Interaction of Thalassia testudinum Metabolites with Cytochrome P450 Enzymes and Its Effects on Benzo(a)pyrene-Induced Mutagenicity

 , and
, and
Abstract
:1. Introduction
2. Results
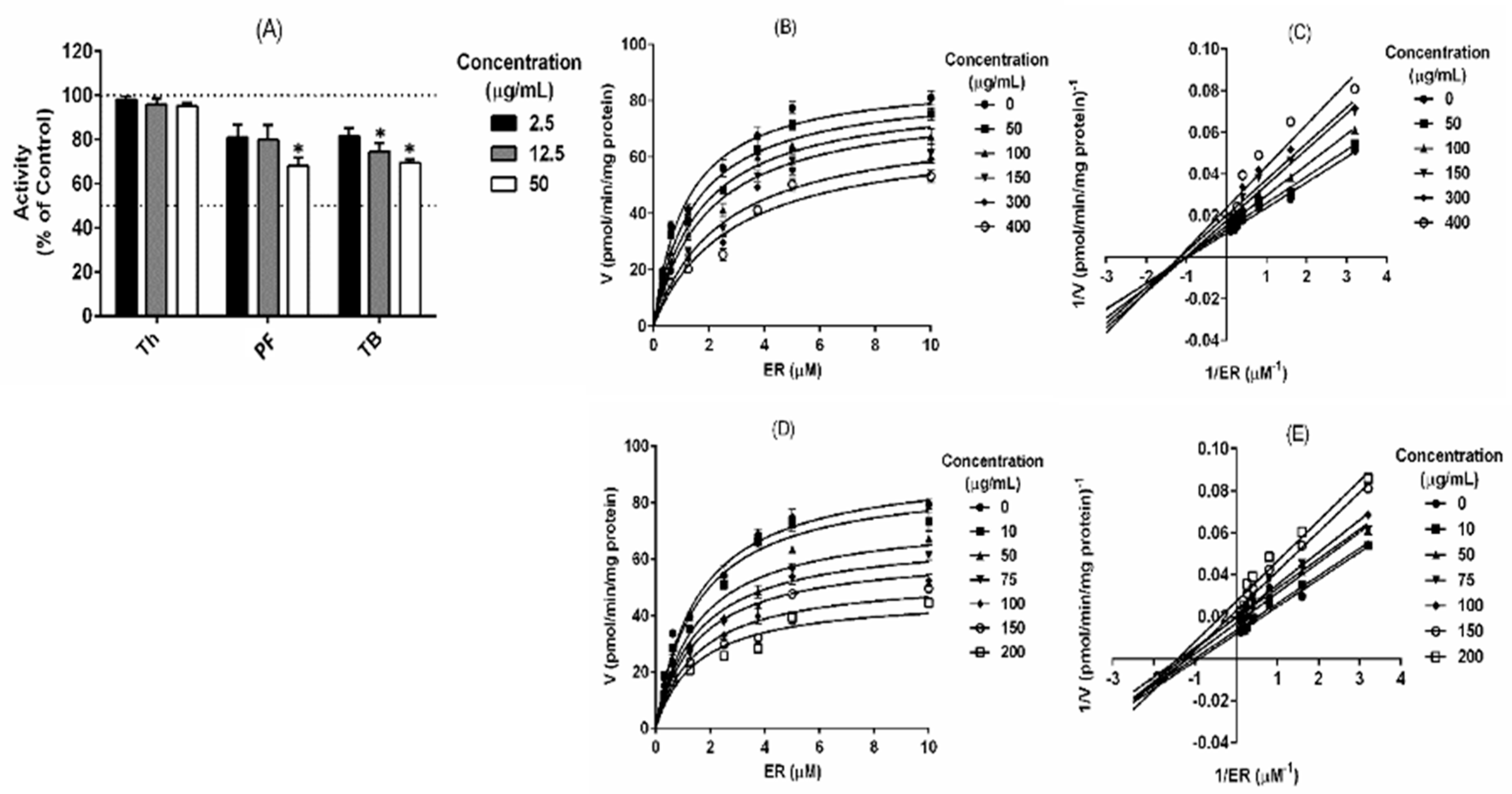
2.1. Tested Compounds Modulate Rat CYP1A But Not CYPB2 Activity
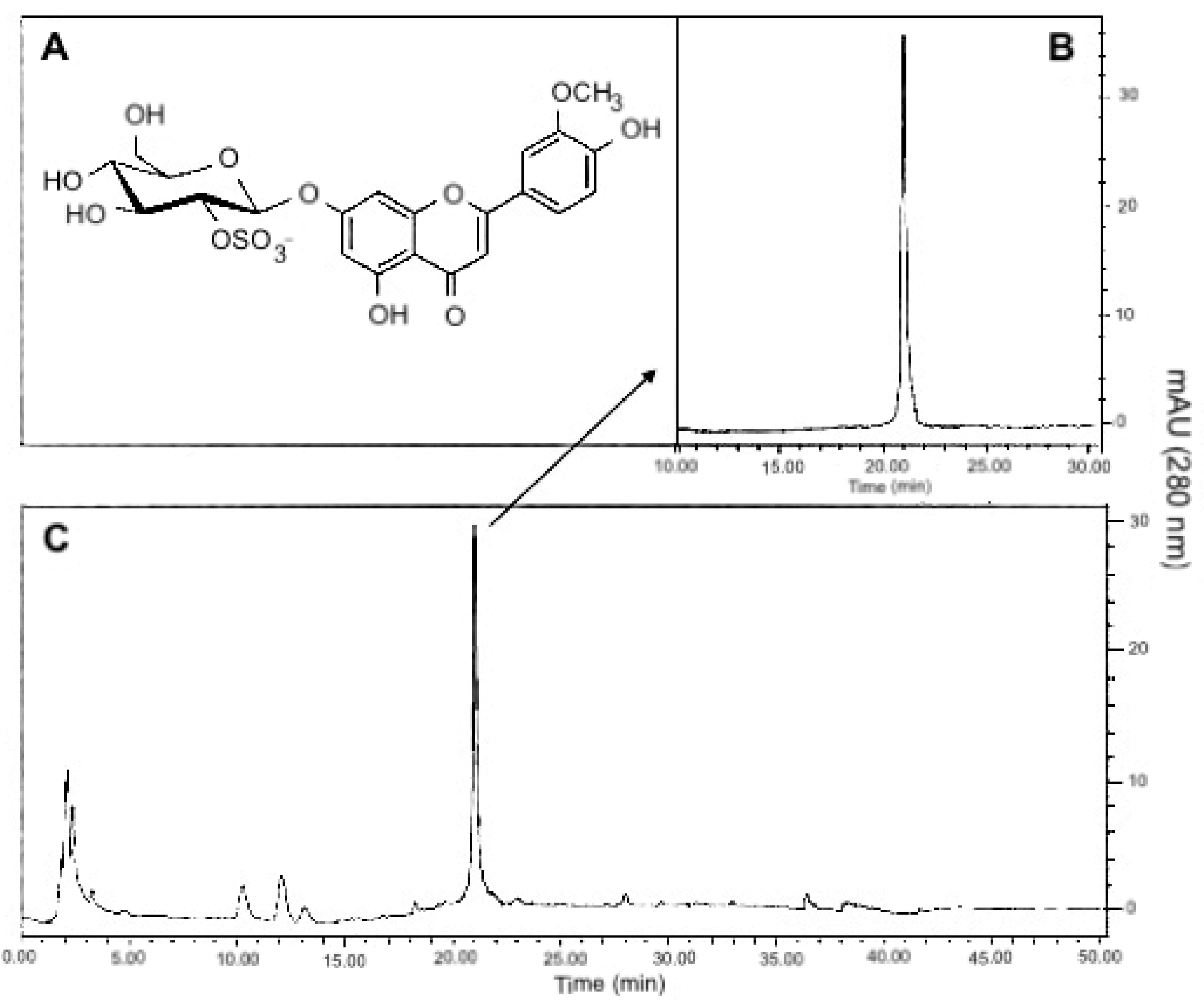
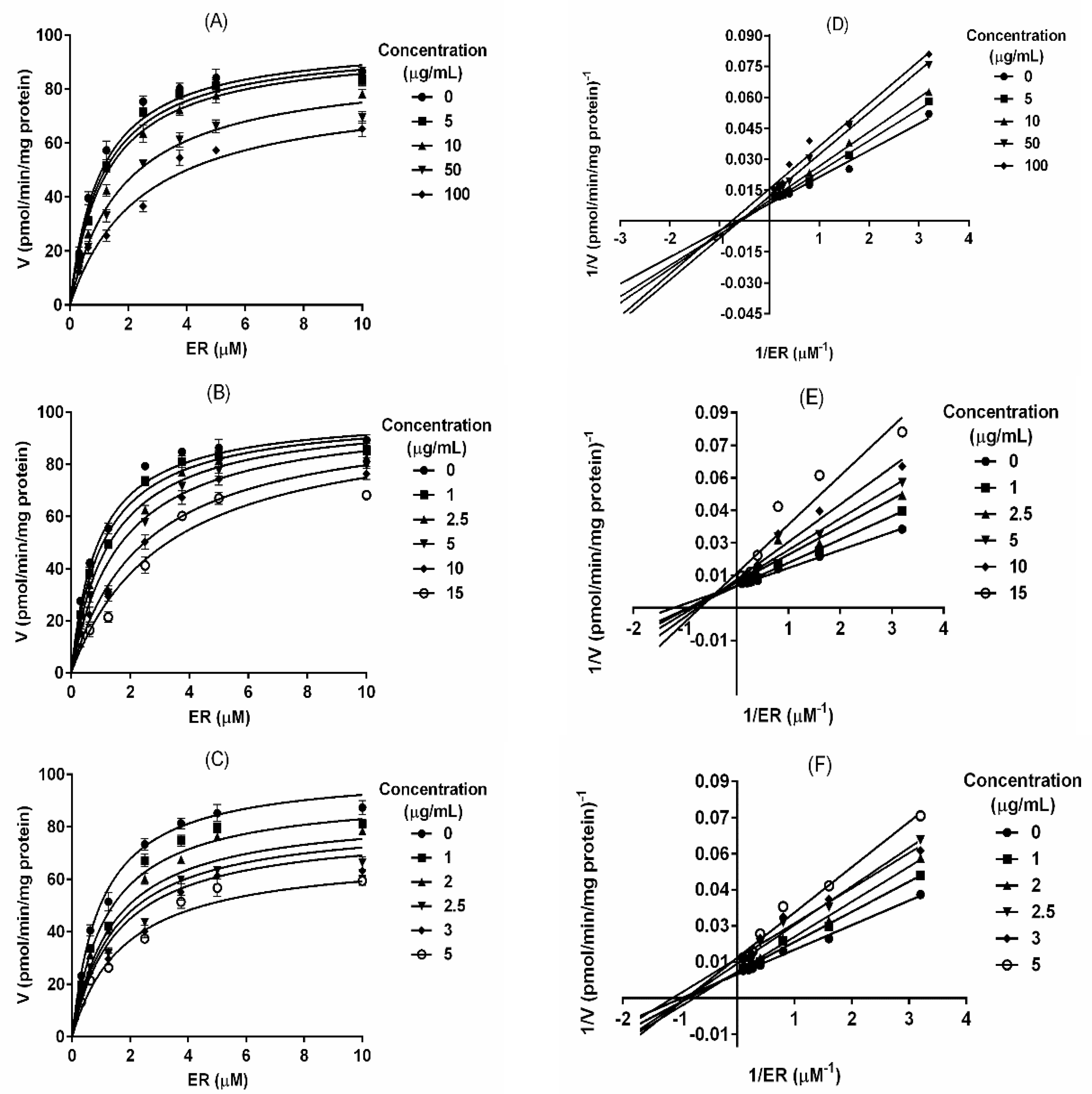
2.2. T. testudinum Extract, Polyphenolic Fraction and Thalassiolin B Are CYP1A1 Mixed-Type Inhibitors
2.3. Polyphenolic Fraction and Thalassiolin B Modulate the Human CYP1A1 Activity
2.4. Antimutagenic Effect of T. testudinum Extract, Polyphenolic Fraction and Thalassiolin B against Benzo[a]pyrene-Induced Mutagenicity in S. typhimurium
2.5. T. testudinum Extract Reduces Oxidative Damage and Micronuclei Formation in BP-Exposed Mice
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Vegetal Material
4.3. Rat Liver S9 and Microsomal Fraction Obtaining
4.4. Bacterial Membrane Fraction Obtaining
4.5. Enzymatic Activity Assays
4.5.1. CYP1A1 and CYP1A2 Activities
4.5.2. CYP2B1 and CYP2B2 Activities
4.6. Kinetic Analysis of Enzyme Inhibition
4.7. Ames Test
4.8. Effects of the T. testudinum Extract against BP-Induced DNA Damage in Mice
4.8.1. Oxidative Stress Biomarkers Determination
4.8.2. Micronuclei Formation Determination
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. 7 Million Premature Deaths Annually Linked to Air Pollution. Available online: https://www.who.int/mediacentre/news/releases/2014/air-pollution/en/#:~:text=25%20March%202014%20%7C%20Geneva%20%2D%20In,result%20of%20air%20pollution%20exposure (accessed on 24 March 2020).
- Kim, J.H.; Stansbury, K.H.; Trush, M.A.; Strickland, P.T.; Sutter, T.R. Metabolism of benzo[a]pyrene and benzo[a]pyrene-7,8-diol by human cytochrome P450 1B1. Carcinogenesis 1998, 19, 1847–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, D.; Kisselev, P.; Cascorbi, I.; Schunck, W.H.; Roots, I. Differential metabolism of benzo[a]pyrene and benzo[a]pyrene-7,8-dihydrodiol by human CYP1A1 variants. Carcinogenesis 2001, 22, 453–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, M.R.; Brookes, P.; Beland, F.A.; Harvey, R.G. The reaction of (±)-7α, 8β-dihydroxy-9β, 10β-epoxy-7,8,9,10-tetrahydrobenzo(a)pyrene with dna. Int. J. Cancer 1976, 18, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Boysen, G.; Hecht, S.S. Analysis of DNA and protein adducts of benzo[a]pyrene in human tissues using structure-specific methods. Mutat. Res. 2003, 543, 17–30. [Google Scholar] [CrossRef]
- Rojas, M.; Marie, B.; Vignaud, J.M.; Martinet, N.; Siat, J.; Grosdidier, G.; Cascorbi, I.; Alexandrov, K. High DNA damage by benzo[a]pyrene 7,8-diol-9,10-epoxide in bronchial epithelial cells from patients with lung cancer: Comparison with lung parenchyma. Cancer Lett. 2004, 207, 157–163. [Google Scholar] [CrossRef]
- Alexandrov, K.; Rojas, M.; Rolando, C. DNA damage by benzo(a)pyrene in human cells is increased by cigarette smoke and decreased by a filter containing rosemary extract, which lowers free radicals. Cancer Res. 2006, 66, 11938–11945. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, G.; Yoon, J.H.; Liu, L.; Tommasi, S.; Wilczynski, S.P.; Dammann, R. Methylation of the RASSF1A gene in human cancers. Biol. Chem. 2002, 383, 907–914. [Google Scholar] [CrossRef]
- Sporn, M.B.; Suh, N. Chemoprevention of cancer. Carcinogenesis 2000, 21, 525–530. [Google Scholar] [CrossRef]
- Stoner, G.D.; Morse, M.A.; Kelloff, G.J. Perspectives in cancer chemoprevention. Environ. Health Perspect. 1997, 105, 945–954. [Google Scholar]
- Guengerich, F.P.; Shimada, T. Activation of procarcinogens by human cytochrome P450 enzymes. Mutat. Res. 1998, 400, 201–213. [Google Scholar] [CrossRef]
- Yang, C.S.; Smith, T.J.; Hong, J.Y. Cytochrome P-450 Enzymes as Targets for Chemoprevention against Chemical Carcinogenesis and Toxicity: Opportunities and Limitations. Cancer Res. 1994, 54, 1982s–1986s. [Google Scholar] [PubMed]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessien, M.; El-Gendy, S.; Donia, T.; Sikkena, M.A. Growth inhibition of human non-small lung cancer cells h460 by green tea and ginger polyphenols. Anticancer Agents Med. Chem. 2012, 12, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wetta, H. Genetics and epigenetics in tumorigenesis: Acting separately or linked. Austin J. Clin. Med. 2014, 1, 1016. [Google Scholar]
- Barron, C.C.; Moore, J.; Tsakiridis, T.; Pickering, G.; Tsiani, E. Inhibition of human lung cancer cell proliferation and survival by wine. Cancer Cell. Int. 2014, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Bauer, D.; Pimentel de Abreu, J.; Silva, H.S.; Goes-Neto, A.; Bello, M.G.; Teodoro, A.J. Antioxidant Activity and Cytotoxicity Effect of Cocoa Beans Subjected to Different Processing Conditions in Human Lung Carcinoma Cells. Oxid. Med. Cell. Longev. 2016, 2016, 7428515. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Ekambaram, G.; Sakthisekaran, D. Cytoprotective Effect of Mangiferin on Benzo(a)pyrene-Induced Lung Carcinogenesis in Swiss Albino Mice. Basic Clin. Pharmacol. Toxicol. 2008, 103, 137–142. [Google Scholar] [CrossRef]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, X.; Han, L.; Li, X.; Xue, Z.; Zhou, F. Antioxidant and antitumor effects and immunomodulatory activities of crude and purified polyphenol extract from blueberries. Front. Chem. Sci. Eng. 2016, 10, 108–119. [Google Scholar] [CrossRef]
- Amararathna, M.; Johnston, M.R.; Vasantha, H.P. Plant Polyphenols as Chemopreventive Agents for Lung Cancer. Int. J. Mol. Sci. 2016, 17, 1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, T.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine natural products as anticancer drugs. Mol. Cancer Ther. 2005, 4, 333–342. [Google Scholar] [PubMed]
- Folmer, F.; Jaspars, M.; Dicato, M.; Diederich, M. Photosynthetic marine organisms as a source of anticancer compounds. Phytochem. Rev. 2010, 9, 557–579. [Google Scholar] [CrossRef]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied. Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef]
- Rowley, D.C.; Hansen, M.S.; Rhodes, D.; Sotriffer, C.A.; Ni, H.; McCammon, J.A.; Bushman, F.D.; Fenical, W. Thalassiolins A-C: New marine-derived inhibitors of HIV cDNA integrase. Bioorg. Med. Chem. 2003, 10, 3619–3625. [Google Scholar] [CrossRef]
- Kontiza, I.; Stavri, M.; Zloh, M.; Vagias, C.; Gibbons, S.; Roussis, V. New metabolites with antibacterial activity from the marine angiosperm Cymodocea nodosa. Tetrahedron 2008, 64, 1696–1702. [Google Scholar] [CrossRef]
- Achamlale, S.; Rezzonico, B.; Grignon-Dubois, M. Evaluation of Zostera detritus as a potential new source of zosteric acid. J. Appl. Phycol. 2009, 21, 347–352. [Google Scholar] [CrossRef]
- Carbone, M.; Gavagnin, M.; Mollo, E.; Bidello, M.; Roussis, V.; Cimino, G. Further syphonosides from the sea hare Syphonota geographica and the sea-grass Halophila stipulacea. Tetrahedron 2008, 64, 191–196. [Google Scholar] [CrossRef]
- Hamdy, A.H.; Mettwally, W.S.; Abou El Fotouh, M.; Rodriguez, B.; El-Dewany, A.I.; El-Toumy, S.A.; Hussein, A.A. Bioactive Phenolic Compounds from the Egyptian Red Sea Seagrass Thalassodendron ciliatum. Z. Für Nat. C 2012, 67, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Regalado, E.L.; Rodríguez, M.; Menéndez, R.; Concepción, Á.A.; Nogueiras, C.; Laguna, A.; Rodríguez, A.A.; Williams, D.E.; Lorenzo-Luaces, P.; Valdés, O.; et al. Repair of UVB-damaged skin by the antioxidant sulphated flavone glycoside thalassiolin B isolated from the marine plant Thalassia testudinum Banks ex Konig. Mar Biotechnol. 2009, 11, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Regalado, E.L.; Menendez, R.; Valdés, O.; Morales, R.A.; Laguna, A.; Thomas, O.P.; Hernandez, Y.; Nogueiras, C.; Kijjoa, A. Phytochemical Analysis and Antioxidant Capacity of BM-21, a Bioactive Extract Rich in Polyphenolic Metabolites from the Sea Grass Thalassia testudinum. Nat. Prod. Commun. 2012, 7, 47–50. [Google Scholar] [CrossRef]
- Rodeiro, I.; Donato, M.; Martinez, I.; Hernandez, I.; Garrido, G.; González-Lavaut, J.; Menendez, R.; Laguna, A.; Castell, J.V.; Gómez-Lechón, M.J. Potential hepatoprotective effects of new Cuban natural products in rat hepatocytes culture. Toxicol. In Vitro 2008, 22, 1242–1249. [Google Scholar] [CrossRef]
- Garateix, A.; Salceda, E.; Menéndez, R.; Regalado, E.L.; López, O.; García, T.; Morales, R.A.; Laguna, A.; Thomas, O.P.; Soto, E. Antinociception produced by Thalassia testudinum extract BM-21 is mediated by the inhibition of acid sensing ionic channels by the phenolic compound thalassiolin B. Mol. Pain 2011, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodeiro, I.; Hernández, I.; Herrera, J.A.; Riera, M.; Donato, M.T.; Tolosa, L.; González, K.; Ansoar, Y.; Gómez-Lechón, M.J.; Vanden Berghe, W.; et al. Assessment of the cytotoxic potential of an aqueous-ethanolic extract from Thalassia testudinum angiosperm marine grown in the Caribbean Sea. J. Pharm. Pharmacol. 2018, 70, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Rodeiro, I.; Gomez-Lechon, M.J.; Tolosa, L.; Pérez, G.; Hernández, I.; Menéndez, R.; Regalado, E.L.; Castell, J.V.; Donato, M.T. Modulation of biotransformation and elimination systems by BM-21, an aqueous ethanolic extract from Thalassia testudinum, and thalassiolin B on human hepatocytes. J. Funct. Foods 2012, 4, 167–176. [Google Scholar] [CrossRef]
- Miguel, V.; Otero, J.A.; Barrera, B.; Rodeiro, I.; Prieto, J.G.; Merino, G.; Álvarez, A.I. ABCG2/BCRP interaction with the sea grass Thalassia testudinum. Drug Metab. Pers. Ther. 2015, 30, 251–256. [Google Scholar] [CrossRef]
- Rodeiro-Guerra, I.; Hernández-Ojeda, S.L.; Herrera-Isidrón, J.A.; Hernández-Balmaseda, I.; Padrón-Yaquis, S.; del Rosario Olguín-Reyes, S.; Alejo-Rodríguez, P.L.; Ronquillo-Sánchez, M.D.; Camacho-Carranza, R.; del Valle, R.M.; et al. Study of the interaction of an extract obtained from the marine plant Thalassia testudinum with pahse I metabolism in rats. Rev. Int. Contam. Ambient. 2017, 33, 547–557. [Google Scholar] [CrossRef]
- Clementino, M.; Shi, X.; Zhang, Z. Prevention of Polyphenols against Carcinogenesis Induced by Environmental Carcinogens. J. Environ. Pathol. Toxicol. Oncol. 2017, 36, 87–98. [Google Scholar] [CrossRef]
- Omidian, K.; Rafiei, H.; Bandy, B. Polyphenol inhibition of benzo[a]pyrene-induced oxidative stress and neoplastic transformation in an in vitro model of carcinogenesis. Food Chem. Toxicol. 2017, 106, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T. Inhibition of Carcinogen-Activating Cytochrome P450 Enzymes by Xenobiotic Chemicals in Relation to Antimutagenicity and Anticarcinogenicity. Toxicol. Res. 2017, 33, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.H.; Marcus, C.E.; Ho, D.K.; Cassady, J.M.; Baird, W.M. Effects of synthetic and naturally occurring flavonoids on benzo[a]pyrene metabolism by hepatic microsomes prepared from rats treated with cytochrome P-450 inducers. Cancer Lett. 1991, 60, 15–24. [Google Scholar] [CrossRef]
- De la Torre, E.; Rodeiro, I.; Menéndez, R.; Pérez, C.D. Thalassia testudinum, a sea plant with great therapeutical potentialities. Rev. Cubana Plant. Med. 2012, 17, 288–296. [Google Scholar]
- Menéndez, R.; García, T.; Garateix, A.; Morales, R.A.; Regalado, E.L.; Laguna, A.; Valdés, O.; Fernández, M.D. Neuroprotective and antioxidant effects of Thalassia testudinum extract BM-21, against acrylamide-induced neurotoxicity in mice. J. Pharm. Pharmacogn. Res. 2014, 2, 53–62. [Google Scholar]
- Arora, S. In vivo prediction of CYP-mediated metabolic interaction potential of formononetin and biochanin a using in vitro human and rat CYP450 inhibition data. Toxicol. Lett. 2015, 239, 1–8. [Google Scholar] [CrossRef]
- Delgado-Roche, L.; Santes-Palacios, R.; Herrera, J.A.; Hernández, S.L.; Riera, M.; Fernández, M.D.; Mesta, F.; Garrido, G.; Rodeiro, I.; Espinosa-Aguirre, J.J. Regulation of Human Cytochrome P4501A1 (hCYP1A1): A Plausible Target for Chemoprevention? Biomed. Res. Int. 2016, 2016, 5341081. [Google Scholar]
- Santes-Palacios, R.; Marroquín-Pérez, A.; Hernández-Ojeda, S.L.; Camacho-Carranza, R.; Govozensky, T.; Espinosa-Aguirre, J.J. Human CYP1A1 inhibition by flavonoids. Toxicol. In Vitro 2020, 62, 104681. [Google Scholar] [CrossRef]
- Zhai, S.; Dai, R.; Friedman, F.K.; Vestal, R.E. Comparative inhibition of human cytochromes P450 1A1 and 1A2 by flavonoids. Drug Metab. Dispos. 1998, 26, 989–992. [Google Scholar]
- Pastrakuljic, A.; Tang, B.K.; Roberts, E.A.; Kalow, W. Distinction of CYP1A1 and CYP1A2 activity by selective inhibition using fluvoxamine and isosafrole. Biochem. Pharmacol. 1997, 53, 531–538. [Google Scholar] [CrossRef]
- Schwarz, D.; Roots, I. In vitro assessment of inhibition by natural polyphenols of metabolic activation of procarcinogens by human CYP1A1. Biochem. Biophys. Res. Commun. 2003, 303, 902–907. [Google Scholar] [CrossRef]
- Sundberg, K.; Dreij, K.; Seidel, A.; Jernström, B. Glutathione conjugation and DNA adduct formation of dibenzo[a,l]pyrene and benzo[a]pyrene diol epoxides in V79 cells stably expressing different human glutathione transferases. Chem. Res. Toxicol. 2002, 15, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, K.; Cascorbi, I.; Rojas, M.; Bouvier, G.; Kriek, E.; Bartsch, H. CYP1A1 and GSTM1 genotypes affect benzo[a]pyrene DNA adducts in smokers’ lung: Comparison with aromatic/hydrophobic adduct formation. Carcinogenesis 2002, 23, 1969–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, W.; Mahadevan, B. Carcinogenic polycyclic aromatic hydrocarbon-DNA adducts and mechanism of action. Environ. Mol. Mutagen. 2005, 45, 106–114. [Google Scholar] [CrossRef]
- Hodek, P.; Koblihová, J.; Kizek, R.; Frei, E.; Arlt, V.M.; Stivorobá, M. The relationship between DNA adduct formation by benzo[a]pyrene and expression of its activation enzyme cytochrome P450 1A1 in rat. Environ. Toxicol. Pharmacol. 2013, 36, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Fernández, M.D.; Llanio, M.; Arteaga, F.; Dajas, F.; Echeverri, C.; Ferreira, M.; Hernández, I.; Cabrera, B.; Rodríguez, M.; Aneiros, A. Propiedades anti-inflamatoria-analgésica y antioxidante de una planta marina. Avicennia 2003, 16, 31–35. [Google Scholar]
- Kamaraj, S.; Vinodhkumar, R.; Anandakumar, P.; Jagan, S.; Ramakrishnan, G.; Devaki, T. The effects of quercetin on antioxidant status and tumor markers in the lung and serum of mice treated with benzo(a)pyrene. Biol. Pharm. Bull. 2007, 30, 2268–2273. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.P.; Batish, D.; Kohli, R.; Arora, K. Arsenic-induced root growth inhibition in mung bean (Phaseolus aureus Roxb.) is due to oxidative stress resulting from enhanced lipid peroxidation. Plant. Growth Regul. 2007, 53, 65–73. [Google Scholar] [CrossRef]
- Nebert, D.W.; Shi, Z.; Gálvez-Peralta, M.; Uno, S.; Dragin, N. Oral benzo[a]pyrene: Understanding pharmacokinetics, detoxication, and consequences—Cyp1 knockout mouse lines as a paradigm. Mol. Pharmacol. 2013, 84, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Gillam, E.M.; Ohmori, S.; Tukey, R.H.; Guengerich, F.P. Expression of modified human cytochrome P450 1A1 in Escherichia coli: Effects of 5′ substitution, stabilization, purification, spectral characterization, and catalytic properties. Arch. Biochem. Biophys. 1994, 312, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P.; Martin, M.V. Purification of cytochromes P450: Products of bacterial recombinant expression systems. Methods Mol. Biol. 2006, 320, 31–37. [Google Scholar] [PubMed]
- Burke, M.D.; Thompson, S.; Weaver, R.J.; Wolf, C.R.; Mayer, R.T. Cytochrome P450 specificities of alkoxyresorufin O-dealkylation in human and rat liver. Biochem. Pharmacol. 1994, 48, 923–936. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Hayashi, M.; Tice, R.R.; MacGregor, J.T.; Anderson, D.; Blakey, D.H.; Kirsh-Volders, M.; Oleson, F.B.; Pacchierotti, F.; Romagna, F.; Shimada, H.; et al. In vivo rodent erythrocyte micronucleus assay. Mutat. Res. 1994, 312, 293–304. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CYP | Product | Test Product Concentrations (µg/mL) | ||||
|---|---|---|---|---|---|---|
| rCYP1A1 | 2.5 | 5.0 | 12.5 | 25.0 | 50.0 | |
| Th | 74.88 ±2.92 a | 63.11 ±8.22 a,* | 49.68 ±8.04 a,** | ND | ND | |
| PF | 46.61 ±3.10 b,** | 43.31 ±0.94 b,** | 31.76 ±6.64 b,** | 27.07 ±1.95 a,** | 24.00 ±2.97 a,** | |
| TB | 53.83 ±1.08 c,* | 42.97 ±4.66 b,** | 42.35 ±0.84 a,** | 29.99 ±3.03 a,** | 35.15 ±1.92 b,** | |
| rCYP1A2 | 2.5 | 5.0 | 12.5 | 25.0 | 50.0 | |
| Th | 100.47 ±10.66 a | 99.88 ±5.25 a | 94.65 ±14.80 a | ND | ND | |
| PF | 83.93 ±2.55 a | 71.87 ±2.85 b | 62.05 ±4.92 b,* | 55.02 ±2.84 a,* | 56.02 ±2.04 a,* | |
| TB | 81.01 ±8.06 a | 75.04 ±2.78 b | 57.08 ±4.70 b,* | 63.36 ±4.90 a,* | 57.03 ±4.07 a,* | |
| Inhibitor | Parameter | Rat CYP1A1 | Human CYP1A1 |
|---|---|---|---|
| ER | Vmax (pmol/min/mgPr) | 2396.00 ± 116.20 | 95.20 ± 8.14 |
| Km (μM) | 0.42 ± 0.05 | 0.34 ± 0.02 | |
| Th | Type of inhibition | Mixed | - |
| Ki (μg/mL) | 54.16 ± 9.09 | - | |
| α | 8.66 ± 2.82 | - | |
| PF | Type of inhibition | Mixed | Mixed |
| Ki (μg/mL) | 5.96 ± 1.55 | 197.10 ± 63.40 | |
| α | 370.60 ± 56.86 | 7.14 ± 5.67 | |
| TB | Type of inhibition | Mixed | Non-competitive |
| Ki (μg/mL) | 3.05 ± 0.89 | 203.10 ± 17.29 | |
| α | 3.65 ± 0.86 | - |
| Treatments | His+ Revertants/Plate (% Inhibition) |
|---|---|
| S9-Control | 23.1 ± 2.0 |
| S9-Control vehicle + BP | 746.2 ± 32.3 |
| S9-Th (10 µg/mL) + BP | 714.4 ± 8.5 |
| S9-Th (100 µg/mL) + BP | 678.0 ± 11.2 ** (10%) |
| S9-Th (1000 µg/mL) + BP | 547.4 ± 5.9 *** (27%) |
| S9-PF (5 µg/mL) + BP | 713.4 ± 8.8 |
| S9-PF (50 µg/mL) + BP | 665.5 ± 10.1 ** (11%) |
| S9-PF (500 µg/mL) + BP | 497.3 ± 10.7 *** (34%) |
| S9-TB (400 µg/mL) + BP | 512.1 ± 28.4 *** (32%) |
| Treatment (mg/kg) | MDA (µM/mgPr) | AOPP (µM chloramines/mgPr) | GSH (µM/mgPr) |
|---|---|---|---|
| Control (vehicle) | 3.17 ± 0.5 | 7.41 ± 1.3 | 495.1 ± 67.8 |
| BP | 7.31 ± 0.2 a | 14.67 ± 1.2 a | 149.7 ± 63.5 a |
| Th + BP | 5.55 ± 0.6 b | 11.64 ± 1.5 b | 352.8 ± 35.1b |
| Treatment (mg/kg) | PCE/NCE | MN/PCE |
|---|---|---|
| Control (vehicle) | 1.8 ± 0.26 b | 4.0 ± 0.7 b |
| BP | 3.4 ± 0.78 a | 17.0 ± 1.7 a |
| Th + BP | 2.0 ± 0.21 b | 7.0 ± 1.0 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Roche, L.; Santes-Palacios, R.; Herrera, J.A.; Hernández, S.L.; Riera, M.; Fernández, M.D.; Mesta, F.; Garrido, G.; Rodeiro, I.; Espinosa-Aguirre, J.J. Interaction of Thalassia testudinum Metabolites with Cytochrome P450 Enzymes and Its Effects on Benzo(a)pyrene-Induced Mutagenicity. Mar. Drugs 2020, 18, 566. https://doi.org/10.3390/md18110566
Delgado-Roche L, Santes-Palacios R, Herrera JA, Hernández SL, Riera M, Fernández MD, Mesta F, Garrido G, Rodeiro I, Espinosa-Aguirre JJ. Interaction of Thalassia testudinum Metabolites with Cytochrome P450 Enzymes and Its Effects on Benzo(a)pyrene-Induced Mutagenicity. Marine Drugs. 2020; 18(11):566. https://doi.org/10.3390/md18110566
Chicago/Turabian StyleDelgado-Roche, Livan, Rebeca Santes-Palacios, José A. Herrera, Sandra L. Hernández, Mario Riera, Miguel D. Fernández, Fernando Mesta, Gabino Garrido, Idania Rodeiro, and Jesús Javier Espinosa-Aguirre. 2020. "Interaction of Thalassia testudinum Metabolites with Cytochrome P450 Enzymes and Its Effects on Benzo(a)pyrene-Induced Mutagenicity" Marine Drugs 18, no. 11: 566. https://doi.org/10.3390/md18110566