound from the marine cyanophyte Lyngbya majuscula. Tetrahedron Lett. 1979, 20, 2003–2006. [Google Scholar] [CrossRef]
- Cardellina II, J.H.; Moore, R.E. The structures of pukeleimides A., B., D., E., F., and G. Tetrahedron Lett. 1979, 20, 2007–2010. [Google Scholar] [CrossRef]
- Milligan, K.E.; Marquez, B.; Williamson, R.T.; Davies-Coleman, M.; Gerwick, W.H. Two new malyngamides from a Madagascan Lyngbya majuscula. J. Nat. Prod. 2000, 63, 965–968. [Google Scholar] [CrossRef]
- Kan, Y.; Sakamoto, B.; Fujita, T.; Nagai, H. New malyngamides from the Hawaiian cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 1599–1602. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T.A.; McPhail, K.L.; Elbandy, M. Malyngamide 4, a new lipopeptide from the Red Sea marine cyanobacterium Moorea producens (formerly Lyngbya majuscula). Phytochem Lett. 2013, 6, 183–188. [Google Scholar] [CrossRef]
- Chang, T.T.; More, S.V.; Lu, I.H.; Hsu, J.C.; Chen, T.J.; Jen, Y.C.; Lu, C.K.; Li, W.S. Isomalyngamide A, A-1 and their analogs suppress cancer cell migration in vitro. Eur. J. Med. Chem. 2011, 46, 3810–3819. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, W.; Othman, R.; Uchida, H.; Watanabe, R.; Suzuki, T.; Sakamoto, B.; Nagai, H. A new malyngamide from the marine cyanobacterium Moorea producens. Nat. Prod. Res. 2018, 32, 97–104. [Google Scholar] [CrossRef]
- Luesch, H.; Toshida, W.Y.; Moore, R.E.; Paul, V.J. Structurally diverse new alkaloids from Palauan collections of the apratoxin-producing marine cyanobacterium Lyngbya sp. Tetrahedron 2002, 58, 7959–7966. [Google Scholar] [CrossRef]
- Simmons, T.L.; McPhail, K.L.; Ortega-Barría, E.; Mooberry, S.L.; Gerwick, W.H. Belamide A, a new antimitotic tetrapeptide from a Panamanian marine cyanobacterium. Tetrahedron Lett. 2006, 47, 3387–3390. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Imperial, L.; Garst, C.; Ratnayake, R.; Dang, L.H.; Paul, V.J.; Luesch, H. Caldoramide, a Modified Pentapeptide from the Marine Cyanobacterium Caldora penicillata. J. Nat. Prod. 2016, 79, 1867–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taori, K.; Liu, Y.; Paul, V.J.; Luesch, H. Combinatorial strategies by marine cyanobacteria: Symplostatin 4, an antimitotic natural dolastatin 10/15 hybrid that synergizes with the coproduced HDAC inhibitor largazole. Chembiochem 2009, 10, 1634–1639. [Google Scholar] [CrossRef]
- Conroy, T.; Guo, J.T.; Linington, R.G.; Hunt, N.H.; Payne, R.J. Total synthesis, stereochemical assignment, and antimalarial activity of gallinamide A. Chem. Eur. J. 2011, 17, 13544–13552. [Google Scholar] [CrossRef]
- Conroy, T.; Guo, J.T.; Hunt, N.H.; Payne, R.J. Total synthesis and antimalarial activity of symplostatin 4. Org. Lett. 2010, 12, 5576–5579. [Google Scholar] [CrossRef]
- Linington, R.G.; Clark, B.R.; Trimble, E.E.; Almanza, A.; Urena, L.D.; Kyle, D.E.; Gerwick, W.H. Antimalarial peptides from marine cyanobacteria: Isolation and structural elucidation of gallinamide A. J. Nat. Prod. 2009, 72, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.; Friedman, A.J.; Choi, H.; Hogan, J.; McCammon, J.A.; Hook, V.; Gerwick, W.H. The marine cyanobacterial metabolite gallinamide A is a potent and selective inhibitor of human cathepsin L. J. Nat. Prod. 2014, 77, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Stolze, S.C.; Deu, E.; Kaschani, F.; Li, N.; Florea, B.I.; Richau, K.H.; Colby, T.; van der Hoorn, R.A.; Overkleeft, H.S.; Bogyo, M.; et al. The antimalarial natural product symplostatin 4 is a nanomolar inhibitor of the food vacuole falcipains. Chem. Biol. 2012, 19, 1546–1555. [Google Scholar] [CrossRef] [Green Version]
- Uka, V.; Moore, G.G.; Arroyo-Manzanares, N.; Nebija, D.; De Saeger, S.; Di Mavungu, J.D. Unravelling the diversity of the cyclopiazonic acid family of mycotoxins in Aspergillus flavus by UHPLC triple-TOF HRMS. Toxins 2017, 9, 35. [Google Scholar] [CrossRef]
- Ostry, V.; Toman, J.; Grosse, Y.; Malir, F. Cyclopiazonic acid: 50th anniversary of its discovery. World Mycotoxin J. 2018, 11, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Hymery, N.; Masson, F.; Barbier, G.; Coton, E. Cytotoxicity and immunotoxicity of cyclopiazonic acid on human cells. Toxicol. Vitr. 2014, 28, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Wang, Y.; Wang, L.; Li, G.; Lan, K.; Altmeyer, R.; Zou, G. Cyclopiazonic acid, an inhibitor of calcium-dependent ATPases with antiviral activity against human respiratory syncytial virus. Antivir. Res. 2016, 132, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.-Q.; Du, L.; Fang, Y.-C.; Wang, F.-Z.; Zhu, T.-J.; Gu, Q.-Q.; Zhu, W.-M. Iso-α-Cyclopiazonic acid, a new natural product isolated from the marine-derived fungus Aspergillus flavus CF-3. Chem. Nat. Compd. 2009, 45, 677–680. [Google Scholar] [CrossRef]
- Sun, K.; Li, Y.; Guo, L.; Wang, Y.; Liu, P.; Zhu, W. Indole diterpenoids and isocoumarin from the fungus, Aspergillus flavus, isolated from the prawn, Penaeus vannamei. Mar. Drugs 2014, 12, 3970–3981. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Chen, C.; Wang, J.; Li, X.N.; Wei, G.; Guo, Y.; Yao, G.; Luo, Z.; Zhang, J.; Xue, Y. Penicamedine A, a highly oxygenated hexacyclic indole alkaloid from Penicillium camemberti. Chem. Biodivers. 2015, 12, 1547–1553. [Google Scholar] [CrossRef]
- Kwon, Y.; Kim, S.H.; Shin, Y.; Bae, M.; Kim, B.Y.; Lee, S.K.; Oh, K.B.; Shin, J.; Oh, D.C. A new benzofuran glycoside and indole alkaloids from a sponge-associated rare actinomycete, Amycolatopsis sp. Mar. Drugs 2014, 12, 2326–2340. [Google Scholar] [CrossRef] [Green Version]
- Lan, W.-J.; Wang, K.-T.; Xu, M.-Y.; Zhang, J.-J.; Lam, C.-K.; Zhong, G.-H.; Xu, J.; Yang, D.-P.; Li, H.-J.; Wang, L.-Y. Secondary metabolites with chemical diversity from the marine-derived fungus Pseudallescheria boydii F19-1 and their cytotoxic activity. RSC Adv. 2016, 6, 76206–76213. [Google Scholar] [CrossRef]
- Cao, T.; Ling, J.; Liu, Y.; Chen, X.; Tian, X.; Meng, D.; Pan, H.; Hu, J.; Wang, N. Characterization and abolishment of the cyclopiazonic acids produced by Aspergillus oryzae HMP-F28. Biosci. Biotechnol. Biochem. 2018, 82, 1832–1839. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Mugishima, T.; Komatsu, K.; Sone, T.; Tanaka, M.; Mikami, Y.; Shiro, M.; Hirai, M.; Ohizumi, Y.; Kobayashi, J.I. Speradine A, a new pentacyclic oxindole alkaloid from a marine-derived fungus Aspergillus tamarii. Tetrahedron 2003, 59, 3227–3230. [Google Scholar] [CrossRef]
- Wang, N.; Hu, J.C.; Liu, W.; Wang, S.J. Cyclopiazonic Acid Compound, and Preparation and Application Thereof. Chinese Patent CN103183666A, 3 July 2013. [Google Scholar]
- Zhang, Q.-Q.; Chen, L.; Hu, X.; Xia, Q.-W.; Zhao, Y.-Y.; Zheng, Q.-H.; Liu, Q.-Y. Speradines B-E, Four Novel Tetracyclic Oxindole Alkaloids from the Marine-Derived Fungus Aspergillus oryzae. Heterocycles 2014, 89, 1662–1669. [Google Scholar]
- Hu, X.; Xia, Q.W.; Zhao, Y.Y.; Zheng, Q.H.; Liu, Q.Y.; Chen, L.; Zhang, Q.Q. Speradines F-H, three new oxindole alkaloids from the marine-derived fungus Aspergillus oryzae. Chem. Pharm. Bull. 2014, 62, 942–946. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Peng, J.; Wu, G.; Zhu, T.; Li, G.; Gu, Q.; Li, D. Speradines B-D, oxygenated cyclopiazonic acid alkaloids from the sponge-derived fungus Aspergillus flavus MXH-X104. Tetrahedron 2015, 71, 3522–3527. [Google Scholar] [CrossRef]
- Wang, D.; Bao, Y.-R.; Yang, X.-X.; Meng, X.-S.; Chen, G. A New Alkaloid from Penicillium Dipodomyicola. Chem. Nat. Compd. 2015, 51, 733–735. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Nong, X.; Wei, X.; Qi, S. Oxindole alkaloids from the fungus Penicillium commune DFFSCS026 isolated from deep-sea-derived sediments. Tetrahedron 2015, 71, 610–615. [Google Scholar] [CrossRef]
- Zhou, M.; Miao, M.M.; Du, G.; Li, X.N.; Shang, S.Z.; Zhao, W.; Liu, Z.H.; Yang, G.Y.; Che, C.T.; Hu, Q.F.; et al. Aspergillines A-E, highly oxygenated hexacyclic indole-tetrahydrofuran-tetramic acid derivatives from Aspergillus versicolor. Org. Lett. 2014, 16, 5016–5019. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, L.; Wang, L.; Fang, X.; Zhao, J.; Wang, X.; Li, L.; Liu, H.; Wei, Y.; You, X.; et al. Griseofulvin Derivative and Indole Alkaloids from Penicillium griseofulvum CPCC 400528. J. Nat. Prod. 2017, 80, 371–376. [Google Scholar] [CrossRef]
- Asiri, I.A.M.; Badr, J.M.; Youssef, D.T.A. Penicillivinacine, antimigratory diketopiperazine alkaloid from the marine-derived fungus Penicillium vinaceum. Phytochem. Lett. 2015, 13, 53–58. [Google Scholar] [CrossRef]
- Hiort, J.; Maksimenka, K.; Reichert, M.; Perović-Ottstadt, S.; Lin, W.; Wray, V.; Steube, K.; Schaumann, K.; Weber, H.; Proksch, P. New Natural Products from the Sponge-Derived Fungus Aspergillus niger. J. Nat. Prod. 2004, 67, 1532–1543. [Google Scholar] [CrossRef]
- Schlingmann, G.; Taniguchi, T.; He, H.; Bigelis, R.; Yang, H.Y.; Koehn, F.E.; Carter, G.T.; Berova, N. Reassessing the structure of pyranonigrin. J. Nat. Prod. 2007, 70, 1180–1187. [Google Scholar] [CrossRef]
- Meng, L.-H.; Li, X.-M.; Liu, Y.; Wang, B.-G. Polyoxygenated dihydropyrano [2, 3-c] pyrrole-4, 5-dione derivatives from the marine mangrove-derived endophytic fungus Penicillium brocae MA-231 and their antimicrobial activity. Chin. Chem. Lett. 2015, 26, 610–612. [Google Scholar] [CrossRef]
- Miyake, Y.; Mochizuki, M.; Ito, C.; Itoigawa, M.; Osawa, T. Antioxidative pyranonigrins in rice mold starters and their suppressive effect on the expression of blood adhesion molecules. Biosci. Biotechnol. Biochem. 2008, 72, 1580–1585. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Ito, C.; Tokuda, H.; Osawa, T.; Itoigawa, M. Evaluation of flavoglaucin, its derivatives and pyranonigrins produced by molds used in fermented foods for inhibiting tumor promotion. Biosci. Biotechnol. Biochem. 2010, 74, 1120–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, Y.; Ito, C.; Itoigawa, M.; Osawa, T. Isolation of the antioxidant pyranonigrin-A from rice mold starters used in the manufacturing process of fermented foods. Biosci. Biotechnol. Biochem. 2007, 71, 2515–2521. [Google Scholar] [CrossRef]
- Riko, R.; Nakamura, H.; Shindo, K. Studies on pyranonigrins-isolation of pyranonigrin E and biosynthetic studies on pyranonigrin A. J. Antibiot. 2014, 67, 179–181. [Google Scholar] [CrossRef]
- Dong, J.-J.; Bao, J.; Zhang, X.-Y.; Xu, X.-Y.; Nong, X.-H.; Qi, S.-H. Alkaloids and citrinins from marine-derived fungus Nigrospora oryzae SCSGAF 0111. Tetrahedron Lett. 2014, 55, 2749–2753. [Google Scholar] [CrossRef]
- Hosoe, T.; Fukushima, K.; Takizawa, K. A new pyrrolidine-2, 4-dione derivative, vermelhotin, isolated from unidentified fungus IFM 52672. Heterocycles 2006, 68, 1949–1953. [Google Scholar]
- Kasettrathat, C.; Ngamrojanavanich, N.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Cytotoxic and antiplasmodial substances from marine-derived fungi, Nodulisporium sp. and CRI247-01. Phytochemistry 2008, 69, 2621–2626. [Google Scholar] [CrossRef]
- Leyte-Lugo, M.; Gonzalez-Andrade, M.; Gonzalez Mdel, C.; Glenn, A.E.; Cerda-Garcia-Rojas, C.M.; Mata, R. (+)-Ascosalitoxin and vermelhotin, a calmodulin inhibitor, from an endophytic fungus isolated from Hintonia latiflora. J. Nat. Prod. 2012, 75, 1571–1577. [Google Scholar] [CrossRef]
- Pansanit, A.; Park, E.J.; Kondratyuk, T.P.; Pezzuto, J.M.; Lirdprapamongkol, K.; Kittakoop, P. Vermelhotin, an anti-inflammatory agent, suppresses nitric oxide production in RAW 264.7 cells via p38 inhibition. J. Nat. Prod. 2013, 76, 1824–1827. [Google Scholar] [CrossRef]
- Ganihigama, D.U.; Sureram, S.; Sangher, S.; Hongmanee, P.; Aree, T.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Antimycobacterial activity of natural products and synthetic agents: Pyrrolodiquinolines and vermelhotin as anti-tubercular leads against clinical multidrug resistant isolates of Mycobacterium tuberculosis. Eur. J. Med. Chem. 2015, 89, 1–12. [Google Scholar] [CrossRef]
- Zhu, G.; Kong, F.; Wang, Y.; Fu, P.; Zhu, W. Cladodionen, a Cytotoxic Hybrid Polyketide from the Marine-Derived Cladosporium sp. OUCMDZ-1635. Mar. Drugs 2018, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Sun, X.; Yu, G.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Cladosins A-E, hybrid polyketides from a deep-sea-derived fungus, Cladosporium sphaerospermum. J. Nat. Prod. 2014, 77, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.H.; Wu, G.W.; Zhu, T.J.; Gu, Q.Q.; Li, D.H. Cladosins F and G, two new hybrid polyketides from the deep-sea-derived Cladosporium sphaerospermum 2005-01-E3. J. Asian Nat. Prod. Res. 2015, 17, 120–124. [Google Scholar] [CrossRef]
- Zhang, Z.; He, X.; Wu, G.; Liu, C.; Lu, C.; Gu, Q.; Che, Q.; Zhu, T.; Zhang, G.; Li, D. Aniline-tetramic acids from the deep-sea-derived fungus Cladosporium sphaerospermum L3P3 cultured with the hdac inhibitor saha. J. Nat. Prod. 2018, 81, 1651–1657. [Google Scholar] [CrossRef]
- Chen, S.C.; Liu, Z.M.; Tan, H.B.; Chen, Y.C.; Li, S.N.; Li, H.H.; Guo, H.; Zhu, S.; Liu, H.X.; Zhang, W.M. Tersone A-G, New Pyridone Alkaloids from the Deep-Sea Fungus Phomopsis tersa. Mar. Drugs 2019, 17, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manam, R.R.; Teisan, S.; White, D.J.; Nicholson, B.; Grodberg, J.; Neuteboom, S.T.; Lam, K.S.; Mosca, D.A.; Lloyd, G.K.; Potts, B.C. Lajollamycin, a Nitro-tetraene Spiro-β-lactone-γ-lactam Antibiotic from the Marine Actinomycete Streptomyces nodosus. J. Nat. Prod. 2005, 68, 240–243. [Google Scholar] [CrossRef]
- Ko, K.; Lee, S.H.; Kim, S.H.; Kim, E.H.; Oh, K.B.; Shin, J.; Oh, D.C. Lajollamycins, nitro group-bearing spiro-beta-lactone-gamma-lactams obtained from a marine-derived Streptomyces sp. J. Nat. Prod. 2014, 77, 2099–2104. [Google Scholar] [CrossRef]
- Shin, H.J.; Kim, T.S.; Lee, H.S.; Park, J.Y.; Choi, I.K.; Kwon, H.J. Streptopyrrolidine, an angiogenesis inhibitor from a marine-derived Streptomyces sp. KORDI-3973. Phytochemistry 2008, 69, 2363–2366. [Google Scholar] [CrossRef]
- Fu, P.; La, S.; MacMillan, J.B. 1,3-Oxazin-6-one Derivatives and Bohemamine-Type Pyrrolizidine Alkaloids from a Marine-Derived Streptomyces spinoverrucosus. J. Nat. Prod. 2016, 79, 455–462. [Google Scholar] [CrossRef]
- Kakeya, H.; Takahashi, I.; Okada, G.; Isono, K.; Osada, H. Epolactaene, a novel neuritogenic compound in human neuroblastoma cells, produced by a marine fungus. J. Antibiot. 1995, 48, 733–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushina, Y.; Kobayashi, S.; Kuramochi, K.; Nagata, S.; Sugawara, F.; Sakaguchi, K. Epolactaene, a novel neuritogenic compound in human neuroblastoma cells, selectively inhibits the activities of mammalian DNA polymerases and human DNA topoisomerase II. Biochem. Biophys. Res. Commun. 2000, 273, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, Y.; Kakeya, H.; Shoji, M.; Hayashi, Y.; Dohmae, N.; Osada, H. Epolactaene binds human Hsp60 Cys442 resulting in the inhibition of chaperone activity. Biochem. J. 2005, 387, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Nagumo, Y.; Kakeya, H.; Yamaguchi, J.; Uno, T.; Shoji, M.; Hayashi, Y.; Osada, H. Structure-activity relationships of epolactaene derivatives: Structural requirements for inhibition of Hsp60 chaperone activity. Bioorg. Med. Chem. Lett. 2004, 14, 4425–4429. [Google Scholar] [CrossRef]
- Alvi, K.A.; Casey, A.; Nair, B.G. Pulchellalactam: A CD45 protein tyrosine phosphatase inhibitor from the marine fungus Corollospora pulchella. J. Antibiot. 1998, 51, 515–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, H.; Iwasaki, A.; Sumimoto, S.; Iwatsuki, M.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Omura, S.; Suenaga, K. Isolation and Total Synthesis of Hoshinolactam, an Antitrypanosomal Lactam from a Marine Cyanobacterium. Org. Lett. 2017, 19, 890–893. [Google Scholar] [CrossRef]
- Sangnoi, Y.; Plubrukarn, A.; Arunpairojana, V.; Kanjana-Opas, A. A new antibacterial amino phenyl pyrrolidone derivative from a novel marine gliding bacterium Rapidithrix thailandica. World J. Microbiol. Biotechnol. 2014, 30, 1135–1139. [Google Scholar] [CrossRef]
- Needham, J.; Kelly, M.T.; Ishige, M.; Andersen, R.J. Andrimid and moiramides A-C, metabolites produced in culture by a marine isolate of the bacterium Pseudomonas fluorescens: Structure elucidation and biosynthesis. J. Org. Chem. 1994, 59, 2058–2063. [Google Scholar] [CrossRef]
- Freiberg, C.; Brunner, N.A.; Schiffer, G.; Lampe, T.; Pohlmann, J.; Brands, M.; Raabe, M.; Habich, D.; Ziegelbauer, K. Identification and characterization of the first class of potent bacterial acetyl-CoA carboxylase inhibitors with antibacterial activity. J. Biol. Chem. 2004, 279, 26066–26073. [Google Scholar] [CrossRef] [Green Version]
- Pohlmann, J.; Lampe, T.; Shimada, M.; Nell, P.G.; Pernerstorfer, J.; Svenstrup, N.; Brunner, N.A.; Schiffer, G.; Freiberg, C. Pyrrolidinedione derivatives as antibacterial agents with a novel mode of action. Bioorg. Med. Chem. Lett. 2005, 15, 1189–1192. [Google Scholar] [CrossRef]




















© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Chen, S.; Li, J.; Liu, L. The Biological and Chemical Diversity of Tetramic Acid Compounds from Marine-Derived Microorganisms. Mar. Drugs 2020, 18, 114. https://doi.org/10.3390/md18020114
Jiang M, Chen S, Li J, Liu L. The Biological and Chemical Diversity of Tetramic Acid Compounds from Marine-Derived Microorganisms. Marine Drugs. 2020; 18(2):114. https://doi.org/10.3390/md18020114
Chicago/Turabian StyleJiang, Minghua, Senhua Chen, Jing Li, and Lan Liu. 2020. "The Biological and Chemical Diversity of Tetramic Acid Compounds from Marine-Derived Microorganisms" Marine Drugs 18, no. 2: 114. https://doi.org/10.3390/md18020114